
Enhancement of Phenol Biodegradation by Pseudochrobactrum sp. through Ultraviolet-Induced Mutation

Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Identification of the Strain

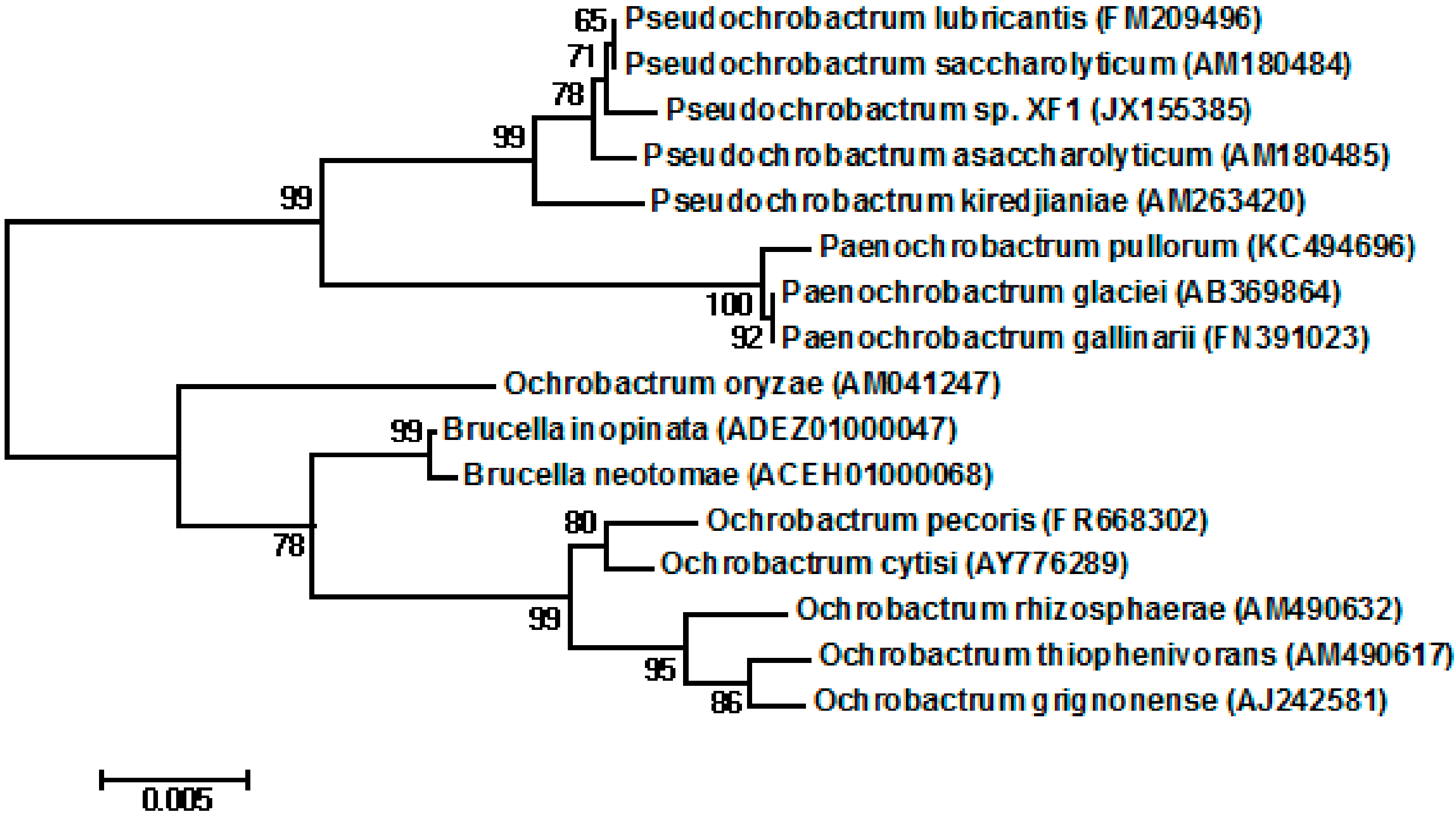{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Strain XF1 | Strain KSS 7.8T |
|---|---|---|
| Morphology | rod-shaped | rod-shaped |
| Pigmentation | white | white |
| Motility | − | − |
| Gram reaction | − | − |
| Catalase | + | not detected |
| Nitrate reduction | + | + |
| Sugars assimilation: | ||
| Glucose | + | + |
| Arabinose | − | − |
| Mannose | + | + |
| Fructose | + | + |
| N-Acetyl-glucosamine | (+) | + |
| Galactose | + | + |

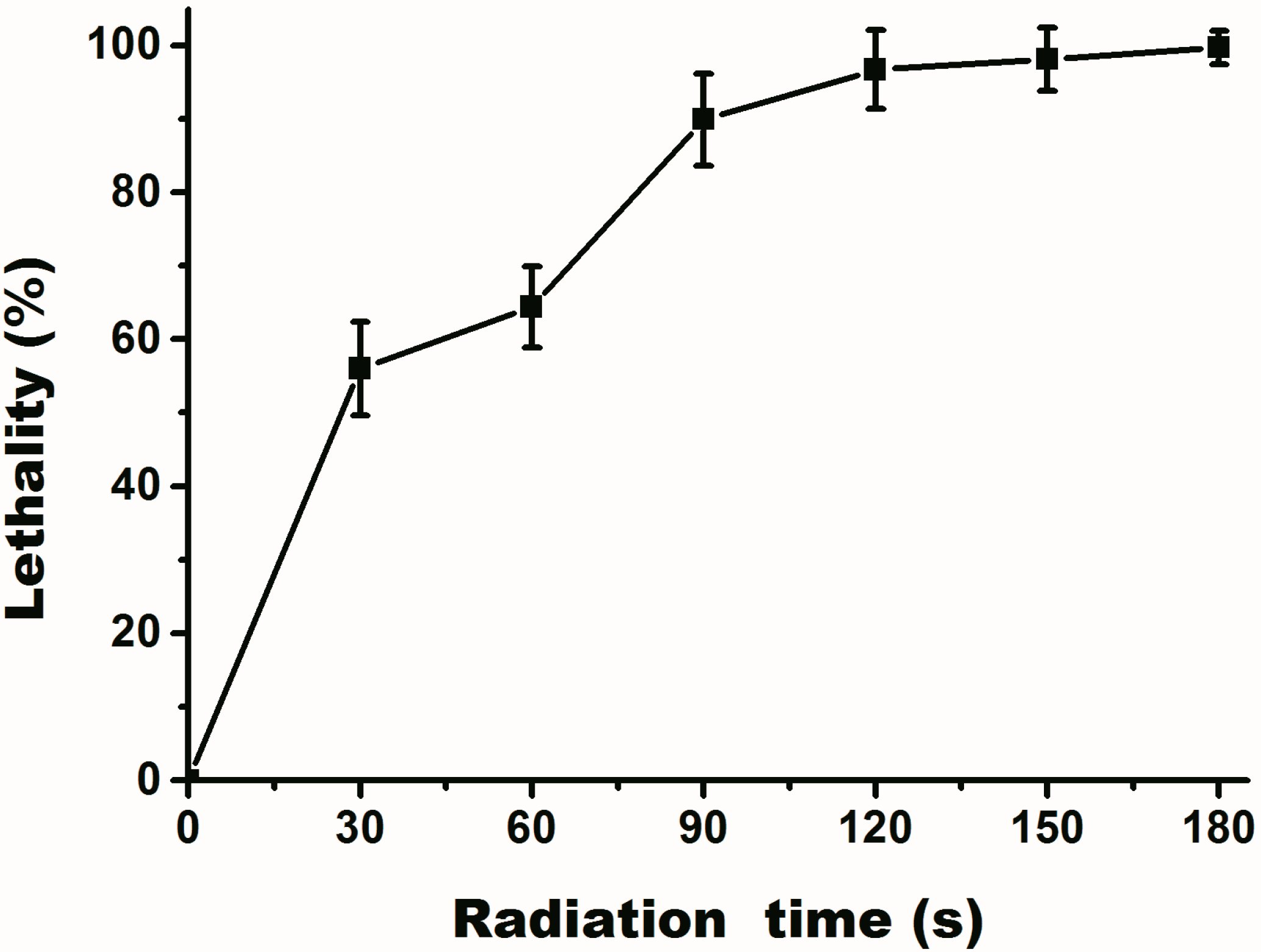
2.2. Assay of UV Mutation

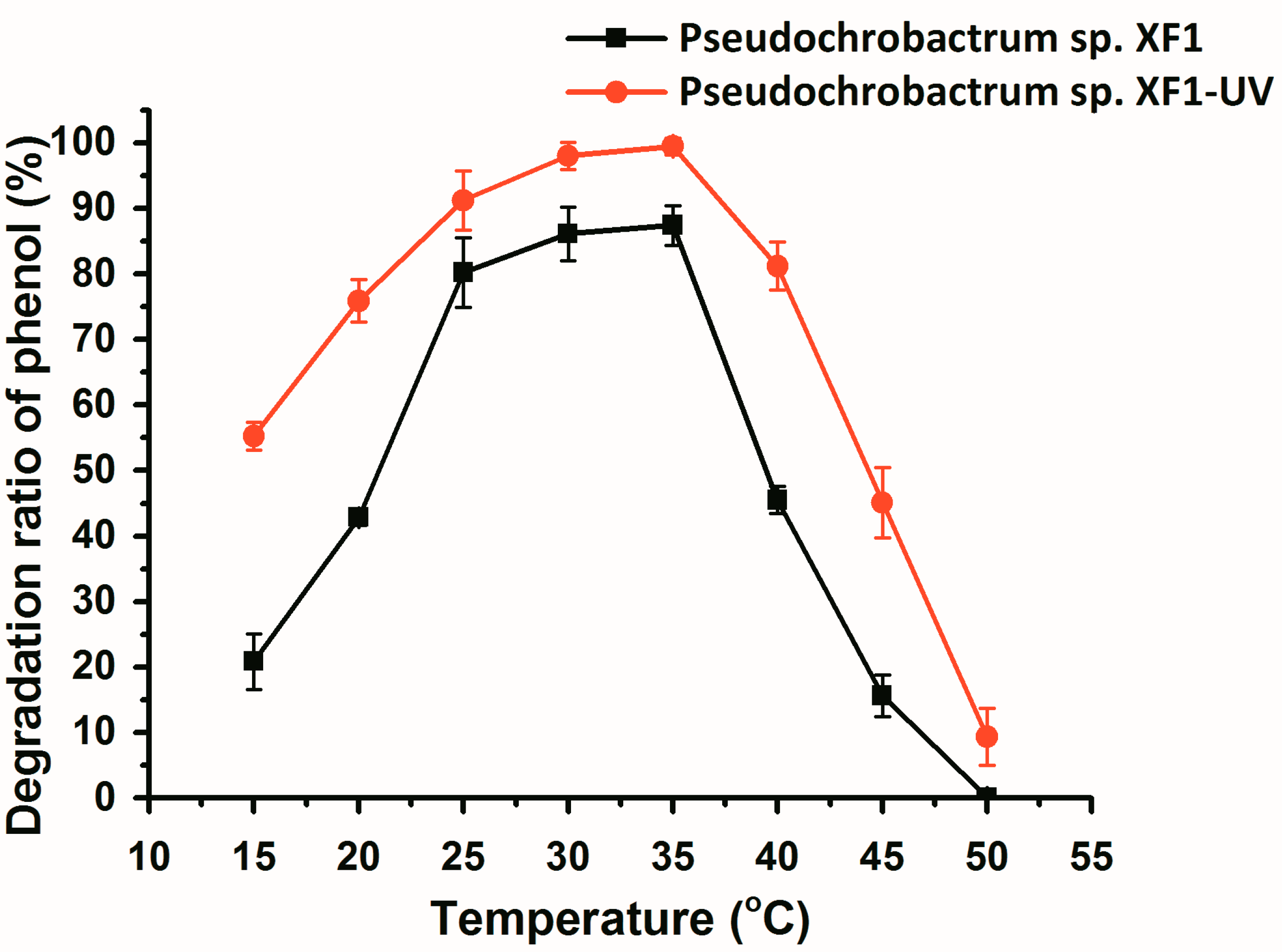
2.3. Effect of pH and Temperature on Degradation of Phenol by Wild and Mutant Bacteria


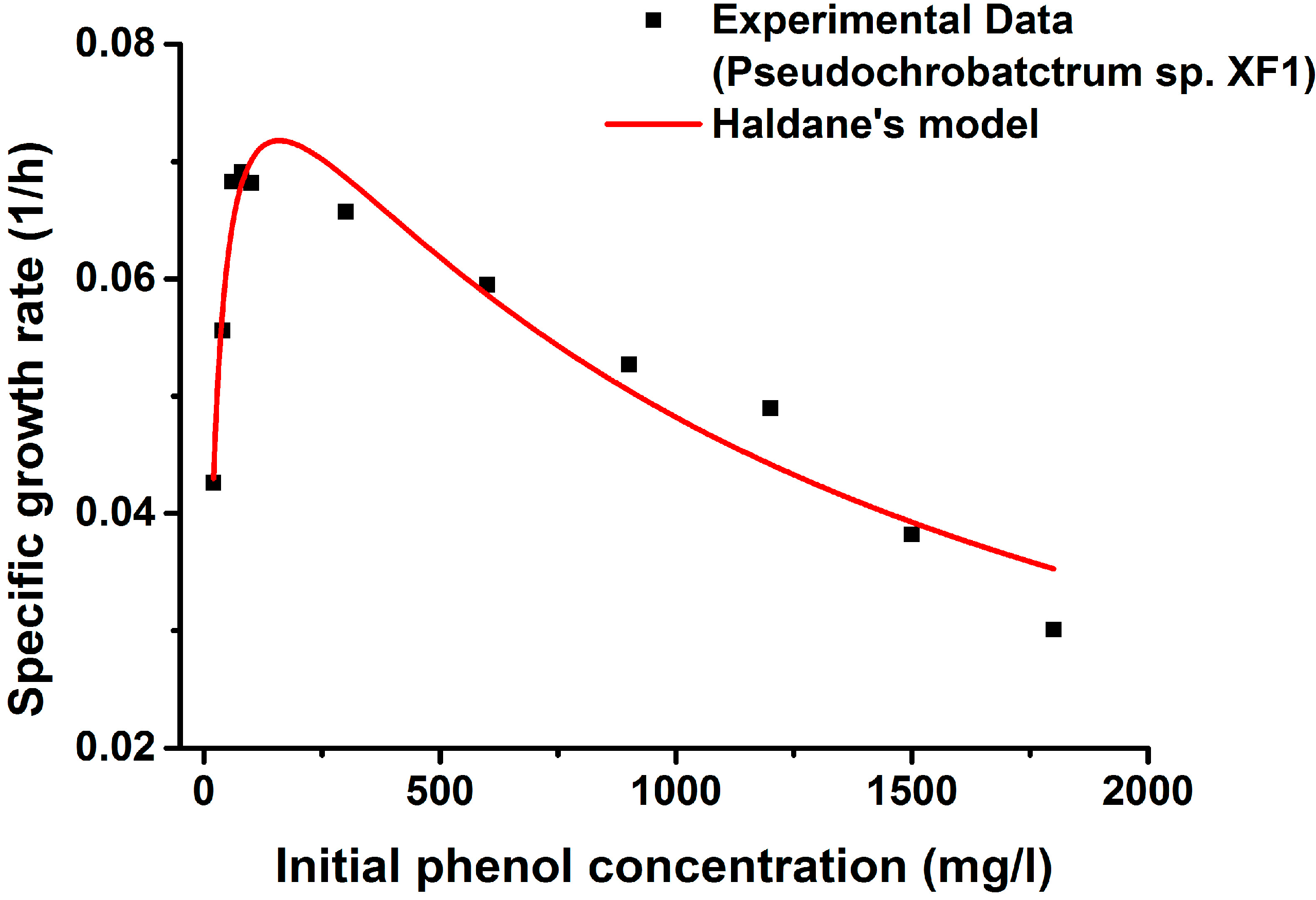
2.4. Kinetics of Phenol Degradation


2.5. Activity of Enzymes in the Phenol Degradation Pathway
| Strain | Specific Activity (U·mg·Protein−1) | ||
|---|---|---|---|
| Phenol Hydroxylase | Catechol 1,2-Dioxygenase | Catechol 2,3-Dioxygenase | |
| XF1 | 0.306 ± 0.009 | 0.324 ± 0.021 | N.D. |
| XF1-UV | 0.761 ± 0.053 | 0.392 ± 0.037 | N.D. |
3. Experimental Section
3.1. Chemicals and Culture Medium
3.2. Selection and Isolation of the Phenol-Degrading Strains
3.3. Identification of the Isolated Bacteria
3.4. Mutagenesis and Screening of Mutants
3.5. Biodegradation Experiments
3.6. Phenol Degradation Kinetics
3.7. Enzyme Assays
3.8. Analytical Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Park, J.S.; Brown, M.T.; Han, T. Phenol toxicity to the aquatic macrophyte Lemna paucicostata. Aquat. Toxicol. 2012, 106, 182–188. [Google Scholar] [CrossRef] [PubMed]
- McCall, I.C.; Betanzos, A.; Weber, D.A.; Nava, P.; Miller, G.W.; Parkos, C.A. Effects of phenol on barrier function of a human intestinal epithelial cell line correlate with altered tight junction protein localization. Toxicol. Appl. Pharmacol. 2009, 241, 61–70. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Dong, Y.M.; Li, Z.; Yin, L.; Zhang, A.M.; Zheng, Y.C. Catalytic ozonation of phenol in water with natural brucite and magnesia. J. Hazard. Mater. 2008, 159, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Yamaguchi, E. Influence of oxalic acid formed on the degradation of phenol by Fenton reagent. Chemosphere 2012, 88, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.H.; Chiou, C.H.; Chang, C.K.; Juang, R.S. Photocatalytic degradation of phenol on different phases of TiO2 particles in aqueous suspensions under UV irradiation. J. Environ. Manag. 2011, 92, 3098–3104. [Google Scholar] [CrossRef]
- Adan, C.; Carbajo, J.; Bahamonde, A.; Martinez-Arias, A. Phenol photodegradation with oxygen and hydrogen peroxide over TiO2 and Fe-doped TiO2. Catal. Today 2009, 143, 247–252. [Google Scholar] [CrossRef]
- Ho, K.L.; Lin, B.; Chen, Y.Y.; Lee, D.J. Biodegradation of phenol using Corynebacterium sp. DJ1 aerobic granules. Bioresour. Technol. 2009, 100, 5051–5055. [Google Scholar] [CrossRef]
- Christen, P.; Davidson, S.; Combet-Blanc, Y.; Auria, R. Phenol biodegradation by the thermoacidophilic archaeon Sulfolobus solfataricus 98/2 in a fed-batch bioreactor. Biodegradation 2011, 22, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Ghoshal, A.K. Phenol degradation by Bacillus cereus: Pathway and kinetic modeling. Bioresour. Technol. 2010, 101, 5501–5507. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.J.; Li, J.W.; Zhang, Z. The characteristics and mechanisms of phenol biodegradation by Fusarium sp. J. Hazard. Mater. 2007, 148, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.C.; Tsai, L.D.; Li, Y.K. An isolated Candida albicans TL3 capable of degrading phenol at large concentration. Biosci. Biotechnol. Biochem. 2005, 69, 2358–2367. [Google Scholar] [CrossRef] [PubMed]
- Agarry, S.E.; Durojaiye, A.O.; Solomon, B.O. Microbial degradation of phenols: A review. Int. J. Environ. Pollut. 2008, 32, 12–28. [Google Scholar] [CrossRef]
- Huang, D.Y.; Zhou, S.G.; Chen, Q.; Zhao, B.; Yuan, Y.; Zhuang, L. Enhanced anaerobic degradation of organic pollutants in a soil microbial fuel cell. Chem. Eng. J. 2011, 172, 647–653. [Google Scholar] [CrossRef]
- Vazquez-Rodriguez, G.A.; Beltran-Hernandez, R.I.; Lucho-Constantino, C.A.; Blasco, J.L. A method for measuring the anoxic biodegradability under denitrifying conditions. Chemosphere 2008, 71, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Kurzbaum, E.; Kirzhner, F.; Sela, S.; Zimmels, Y.; Armon, R. Efficiency of phenol biodegradation by planktonic Pseudomonas pseudoalcaligenes (a constructed wetland isolate) vs. Root and gravel biofilm. Water Res. 2010, 44, 5021–5031. [Google Scholar]
- Li, Y.; Li, J.; Wang, C.; Wang, P.F. Growth kinetics and phenol biodegradation of psychrotrophic Pseudomonas putida LY1. Bioresour. Technol. 2010, 101, 6740–6744. [Google Scholar] [CrossRef] [PubMed]
- Yamaga, F.; Washio, K.; Morikawa, M. Sustainable biodegradation of phenol by Acinetobacter calcoaceticus P23 isolated from the rhizosphere of duckweed Lemna aoukikusa. Environ. Sci. Technol. 2010, 44, 6470–6474. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Zhang, A.N.; Wang, X.C. Biodegradation of phenol by using free and immobilized cells of Acinetobacter sp XA05 and Sphingomonas sp FG03. Biochem. Eng. J. 2009, 44, 187–192. [Google Scholar] [CrossRef]
- Wang, L.M.; Li, Y.; Yu, P.; Xie, Z.X.; Luo, Y.B.; Lin, Y.W. Biodegradation of phenol at high concentration by a novel fungal strain Paecilomyces variotii JH6. J. Hazard. Mater. 2010, 183, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, Q.J.; Wang, G.X.; Ye, F.; Cong, Y.Y. Biodegradation of phenol at high concentration by a novel yeast Trichosporon montevideense PHE1. Process. Biochem. 2011, 46, 1678–1681. [Google Scholar] [CrossRef]
- Basak, B.; Bhunia, B.; Dutta, S.; Chakraborty, S.; Dey, A. Kinetics of phenol biodegradation at high concentration by a metabolically versatile isolated yeast Candida tropicalis PHB5. Environ. Sci. Pollut. Res. 2014, 21, 1444–1454. [Google Scholar] [CrossRef]
- Leitao, A.L.; Duarte, M.P.; Oliveira, J.S. Degradation of phenol by a halotolerant strain of Penicillium chrysogenum. Int. Biodeterior. Biodegrad. 2007, 59, 220–225. [Google Scholar] [CrossRef]
- Parekh, S.; Vinci, V.A.; Strobel, R.J. Improvement of microbial strains and fermentation processes. Appl. Microbiol. Biotechnol. 2000, 54, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.M.; Inamdar, S.A.; Jadhav, J.P.; Govindwar, S.P. Random UV mutagenesis approach for enhanced biodegradation of sulfonated azo dye, green HE4B. Appl. Biochem. Biotechnol. 2013, 169, 1467–1481. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Liu, L.F.; Ouml;sterlund, T.; Hou, J.; Huang, M.T.; Fagerberg, L.; Petranovic, D.; Uhlen, M.; Nielsen, J. Improved production of a heterologous amylase in Saccharomyces cerevisiae by inverse metabolic engineering. Appl. Environ. Microbiol. 2014, 80, 5542–5550. [Google Scholar] [CrossRef] [PubMed]
- Heerd, D.; Tari, C.; Fernandez-Lahore, M. Microbial strain improvement for enhanced polygalacturonase production by Aspergillus sojae. Appl. Microbiol. Biotechnol. 2014, 98, 7471–7481. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, P.; Huber, B.; Lodders, N.; Warfolomeow, I.; Busse, H.J.; Scholz, H.C. Pseudochrobactrum lubricantis sp nov., isolated from a metal-working fluid. Int. J. Syst. Evol. Microbiol. 2009, 59, 2464–2467. [Google Scholar] [CrossRef]
- Kampfer, P.; Rossello-Mora, R.; Scholz, H.C.; Welinder-Olsson, C.; Falsen, E.; Busse, H.J. Description of Pseudochrobactrum gen. Nov., with the two species Pseudochrobactrum asaccharolyticum sp. nov and Pseudochrobactrum saccharolyticum sp nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 1823–1829. [Google Scholar]
- Long, D.Y.; Tang, X.J.; Cai, K.; Chen, G.C.; Shen, C.F.; Shi, J.Y.; Chen, L.G.; Chen, Y.X. Cr(vi) resistance and removal by indigenous bacteria isolated from chromium-contaminated soil. J. Microbiol. Biotechnol. 2013, 23, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Dong, X.; Zhou, J.; Ge, S. Comparative evaluations on bio-treatment of hexavalent chromate by resting cells of Pseudochrobactrum sp. and Proteus sp. In wastewater. J. Environ. Manag. 2013, 126, 7–12. [Google Scholar]
- Chandra, R.; Singh, R. Decolourisation and detoxification of rayon grade pulp paper mill effluent by mixed bacterial culture isolated from pulp paper mill effluent polluted site. Biochem. Eng. J. 2012, 61, 49–58. [Google Scholar] [CrossRef]
- Liu, J.Z.; Wang, Q.; Yan, J.B.; Qin, X.R.; Li, L.L.; Xu, W.; Subramaniam, R.; Bajpai, R.K. Isolation and characterization of a novel phenol degrading bacterial strain WUST-C1. Ind. Eng. Chem. Res. 2013, 52, 258–265. [Google Scholar]
- Paisio, C.E.; Talano, M.A.; Gonzalez, P.S.; Busto, V.D.; Talou, J.R.; Agostini, E. Isolation and characterization of a Rhodococcus strain with phenol-degrading ability and its potential use for tannery effluent biotreatment. Environ. Sci. Pollut. Res. 2012, 19, 3430–3439. [Google Scholar] [CrossRef]
- Mohite, B.V.; Pawar, S.P.; Morankar, A. Isolation, selection and biodegradation profile of phenol degrading bacteria from oil contaminated soil. Bull. Environ. Contam. Toxicol. 2011, 87, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Q.Y.; Huang, T.P.; Zhang, Y.K.; Ding, R.F. Enhanced bioremediation of soil contaminated with viscous oil through microbial consortium construction and ultraviolet mutation. World J. Microbiol. Biotechnol. 2011, 27, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Sivaprakasam, S.; Mahadevan, S. Investigations on ultraviolet light and nitrous acid induced mutations of halotolerant bacterial strains for the treatment of tannery soak liquor. Int. Biodeterior. Biodegrad. 2009, 63, 176–181. [Google Scholar] [CrossRef]
- Kim, D.J.; Choi, J.W.; Choi, N.C.; Mahendran, B.; Lee, C.E. Modeling of growth kinetics for Pseudomonas spp. during benzene degradation. Appl. Microbiol. Biotechnol. 2005, 69, 456–462. [Google Scholar] [CrossRef]
- Wei, G.; Yu, J.; Zhu, Y.; Chen, W.; Wang, L. Characterization of phenol degradation by Rhizobium sp CCNWTB 701 isolated from Astragalus chrysopteru in mining tailing region. J. Hazard. Mater. 2008, 151, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Ghoshal, A.K. Isolation and characterization of hyper phenol tolerant Bacillus sp. from oil refinery and exploration sites. J. Hazard. Mater. 2010, 176, 85–91. [Google Scholar]
- Adsul, M.G.; Bastawde, K.B.; Varma, A.J.; Gokhale, D.V. Strain improvement of Penicillium janthinellum NCIM 1171 for increased cellulase production. Bioresour. Technol. 2007, 98, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.J.; Hong, Q.; He, L.J.; Jiang, X.; Li, S.P. Characterization of phenol-degrading bacterial strains isolated from natural soil. Int. Biodeterior. Biodegrad. 2008, 62, 257–262. [Google Scholar] [CrossRef]
- Alexievaa, Z.; Gerginova, M.; Zlateva, P.; Peneva, N. Comparison of growth kinetics and phenol metabolizing enzymes of Trichosporon cutaneum R57 and mutants with modified degradation abilities. Enzym. Microb. Technol. 2004, 34, 242–247. [Google Scholar] [CrossRef]
- Pradhan, N.; Ingle, A.O. Mineralization of phenol by a Serratia plymuthica strain GC isolated from sludge sample. Int. Biodeterior. Biodegrad. 2007, 60, 103–108. [Google Scholar] [CrossRef]
- Kämpfer, P.; Steiof, M.; Dott, W. Microbiological characterization of a fuel-oil contaminated site including numerical identification of heterotrophic water and soil bacteria. Microb. Ecol. 1991, 21, 227–251. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Shin, Y.K.; Ueda, Y.; Sugiyama, J. Automated sequencing of PCR-amplified 16s rdna on “hydrolink gels”. J. Microbiol. Methods 1994, 19, 145–154. [Google Scholar] [CrossRef]
- Gaal, A.; Neujahr, H.Y. Metabolism of phenol and resorcinol in Trichosporon cutaneum. J. Bacteriol. 1979, 137, 13–21. [Google Scholar] [PubMed]
- Neujahr, H.Y.; Gaal, A. Phenol hydroxylase from yeast. Eur. J. Biochem. 1973, 35, 386–400. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.M.; Neujahr, H.Y. Purification and properties of catechol 1,2-oxygenase from Trichosporon cutaneum. Eur. J. Biochem. 1970, 12, 427–434. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Z.; Yu, C.; Xin, L. Enhancement of Phenol Biodegradation by Pseudochrobactrum sp. through Ultraviolet-Induced Mutation. Int. J. Mol. Sci. 2015, 16, 7320-7333. https://doi.org/10.3390/ijms16047320
Mao Z, Yu C, Xin L. Enhancement of Phenol Biodegradation by Pseudochrobactrum sp. through Ultraviolet-Induced Mutation. International Journal of Molecular Sciences. 2015; 16(4):7320-7333. https://doi.org/10.3390/ijms16047320
Chicago/Turabian StyleMao, Zhen, Chenyang Yu, and Lingling Xin. 2015. "Enhancement of Phenol Biodegradation by Pseudochrobactrum sp. through Ultraviolet-Induced Mutation" International Journal of Molecular Sciences 16, no. 4: 7320-7333. https://doi.org/10.3390/ijms16047320