
Potential Effect of CD271 on Human Mesenchymal Stromal Cell Proliferation and Differentiation


 ,
,  , , ,
, , ,
Abstract
:
1. Introduction
2. Results and Discussion
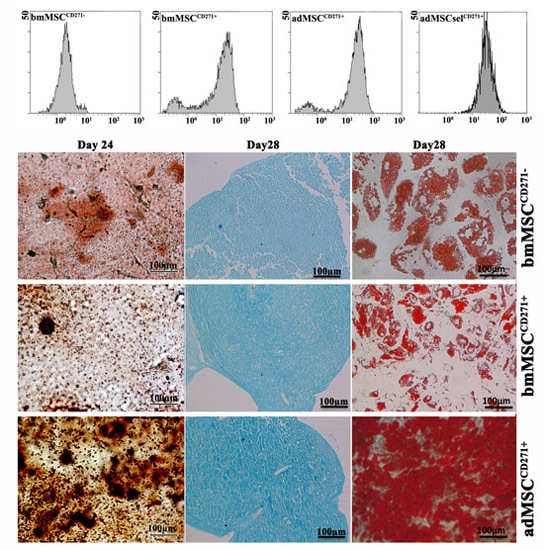
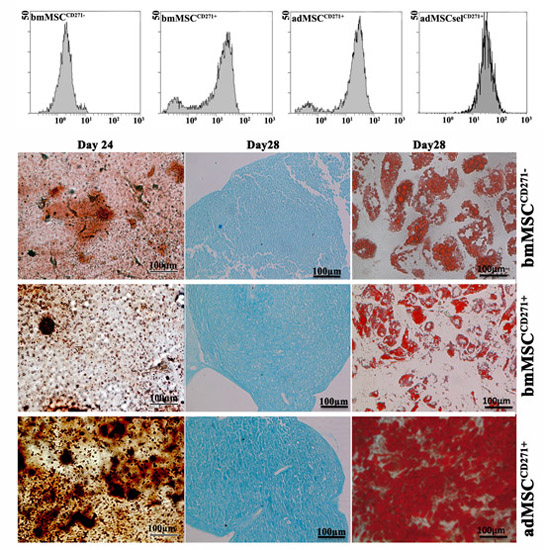
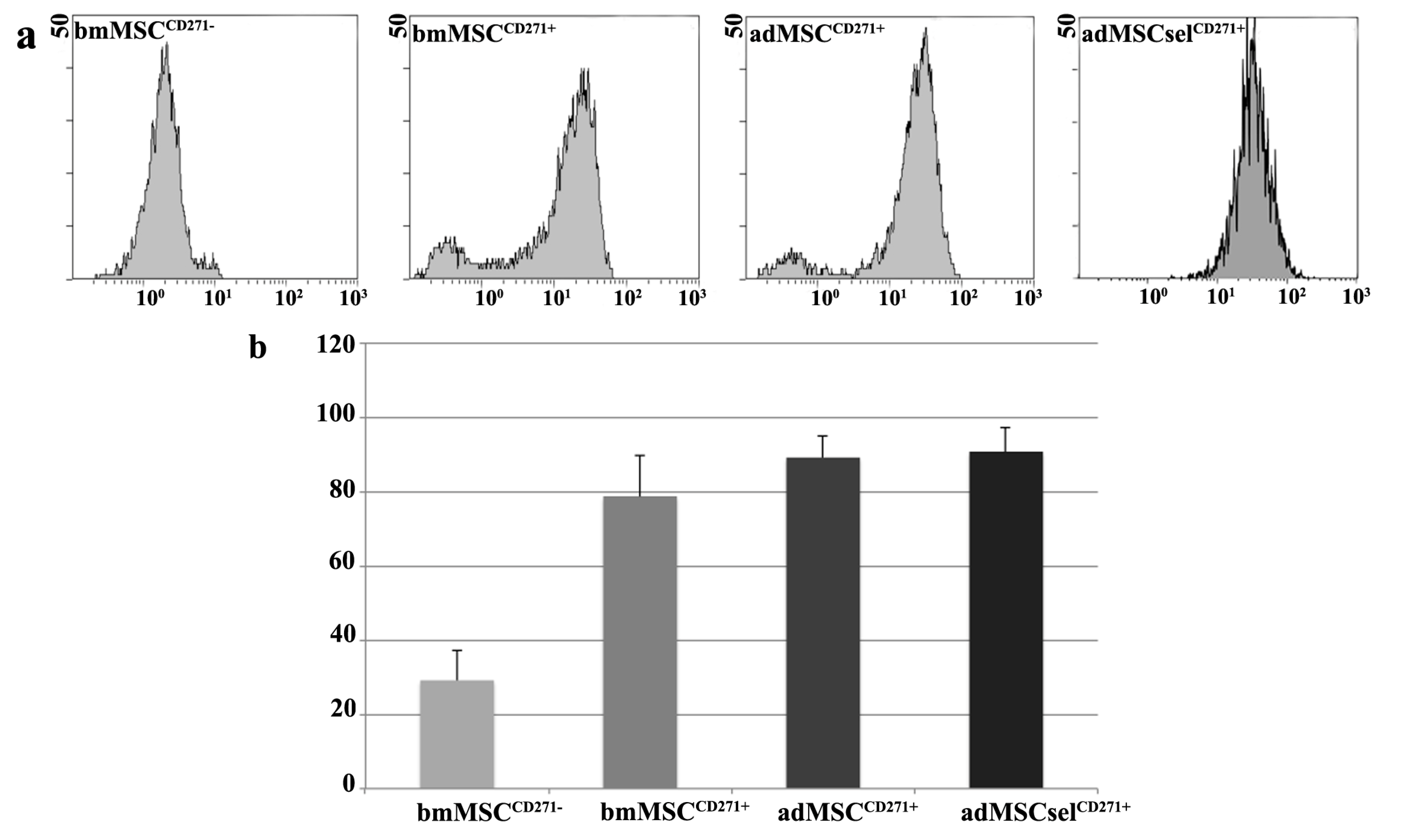
2.1. Phenotypic Characterization of Mesenchymal Stem Cells (MSCs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | bmMSCCD271− (Mean ± s.e.m.) | bmMSCCD271+ (Mean ± s.e.m.) | adMSCCD271+ (Mean ± s.e.m.) | adMSCselCD271+ (Mean ± s.e.m.) | One-Way ANOVA | |
|---|---|---|---|---|---|---|
| p | p-Corrected | |||||
| CD31 | 2.45 ± 0.55 | 7.08 ± 3.74 | 2.23 ± 0.92 | 5.13 ± 2.09 | 0.403 | 1.000 |
| CD45 | 2.30 ± 0.60 | 4.21 ± 1.67 | 2.93 ± 2.25 | 2.4 ± 1.6 | 0.824 | 1.000 |
| CD34 | 3.44 ± 2.10 | 5.17 ± 2.28 | 3.33 ± 1.94 | 3.83 ± 2.63 | 0.933 | 1.000 |
| CD90 | 88.13 ± 0.79 | 96.10 ± 2.41 | 87.00 ± 6.21 | 92.3 ± 6.17 | 0.513 | 1.000 |
| CD73 | 89.30 ± 2.72 | 82.33 ± 6.08 | 95.10 ± 1.75 | 94.13 ± 0.37 | 0.104 | 1.000 |
| CD105 | 88.10 ± 2.72 | 85.85 ± 6.08 | 88.43 ± 1.75 | 91.93 ± 1.68 | 0.77 | 1.000 |
| CD271 | 29.13 ± 8.18 | 78.83 ± 10.89 | 89.20 ± 5.66 | 90.57 ± 4.16 | 0.001 ** | 0.01 ** |
| Tukey’s post-hoc | ||||||
| Sample Types | diff | ler | upr | p | ||
| adMSCselCD271+ vs. adMSCCD271+ | 1.366667 | −33.32805 | 36.06138 | 0.999 | ||
| bmMSCCD271− vs. adMSCCD271+ | −60.066667 | −94.76138 | −25.37195 | 0.002 ** | ||
| bmMSCCD271+ vs. adMSCCD271+ | −10.373333 | −45.06805 | 24.32138 | 0.776 | ||
| bmMSCCD271− vs. adMSCselCD271+ | −61.433333 | −96.12805 | −26.73862 | 0.002 ** | ||
| bmMSCCD271+ vs. adMSCselCD271+ | −11.740000 | −46.43471 | 22.95471 | 0.708 | ||
| bmMSCCD271+ vs. bmMSCCD271− | 49.693333 | 14.99862 | 84.38805 | 0.007 ** | ||

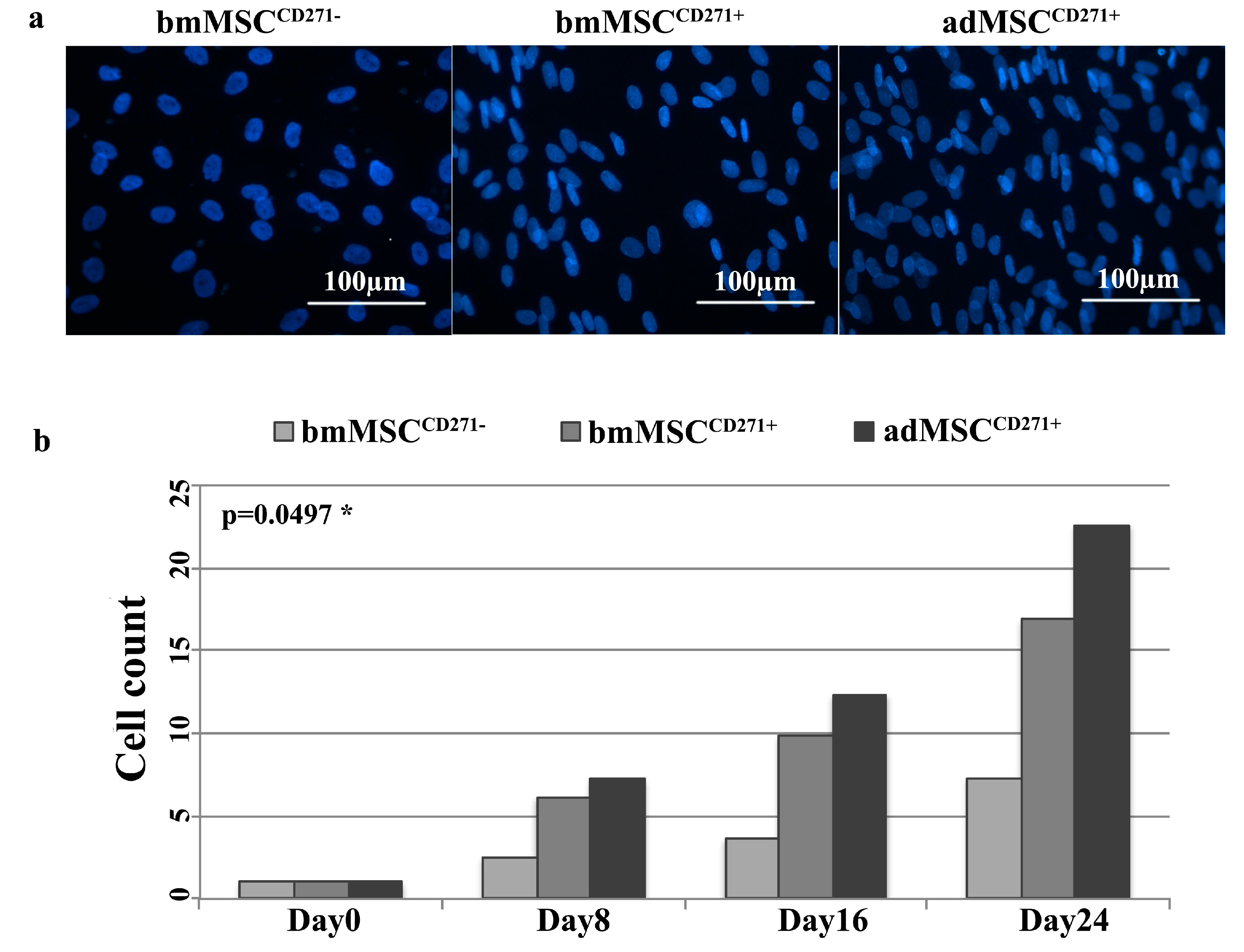
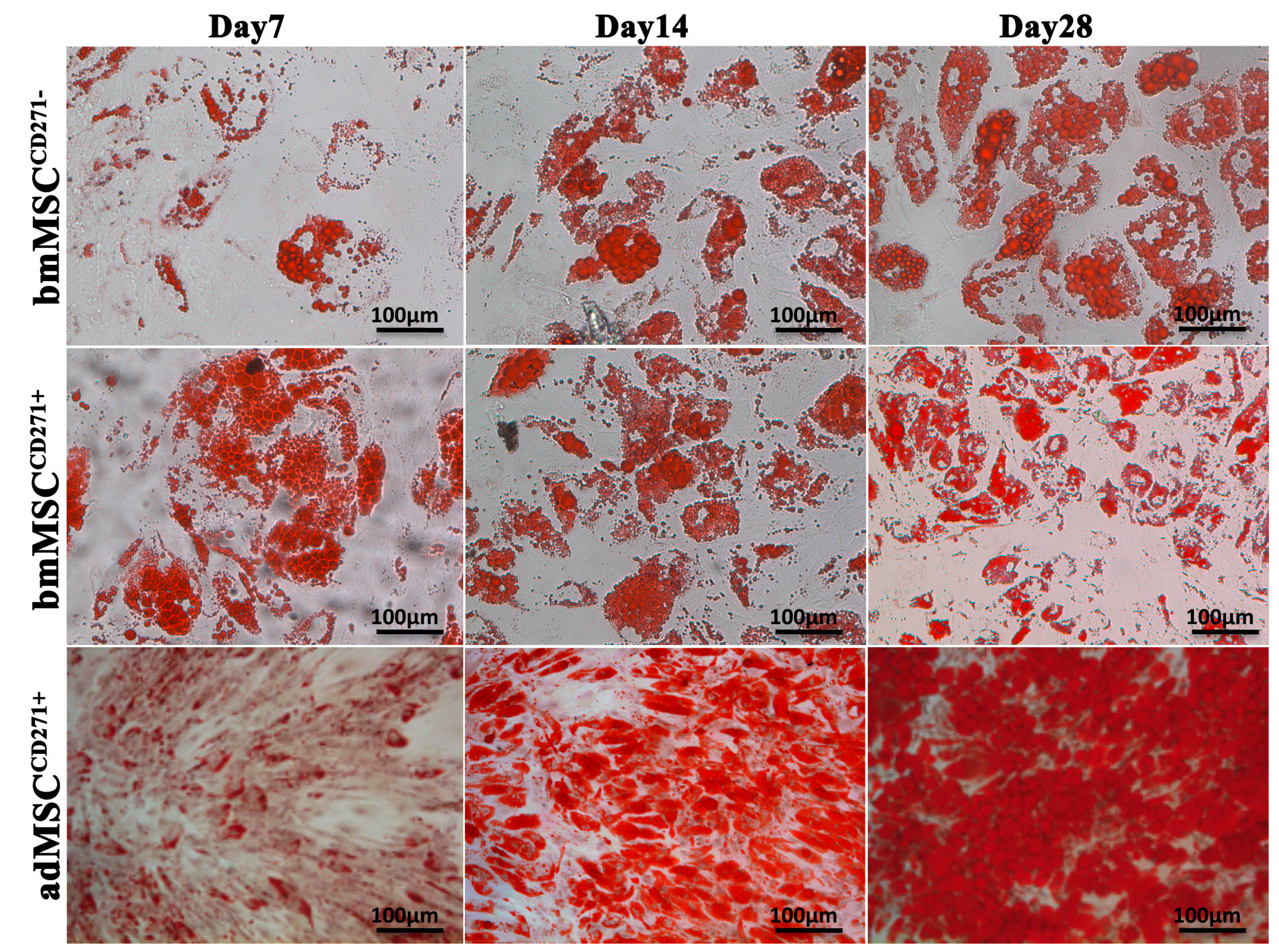
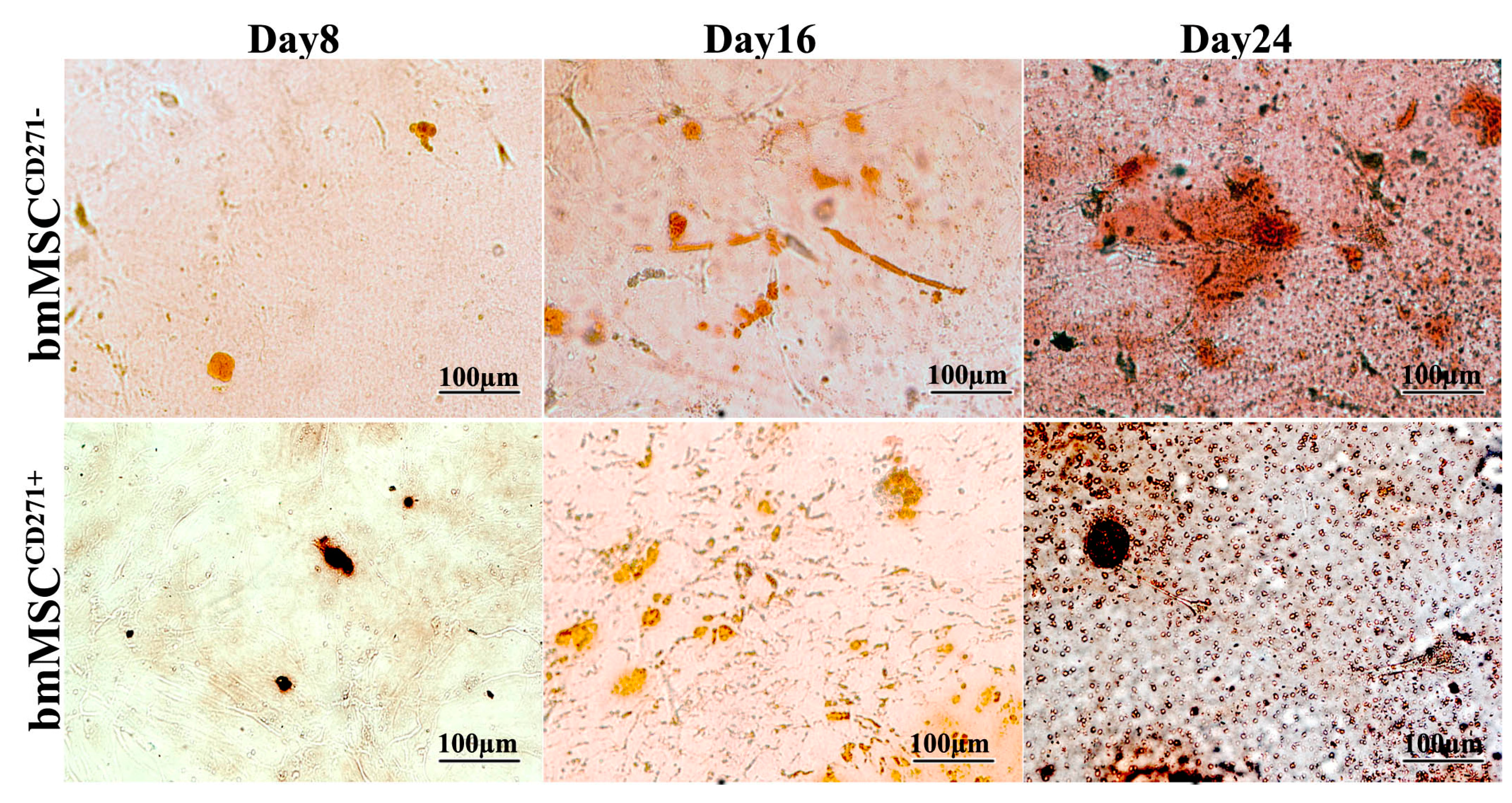
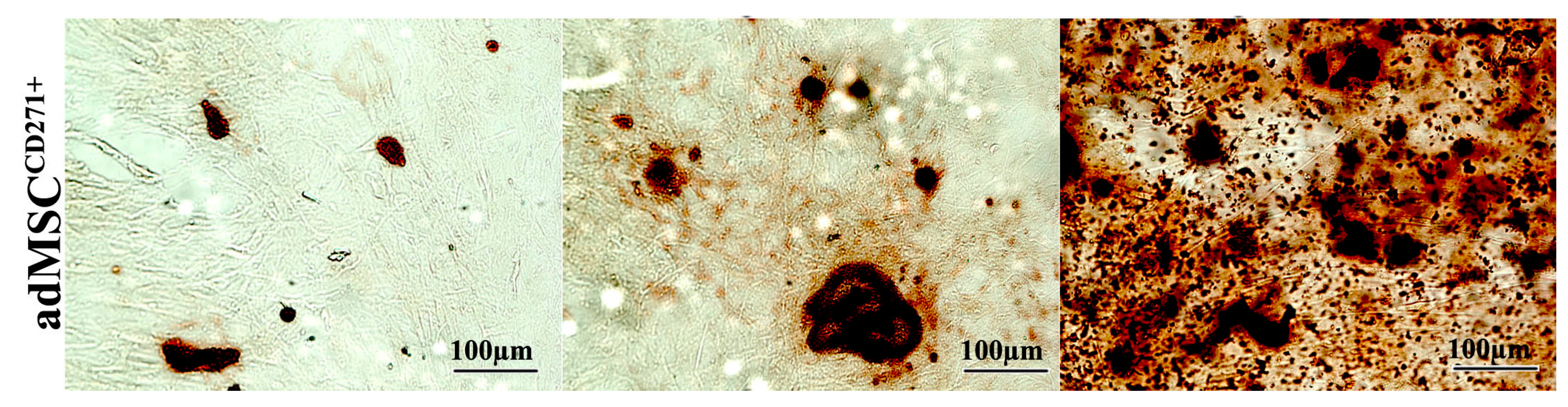
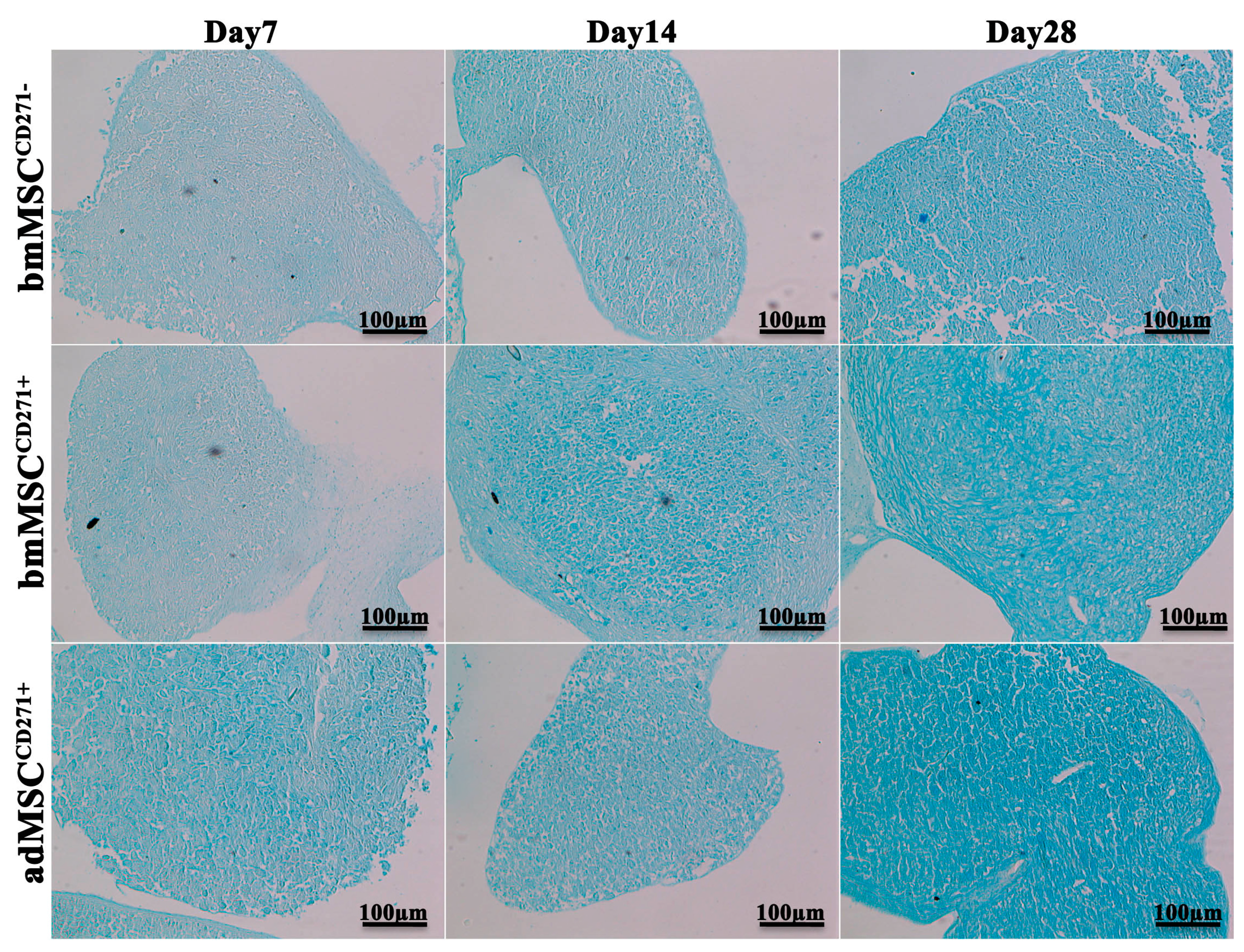
2.2. The Expression of CD271 by MSCs Could Increase Proliferative and Trilineage Differentiation Potential





3. Experimental Section
3.1. Isolation, Expansion and Characterization of MSCs Derived from Adipose Tissue/Lipoaspirate and Bone Marrow
| Donor | Age | Sex | Sources | Disease | Diagnosis | Follow-up |
|---|---|---|---|---|---|---|
| BM_1 | 66 | M | Aspirate | CHL | Post-chemiotherapy | Healthy |
| BM_2 | 70 | M | Aspirate | CML | Post-chemiotherapy | Healthy |
| BM_3 | 64 | F | Aspirate | CML | Onset | Healthy |
| AD_1 | 62 | M | Kidney | – | No chemiotherapy | Healthy |
| AD_2 | 61 | F | Breast | – | No chemiotherapy | Healthy |
| AD_3 | 60 | F | Colon | – | No chemiotherapy | Healthy |
3.2. Proliferative and Trilineage Differentiation Potential of CD271+-MSCs and CD271−-MSCs. Proliferation Assay Was Carried out by DAPI and Trypan Blue Staining
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Tissue Kinet. 1970, 3, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; de Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Dicker, A.; le Blanc, K.; Aström, G.; van Harmelen, V.; Götherström, C.; Blomqvist, L.; Amer, P.; Ryden, M. Functional studies of mesenchymal stem cells derived from adult human adipose tissue. Exp. Cell Res. 2005, 308, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Zhu, X.Y.; Krier, J.D.; Tang, H.; Jordan, K.L.; Grande, J.P.; Lerman, A.; Textor, S.C.; Lerman, L.O. Adipose tissue-derived mesenchymal stem cells improve revascularization outcomes to restore renal function in swine atherosclerotic renal artery stenosis. Stem Cells 2012, 30, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Campagnoli, C.; Roberts, I.A.G.; Kumar, S.; Bennett, P.R.; Bellantuono, I.; Fisk, N.M. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 2001, 98, 2396–2402. [Google Scholar] [CrossRef] [PubMed]
- In’t Anker, P.S.; Noort, W.A.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Kruisselbrink, A.B.; van Bezooijen, R.L.; Beekhuizen, W.; Willemze, R.; Kanhai, H.H.; Fibbe, W.E. Mesenchymal stem cells in human second-trimester bone marrow, liver, lung, and spleen exhibit a similar immunophenotype but a heterogeneous multilineage differentiation potential. Haematologica 2003, 88, 845–852. [Google Scholar]
- In’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 2003, 102, 1548–1549. [Google Scholar] [CrossRef] [PubMed]
- Bieback, K.; Kluter, H. Mesenchymal stromal cells from umbilical cord blood. Curr. Stem Cell Res. Ther. 2007, 2, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.; Xu, W.; Zhu, W.; Hu, J.; Qian, H.; Yin, Q.; Jiang, R.; Yan, Y.; Mao, F.; Yang, H.; et al. Human mesenchymal stem cells isolated from the umbilical cord. Cell Biol. Int. 2008, 32, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Secco, M.; Zucconi, E.; Vieira, N.M.; Fogaça, L.L.; Cerqueira, A.; Carvalho, M.D.; Jazedje, T.; Okamoto, O.K.; Muotri, A.R.; Zatz, M. Mesenchymal stem cells from umbilical cord: Do not discard the cord! Neuromuscul. Disord. 2008, 18, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Sarugaser, R.; Lickorish, D.; Baksh, D.; Hosseini, M.M.; Davies, J.E. Human umbilical cord perivascular (HUCPV) cells: A source of mesenchymal progenitors. Stem Cells 2005, 23, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Martin-Rendon, E.; Sweeney, D.; Lu, F.; Girdlestone, J.; Navarrete, C.; Watt, S.M. 5-Azacytidine-treated human mesenchymal stem/progenitor cells derived from umbilical cord, cord blood and bone marrow do not generate cardiomyocytes in vitro at high frequencies. Vox Sang. 2008, 95, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Jin, J.; Chen, L.; Zhu, J.; Huang, W.; Zhao, J.; Qian, H.; Zhang, X. Isolation of mesenchymal stem cells from human placenta: Comparison with human bone marrow mesenchymal stem cells. Cell Biol. Int. 2006, 30, 681–687. [Google Scholar] [CrossRef] [PubMed]
- In’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; de Groot-Swings, G.M.; Claas, F.H.; Fibbe, W.E.; Kanhai, H.H. Isolation of mesenchymal stem cells of fetal or maternal origin from human placenta. Stem Cells 2004, 22, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Fukuchi, Y.; Nakajima, H.; Sugiyama, D.; Hirose, I.; Kitamura, T.; Tsuji, K. Human placenta-derived cells have mesenchymal stem/progenitor cell potential. Stem Cells 2004, 22, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Schwab, K.E.; Gargett, C.E. Co-expression of two perivascular cell markers isolates mesenchymal stem-like cells from human endometrium. Hum. Reprod. 2007, 22, 2903–2911. [Google Scholar] [CrossRef] [PubMed]
- Schwab, K.E.; Hutchinson, P.; Gargett, C.E. Identification of surface markers for prospective isolation of human endometrial stromal colony-forming cells. Hum. Reprod. 2008, 23, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Pierdomenico, L.; Bonsi, L.; Calvitti, M.; Rondelli, D.; Arpinati, M.; Chirumbolo, G.; Becchetti, E.; Marchionni, C.; Alviano, F.; Fossati, V.; et al. Multipotent mesenchymal stem cells with immunosuppressive activity can be easily isolated from dental pulp. Transplantation 2005, 8, 836–842. [Google Scholar] [CrossRef]
- Huang, G.T.; Gronthos, S.; Shi, S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in regenerative medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.L.; Beare, A.; Zola, H.; Rush, R.A. CD271 (P75 neurotrophin receptor). J. Biol. Regul. Homeost. Agents 2008, 22, 1–6. [Google Scholar] [PubMed]
- Casaccia-Bonnefil, P.; Gu, C.; Chao, M.V. Neurotrophins in cell survival/death decisions. Adv. Exp. Med. Biol. 1999, 468, 275–282. [Google Scholar] [PubMed]
- Yan, H.; Schlessinger, J.; Chao, M.V. Chimeric NGF-EGF receptors define domains responsible for neuronal differentiation. Science 1991, 252, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Sailer, M.H.; Gerber, A.; Tostado, C.; Hutter, G.; Cordier, D.; Mariani, L.; Ritz, M.F. Non-invasive neural stem cells become invasive in vitro by combined FGF2 and BMP4 signaling. J. Cell Sci. 2013, 126, 3533–3540. [Google Scholar] [CrossRef] [PubMed]
- Boiko, A.D.; Razorenova, O.V.; van de Rijn, M.; Swetter, S.M.; Johnson, D.L.; Ly, D.P.; Butler, P.D.; Yang, G.P.; Joshua, B.; Kaplan, M.J.; et al. Human melanoma-initiating cells express neural crest nerve growth factor receptor CD271. Nature 2010, 466, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Civenni, G.; Walter, A.; Kobert, N.; Mihic-Probst, D.; Zipser, M.; Belloni, B.; Seifert, B.; Moch, H.; Dummer, R.; van den Broek, M.; et al. Human CD271-positive melanoma stem cells associated with metastasis establish tumor heterogeneity and long-term growth. Cancer Res. 2011, 71, 3098–3109. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.D.; Yuan, Y.; Liu, X.H.; Gong, D.J.; Bai, C.G.; Wang, F.; Luo, J.H.; Xu, Z.Y. Self-renewal and chemotherapy resistance of p75NTR positive cells in esophageal squamous cell carcinomas. BMC Cancer 2009, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Okumura, T.; Shimada, Y.; Imamura, M.; Yasumoto, S. Neurotrophin receptor p75(NTR) characterizes human esophageal keratinocyte stem cells in vitro. Oncogene 2003, 22, 4017–4026. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Tamai, K.; Oizumi, S.; Oyama, K.; Yamaguchi, K.; Sato, I.; Satoh, K.; Matsuura, K.; Saijo, S.; Sugamura, K.; et al. CD271 defines a stem cell-like population in hypopharyngeal cancer. PLoS ONE 2013, 8, e62002. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Sauca, O.; Chung, M.K.; Shin, J.H.; Karamboulas, C.; Kwok, S.; Jung, Y.H.; Oakley, R.; Tysome, J.R.; Farnebo, L.O.; Kaplan, M.J.; et al. CD271 is a functional and targetable marker of tumor-initiating cells in head and neck squamous cell carcinoma. Oncotarget 2014, 5, 6854–6866. [Google Scholar] [PubMed]
- Kuçi, Z.; Seiberth, J.; Latifi-Pupovci, H.; Wehner, S.; Stein, S.; Grez, M.; Bönig, H.; Köhl, U.; Klingebiel, T.; Bader, P.; et al. Clonal analysis of multipotent stromal cells derived from CD271+ bone marrow mononuclear cells: Functional heterogeneity and different mechanisms of allosuppression. Haematologica 2013, 98, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Kuçi, S.; Kuçi, Z.; Kreyenberg, H.; Deak, E.; Pütsch, K.; Huenecke, S.; Amara, C.; Koller, S.; Rettinger, E.; Grez, M.; et al. CD271 antigen defines a subset of multipotent stromal cells with immunosuppressive and lymphohematopoietic engraftment-promoting properties. Haematologica 2010, 95, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Viejo, M.; Menendez-Menendez, Y.; Blanco-Gelaz, M.A.; Ferrero-Gutierrez, A.; Fernandez-Rodriguez, M.A.; Perez-Basterrechea, M.; Gracia-Gala, J.M.; Perez-Lopez, S. Otero-Hernandez, J. LNGFR (CD271) as marker to identify mesenchymal stem cells from different human sources: Umbilical cord blood, Wharton’s Jelly and bone marrow. J. Bone Marrow Res. 2013, 1, 1000132. [Google Scholar]
- Lv, F.J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. The surface markers and identity of human mesenchymal stem cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Strioga, M.; Viswanathan, S.; Darinskas, A.; Slaby, O.; Michalek, J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012, 21, 2724–2752. [Google Scholar] [CrossRef] [PubMed]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef]
- Poloni, A.; Maurizi, G.; Rosini, V.; Mondini, E.; Mancini, S.; Discepoli, G.; Biasio, S.; Battaglini, G.; Felicetti, S.; Berardinelli, E.; et al. Selection of CD271+ cells and human AB serum allows a large expansion of mesenchymal stromal cells from human bone marrow. Cytotherapy 2009, 11, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Kermani, P.; Teng, K.K.; Hempstead, B.L. Regulation of cell survival by secreted proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Massa, S.M.; Xie, Y.; Yang, T.; Harrington, W.A.; Kim, M.L.; Yoon, S.O.; Kraemer, R.; Moore, L.A.; Hempstead, B.L.; Longo, F.M. Small, nonpeptide p75NTR ligands induce survival signaling and inhibit proNGF-induced death. J. Neurosci. 2006, 26, 5288–300. [Google Scholar] [CrossRef] [PubMed]
- Moscatelli, I.; Pierantozzi, E.; Camaioni, A.; Siracusa, G.; Campagnolo, L. p75 neurotrophin receptor is involved in proliferation of undifferentiated mouse embryonic stem cells. Exp. Cell Res. 2009, 315, 3220–3232. [Google Scholar] [CrossRef] [PubMed]
- Cragnolini, A.B.; Friedman, W.J. The function of p75NTR in glia. Trends Neurosci. 2008, 31, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Tomellini, E.; Lagadec, C.; Polakowska, R.; le Bourhis, X. Role of p75 neurotrophin receptor in stem cell biology: More than just a marker. Cell. Mol. Life Sci. 2014, 71, 2467–2481. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Li, X.; Si, M.; Liu, T.; Li, J. CD271+ osteosarcoma cells display stem-like properties. PLoS ONE 2014, 9, e98549. [Google Scholar] [CrossRef] [PubMed]
- Cheli, Y.; Bonnazi, V.F.; Jacquel, A.; Allegra, M.; de Donatis, G.M.; Bahadoran, P.; Bertolotto, C.; Ballotti, R. CD271 is an imperfect marker for melanoma initiating cells. Oncotarget 2014, 5, 5272–5283. [Google Scholar] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Jahagirdar, B.N.; Reinhardt, R.L.; ScHwartz, R.E.; Keene, C.D.; Ortiz-Gonzalez, X.R.; Reyes, M.; Lenvik, T.; Lund, T.; Blackstad, M.; et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Strem, B.M.; Hicok, K.C.; Zhu, M.; Wulur, I.; Alfonso, Z.; Schreiber, R.E.; Fraser, J.K.; Hedrick, M.H. Multipotential differentiation of adipose tissue-derived stem cells. Keio J. Med. 2005, 54, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Akamatsu, H.; Hasegawa, S.; Yamada, T.; Nakata, S.; Ohkuma, M.; Miyachi, E.; Marunouchi, T.; Matsunaga, K. Isolation of multipotent stem cells from mouse adipose tissue. J. Dermatol. Sci. 2007, 48, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Guo, Z.; Jiang, X.; Li, H.; Wang, X.; Yao, H.; Zhang, Y.; Mao, N. A protocol for isolation and culture of mesenchymal stem cells from mouse compact bone. Nat. Protoc. 2010, 5, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Floyd, Z.E.; Wu, X.; Halvorsen, Y.D.; Gimble, J.M. Isolation of human adipose-derived stem cells from lipoaspirates. In Adipose-Derived Stem Cells: Methods and Protocols; Gimble, J.M., Bunnell, B.A., Eds.; Springer: New York, NY, USA, 2011; Volume 702, pp. 17–27. [Google Scholar]
- Chambers, J.M.; Freeny, A.; Heiberger, R.M. Analysis of variance; designed experiments. In Chapter 5 of Statistical Models in S; Chambers, J.M., Hastie, T.J., Eds.; Wadsworth & Brooks/Cole; Springer: New York, NY, USA, 1992. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2015. Available online: http://www.R-project.org/ (accessed on 20 May 2015).
- Miller, R.G. Simultaneous Statistical Inference; Springer: New York, NY, USA, 1981. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calabrese, G.; Giuffrida, R.; Lo Furno, D.; Parrinello, N.L.; Forte, S.; Gulino, R.; Colarossi, C.; Schinocca, L.R.; Giuffrida, R.; Cardile, V.; et al. Potential Effect of CD271 on Human Mesenchymal Stromal Cell Proliferation and Differentiation. Int. J. Mol. Sci. 2015, 16, 15609-15624. https://doi.org/10.3390/ijms160715609
Calabrese G, Giuffrida R, Lo Furno D, Parrinello NL, Forte S, Gulino R, Colarossi C, Schinocca LR, Giuffrida R, Cardile V, et al. Potential Effect of CD271 on Human Mesenchymal Stromal Cell Proliferation and Differentiation. International Journal of Molecular Sciences. 2015; 16(7):15609-15624. https://doi.org/10.3390/ijms160715609
Chicago/Turabian StyleCalabrese, Giovanna, Raffaella Giuffrida, Debora Lo Furno, Nunziatina Laura Parrinello, Stefano Forte, Rosario Gulino, Cristina Colarossi, Luciana Rita Schinocca, Rosario Giuffrida, Venera Cardile, and et al. 2015. "Potential Effect of CD271 on Human Mesenchymal Stromal Cell Proliferation and Differentiation" International Journal of Molecular Sciences 16, no. 7: 15609-15624. https://doi.org/10.3390/ijms160715609
APA StyleCalabrese, G., Giuffrida, R., Lo Furno, D., Parrinello, N. L., Forte, S., Gulino, R., Colarossi, C., Schinocca, L. R., Giuffrida, R., Cardile, V., & Memeo, L. (2015). Potential Effect of CD271 on Human Mesenchymal Stromal Cell Proliferation and Differentiation. International Journal of Molecular Sciences, 16(7), 15609-15624. https://doi.org/10.3390/ijms160715609