
The Role of CzcRS Two-Component Systems in the Heavy Metal Resistance of Pseudomonas putida X4


Abstract
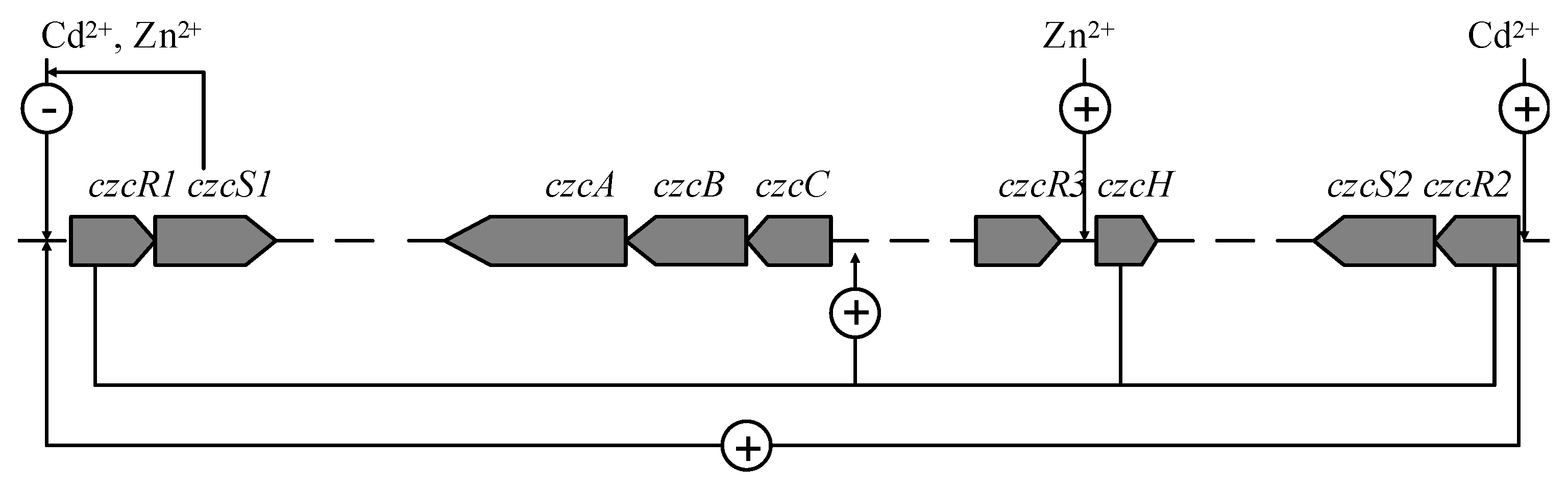
:1. Introduction
2. Results
2.1. Minimum Inhibitory Concentrations (MICs) of the Wild-Type and Mutant Strains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (mmol·L−1) a | |||
|---|---|---|---|---|
| Cd2+ | Co2+ | Cu2+ | Zn2+ | |
| X4 | 4.5 | 4.0 | 5.0 | 8.5 |
| X4 with pvlt31 | 4.5 | 4.0 | 5.0 | 8.5 |
| ΔS1 | 3.0 | 0.5 | 4.25 | 2.25 |
| ΔS2 | 2.0 | 1.5 | 4.75 | 8.5 |
| ΔczcH | 2.0 | 1.75 | 5.0 | 7.0 |
| ΔS1S2 | 1.25 | 0.5 | 4.25 | 5.0 |
| ΔS1c | 4.25 | 3.5 | 4.5 | 7.5 |
| ΔS2c | 4.25 | 4.0 | 4.75 | 8.5 |
| ΔczcHc | 3.75 | 3.5 | 5.0 | 8.25 |
2.2. Genetic Complementation of Mutant Genes
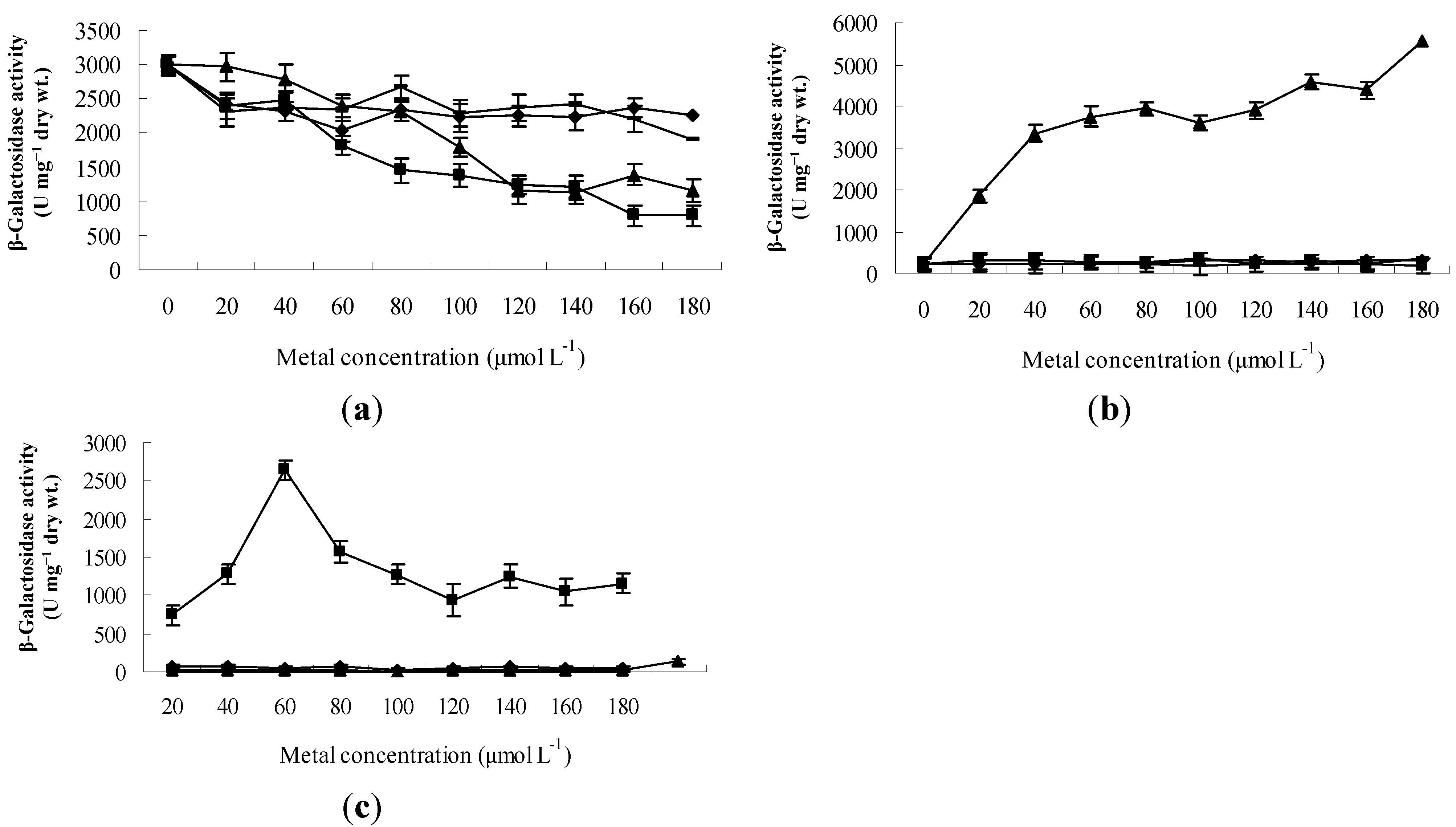
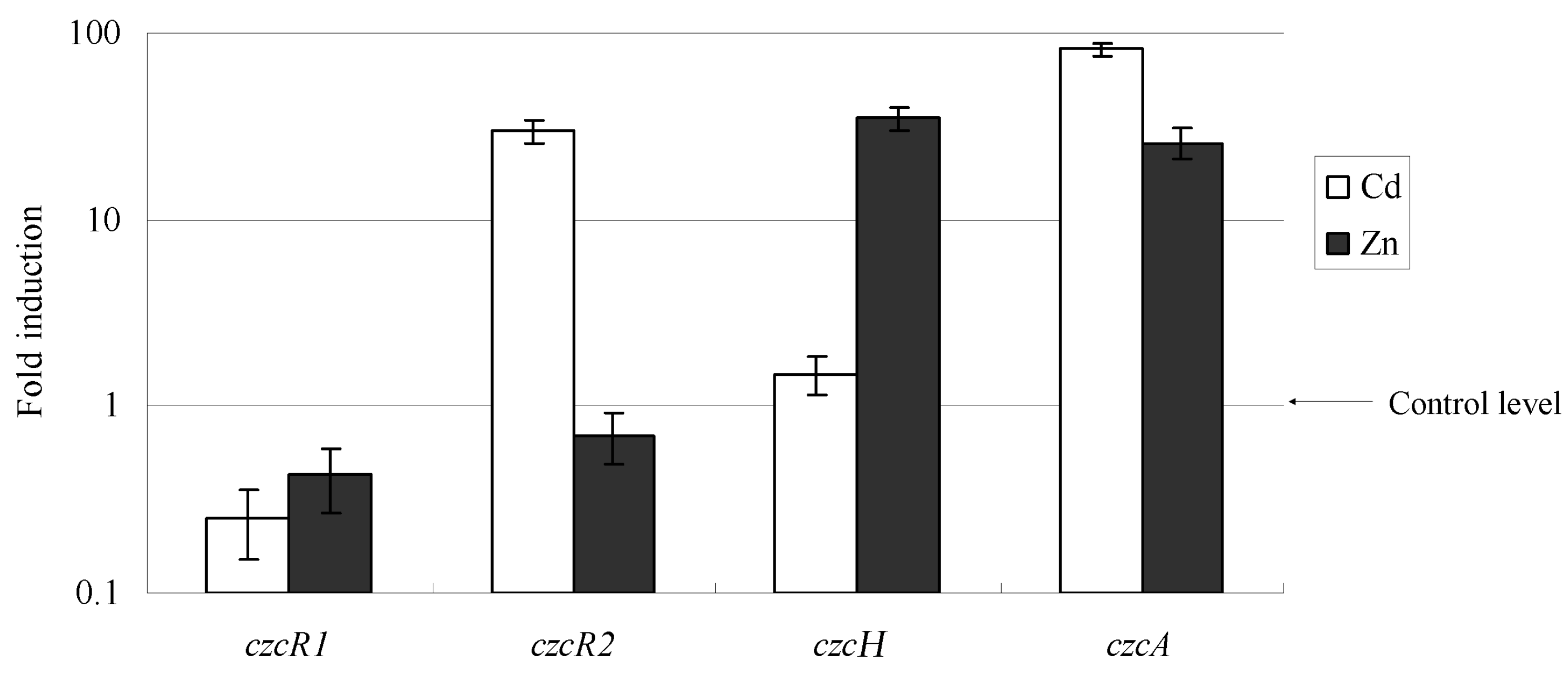
2.3. Cd2+ and Zn2+ Repressed the Transcription of czcRS1, Induced the Transcription of czcRS2 and czcH, Respectively


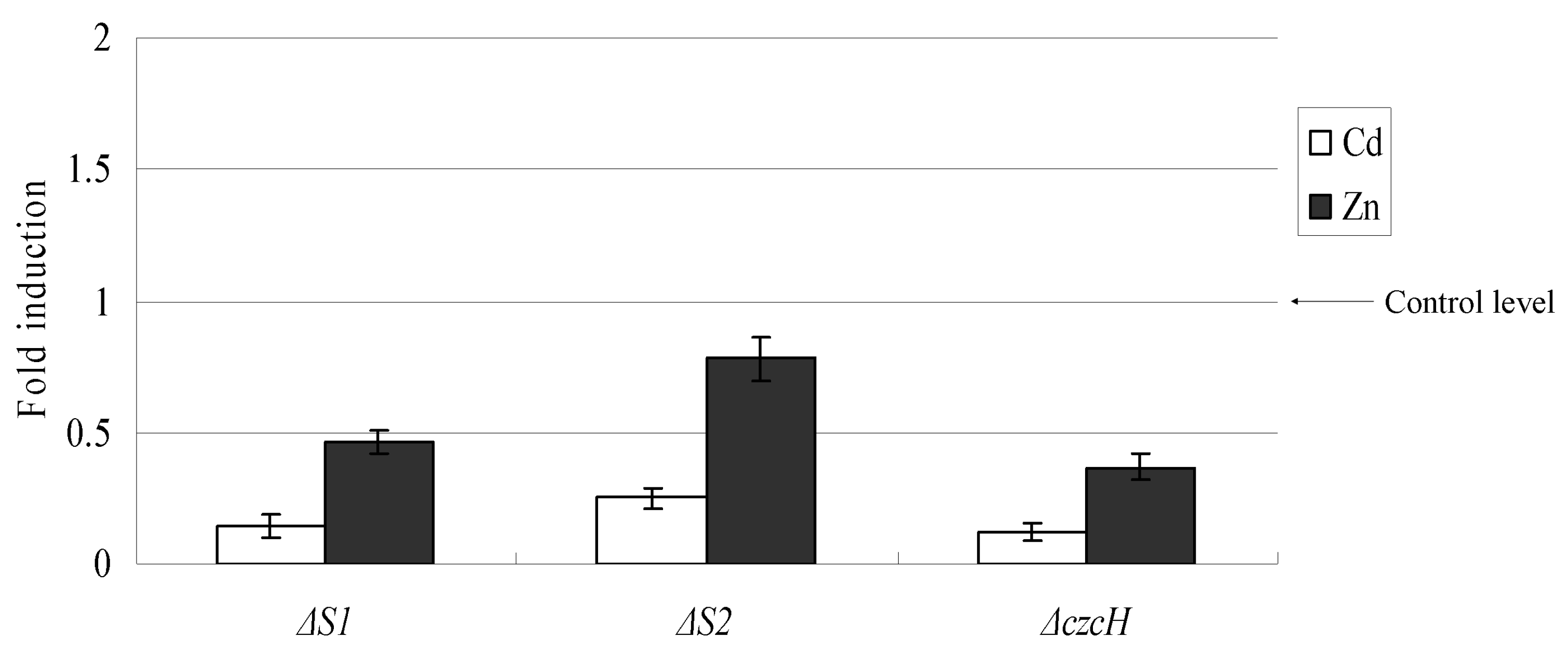
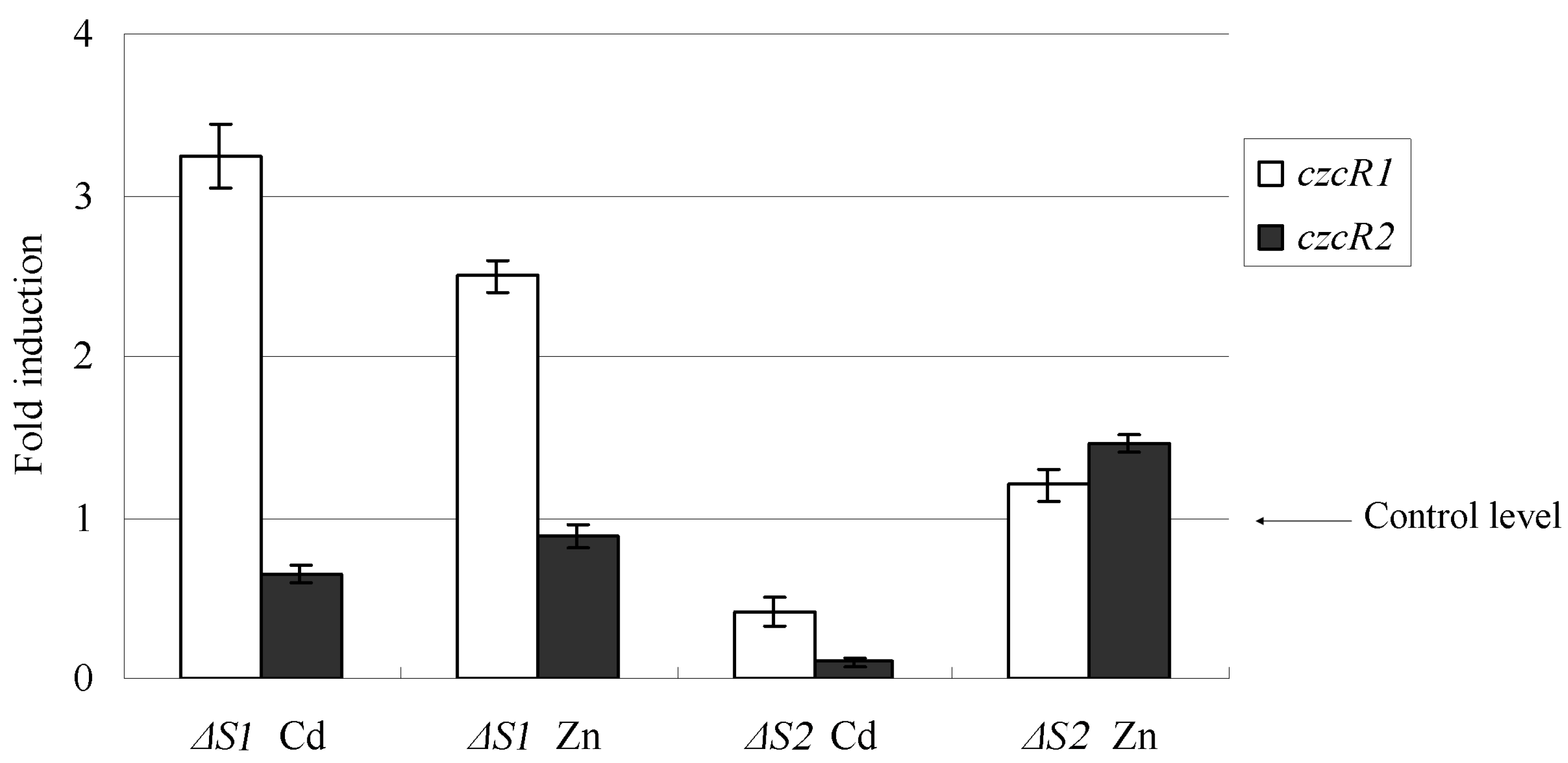
2.4. The Transcription Variation of Mutant Strains


2.5. Deposition of Strains and Nucleotide Sequences
3. Discussion

4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Growth Conditions
| Relevant Characteristics a | Reference | ||
|---|---|---|---|
| Strains | E. coli | ||
| DH5α | supE44 ΔlacU169 (Ф80dlacZΔM15) recA1 endA1 hsdR17 thi-1 gyrA96 relA1 | Invitrogen | |
| C118λpir | Δ(ara-leu) araD ΔlacX74 galE galK phoA20 thi-1 rpsE rpoB argE (Am) recA1 λpir | [23] | |
| MM294 | endA thiA hsdR17 supE44 | [24] | |
| P. putida | |||
| X4 | Apr, Cor, Znr, Cdr, Cur, wild-type | [22] | |
| ΔS1 | X4ΔczcS1 | This study | |
| ΔS1c | X4ΔczcS1, with plasmid pVLTS1 | This study | |
| ΔS2 | X4ΔczcS2 | This study | |
| ΔS2c | X4ΔczcS2, with plasmid pVLTS2 | This study | |
| ΔczcH | X4ΔczcH | This study | |
| ΔczcHc | X4ΔczcH, with plasmid pVLTH | This study | |
| ΔS1S2 | X4ΔczcS1czcS2 | This study | |
| Plasmid | pRK2073 | RK2 helper plasmid, Sper | [24] |
| pTA2 | Cloning vector, Ampr | TOYOBO | |
| pVIK112 | LacZYA, Kmr, suicide vector | [20] | |
| pVIks1 | S1 fragment in pVIK112 | This study | |
| pVIKs2 | S2 fragment in pVIK112 | This study | |
| pVIKH | czcH fragment in pVIK112 | This study | |
| pRRT | Contains a Tetr gene instead of Kmr cassette | This study | |
| pRRTS2 | S2 fragment in pRRT | This study | |
| pVLT31 | lacI, Tetr | [25] | |
| pVLTS1 | pVLT31 with S1 ORF | This study | |
| pVLTS2 | pVLT31 with S2 ORF | This study | |
| pVLTH | pVLT31 with czcH ORF | This study | |
4.2. Determination of MIC Values
4.3. DNA Manipulation
| Purpose | Primer Pair | Sequence (5ʹ-3ʹ) a | Product Length (bp) |
|---|---|---|---|
| For homologous recombination | S1 | gag cag acc tgg aag taa aga | 1113 |
| ggt aga acc gct caa aca a | |||
| S2 | cgt agg cta tgt act tga ggc g | 920 | |
| tgt cgt tga tga tgc ggt tg | |||
| czcH | cac agg gca ttc agg gac caa cgc acg gga taa gag | 496 | |
| gcc cgt tgc acc aca gat | |||
| For qRT-PCR | Qr1 | aca acg gtg tag atg ctc tgc | 121 |
| cgg ctg gtc tta cgg atg g | |||
| Qr2 | gcc gca acg acc agc aac | 144 | |
| gac gca tca gca ggt gta gc | |||
| Qr3 | atg atg ctg acg gcg aga ag | 162 | |
| gcg aat gac ctc tac gga tgc | |||
| QczcA | cca ctg agc acg acc aag g | 128 | |
| aag gtg aag gaa gag gaa ggc | |||
| QrpsL | ctg cgt aaa gta tgc cgt gtg | 174 | |
| gcc cga agt atc cag aga gc | |||
| For complementation experiment | CS1 | cgg ggt acc taa gaa gga gat ata cca tga ggc cat tca gcc tgg | 1455 |
| cta gtc tag att aag cgg cgg tca ttg c | |||
| CS2 | cgg ggt acc taa gaa gga gat ata ccttg aaa aac gcc agc ctg tc | 1419 | |
| cta gtc tag atc act cgg cag gaa aca cca | |||
| CczcH | cgg ggt acc taa gaa gga gat ata cca tga ggt ata gca ttg att atc agc a | 360 | |
| cta gtc tag att ata aga agg cga gcg ag | |||
| For tetracycline resistance gene | Tet | cga cct gca gaa aat agg cgt atc acg agg | 1560 |
| cag cct gca gtc tgc taa cca gta agg caa cga gg |
4.4. Homologous Recombination for Construction of czcS Mutants and lacZ Fusion Reporter Strains
4.5. β-Galactosidase Activity Assay
4.6. Complementation Experiment
4.7. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Blindauer, C.A.; Harrison, M.D.; Parkinson, J.A.; Robinson, A.K.; Turner-Cavfet, J.S.; Robinson, N.J.; Sadler, P.J. A metallothionein containing a zinc finger within a four-metal cluster protects a bacterium from zinc toxicity. Proc. Natl. Acad. Sci. USA 2001, 98, 9593–9598. [Google Scholar] [CrossRef] [PubMed]
- Fantino, J.R.; Py, B.; Fontecave, M.; Barras, F. A genetic analysis of the response of Escherichia coli to cobalt stress. Environ. Microbiol. 2010, 12, 2846–2857. [Google Scholar] [CrossRef] [PubMed]
- Solioz, M.; Stoyanov, J.V. Copper homeostasis in Enterococcus hirae. FEMS Microbiol. Rev. 2003, 27, 183–195. [Google Scholar] [CrossRef]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Cánovas, D.; Cases, I.; de Lorenzo, V. Heavy metal tolerance and metal homeostasis in Pseudomonas putida as revealed by complete genome analysis. Environ. Microbiol. 2003, 5, 1242–1256. [Google Scholar] [CrossRef] [PubMed]
- Leedjarv, A.; Ivask, A.; Virta, M. Interplay of different transporters in the mediation of divalent heavy metal resistance in Pseudomonas putida KT2440. J. Bacteriol. 2008, 190, 2680–2689. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.L.; Stoyanov, J.V.; Kidd, S.P.; Hobman, J.L. The MerR family of transcriptional regulators. FEMS Microbiol. Rev. 2003, 27, 145–163. [Google Scholar] [CrossRef]
- Wu, J.; Rosen, B.P. Metalloregulated expression of the ars operon. J. Biol. Chem. 1993, 268, 52–58. [Google Scholar] [PubMed]
- Caille, O.; Rossier, C.; Perron, K. A copper-activated two-component system interacts with zinc and imipenem resistance in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 4561–4568. [Google Scholar] [CrossRef] [PubMed]
- Große, C.; Grass, G.; Anton, A.; Franke, S.; Santos, A.N.; Lawley, B.; Brown, N.L.; Nies, D.H. Transcriptional organization of the czc heavy-metal homeostasis determinant from Alcaligenes eutrophus. J. Bacteriol. 1999, 181, 2385–2393. [Google Scholar] [PubMed]
- Perron, K.; Caille, O.; Rossier, C.; van Delden, C.; Dumas, J.L.; Köhler, T. CzcR-CzcS, a two-component system involved in heavy metal and carbapenem resistance in Pseudomonas aeruginosa. J. Biol. Chem. 2004, 279, 8761–8768. [Google Scholar] [CrossRef] [PubMed]
- Casino, P.; Rubio, V.; Marina, A. The mechanism of signal transduction by two-component systems. Curr. Opin. Struct. Biol. 2010, 6, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Teitzel, G.M.; Geddie, A.; de Long, S.K.; Kirisits, M.J.; Whiteley, M.; Parsek, M.R. Survival and growth in the presence of elevated copper: Transcriptional profiling of copper-stressed Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 7242–7256. [Google Scholar] [CrossRef] [PubMed]
- Van der Lelie, D.; Schwuchow, T.; Schwidetzky, U.; Wuertz, S.; Baeyens, W.; Mergeay, M.; Nies, D.H. Two-component regulatory system involved in transcriptional control of heavy-metal homoeostasis in Alcaligenes eutrophus. Mol. Microbiol. 1997, 23, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Petit-Haertlen, I.; Girard, E.; Sarret, G.; Hazemann, J.; Gourhant, P.; Kahn, R.; Coves, J. Evidence for conformational changes upon copper binding to Cupriavidus metallidurans CzcE. Biochemistry 2010, 49, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- Grosse, C.; Anton, A.; Hoffmann, T.; Franke, S.; Schleuder, G.; Nies, D.H. Identification of a regulatory pathway that controls the heavy-metal resistance system Czc via promoter czcNp in Ralstonia metallidurans. Arch. Microbiol. 2004, 182, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Timmis, K.N. Pseudomonas putida: A cosmopolitan opportunist par excellence. Environ. Microbiol. 2002, 4, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.C.; Cai, P.; Li, P.X.; Wu, H.Y.; Liang, W.; Rong, X.; Chen, W.; Cai, P. Microcalorimetric and potentiometric titration studies on the adsorption of copper by P. putida and B. thuringiensis and their composites with minerals. J. Hazard. Mater. 2010, 181, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.T.; van der Lelie, D.; Springael, D.; Romling, U.; Ahmed, N.; Mergeay, M. Identification of a gene cluster, czr, involved in cadmium and zinc resistance in Pseudomonas aeruginosa. Gene 1999, 238, 417–425. [Google Scholar] [CrossRef]
- Kalogeraki, V.S.; Winans, S.C. Suicide plasmids containing promoterless reporter genes can simultaneously disrupt and create fusions to target genes of diverse bacteria. Gene 1997, 188, 69–75. [Google Scholar] [CrossRef]
- Stekel, D.J.; Jenkins, D.J. Strong negative self regulation of prokaryotic transcription factors increases the intrinsic noise of protein expression. BMC Syst. Biol. 2008, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.J.; Chen, W.L.; Huang, Q.Y. Construction of two lux-tagged Hg2+-specific biosensors and their luminescence performance. Appl. Microbiol. Biotechnol. 2008, 79, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Keseler, I.M.; Collado-Vides, J.; Gama-Castro, S.; Ingraham, J.; Paley, S.; Paulsen, I.T.; Peralta-Gil, M.; Karp, P.D. A comprehensive database resource for Escherichia coli. Nucleic Acids Res. 2005, 33, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.Y.; Li, Y.G.; Zhou, J.C. Biological characteristics of plasmids of Mesorhizobium huakuii HN3015 from Astragalus sinicus. World J. Microbiol. Biotechnol. 2007, 23, 845–851. [Google Scholar] [CrossRef]
- De Lorenzo, V.; Eltis, L.; Kessler, B.; Timmis, K.N. Analysis of Pseudomonas gene products using lacIq/Ptrp-lac plasmids and transposons that confer conditional phenotypes. Gene 1993, 123, 17–24. [Google Scholar] [CrossRef]
- Mergeay, M.; Nies, D.H.; Schlegel, H.G.; Gerits, J.; Charles, P.; van Gijsegem, F. Alcaligenes eutrophus CH34 is a facultative chemolithotroph with plasmid bound resistance to heavy metals. J. Bacteriol. 1985, 162, 328–334. [Google Scholar] [PubMed]
- Nies, D.H. CzcR and CzcD, gene-products affecting regulation of resistance to cobalt, zinc, and cadmium (czc system) in Alcaligenes eutrophus. J. Bacteriol. 1992, 174, 8102–8110. [Google Scholar] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group wise comparison and stastical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Chen, X.; Huang, Q.; Chen, W. The Role of CzcRS Two-Component Systems in the Heavy Metal Resistance of Pseudomonas putida X4. Int. J. Mol. Sci. 2015, 16, 17005-17017. https://doi.org/10.3390/ijms160817005
Liu P, Chen X, Huang Q, Chen W. The Role of CzcRS Two-Component Systems in the Heavy Metal Resistance of Pseudomonas putida X4. International Journal of Molecular Sciences. 2015; 16(8):17005-17017. https://doi.org/10.3390/ijms160817005
Chicago/Turabian StyleLiu, Pulin, Xi Chen, Qiaoyun Huang, and Wenli Chen. 2015. "The Role of CzcRS Two-Component Systems in the Heavy Metal Resistance of Pseudomonas putida X4" International Journal of Molecular Sciences 16, no. 8: 17005-17017. https://doi.org/10.3390/ijms160817005
APA StyleLiu, P., Chen, X., Huang, Q., & Chen, W. (2015). The Role of CzcRS Two-Component Systems in the Heavy Metal Resistance of Pseudomonas putida X4. International Journal of Molecular Sciences, 16(8), 17005-17017. https://doi.org/10.3390/ijms160817005