
Sulforaphane Prevents Testicular Damage in Kunming Mice Exposed to Cadmium via Activation of Nrf2/ARE Signaling Pathways

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Testis/Body Weight Ratio
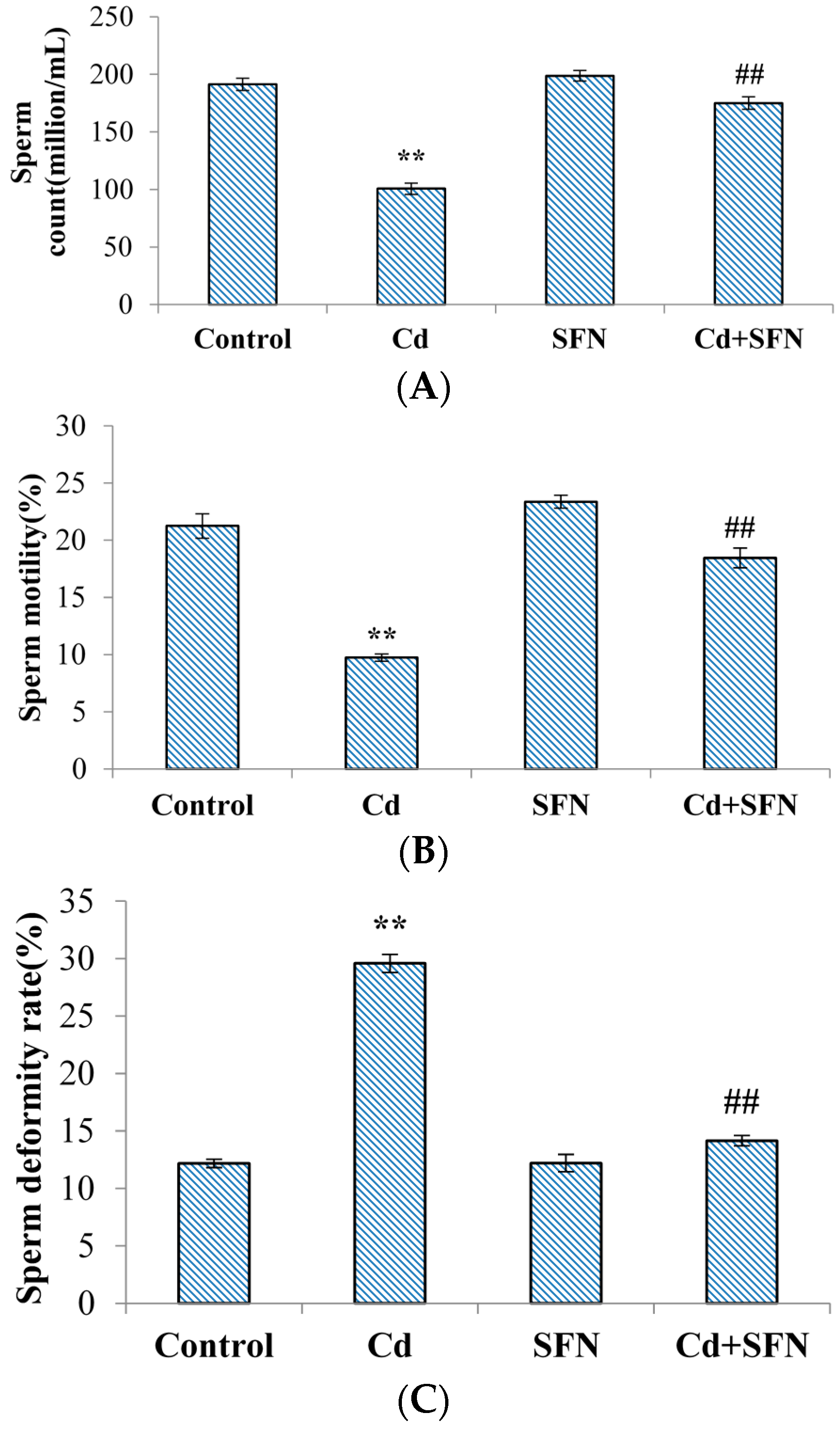
2.2. Spermatological Results
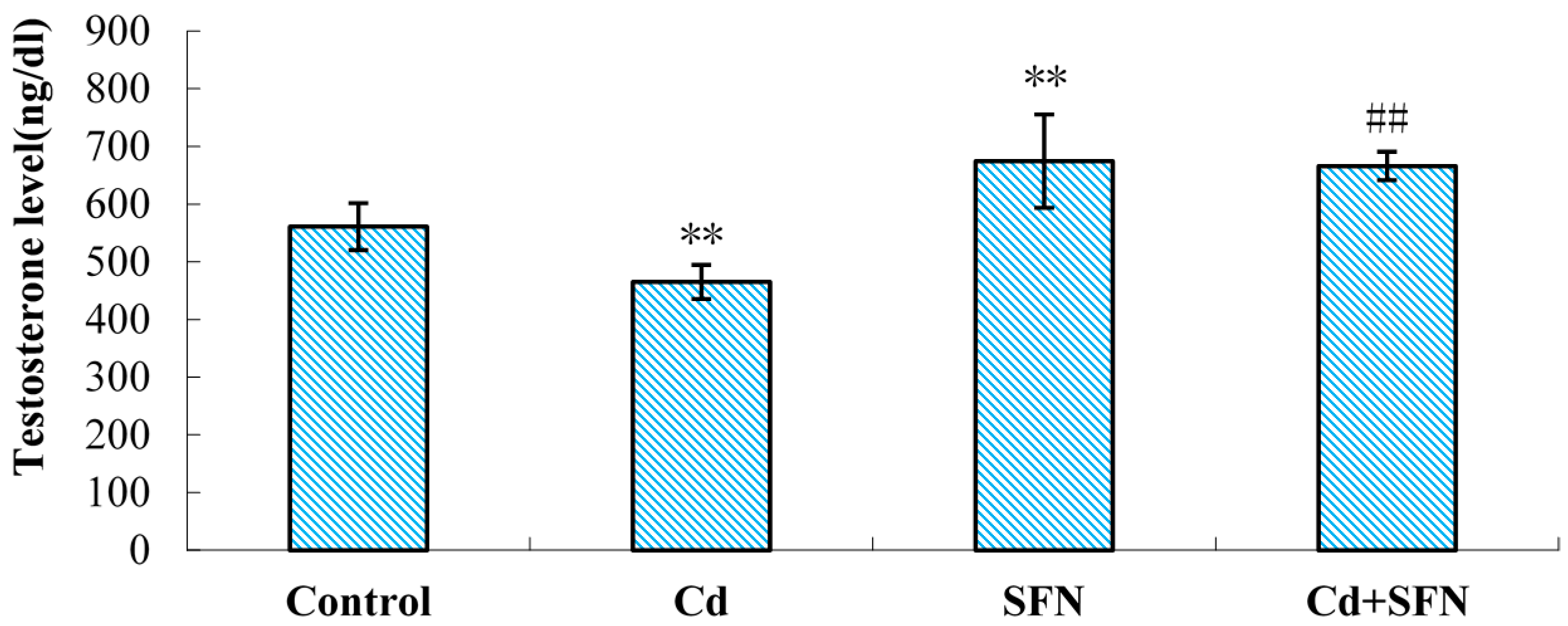
2.3. Serum Testosterone
2.4. Testicular Antioxidant Capacity
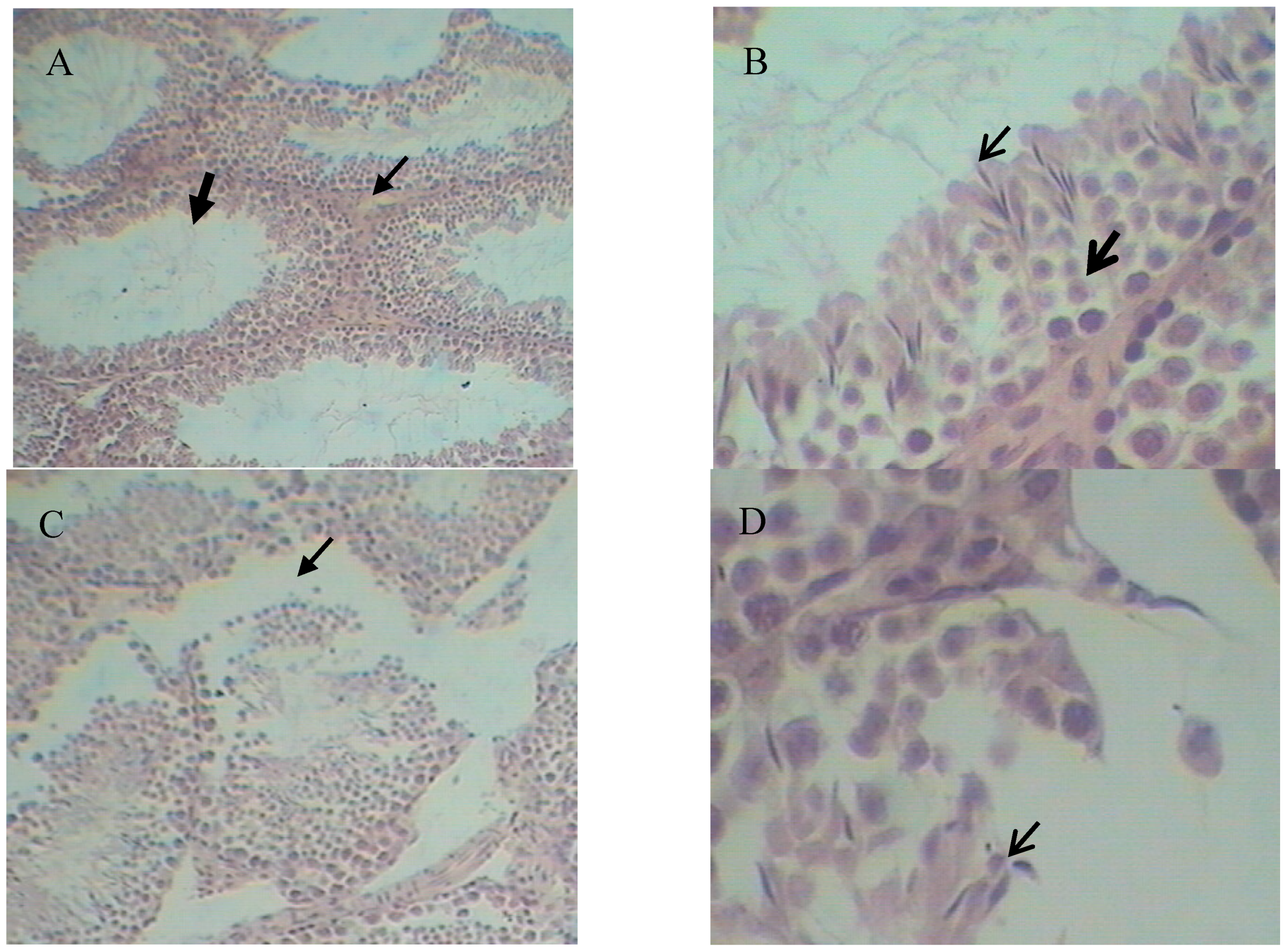
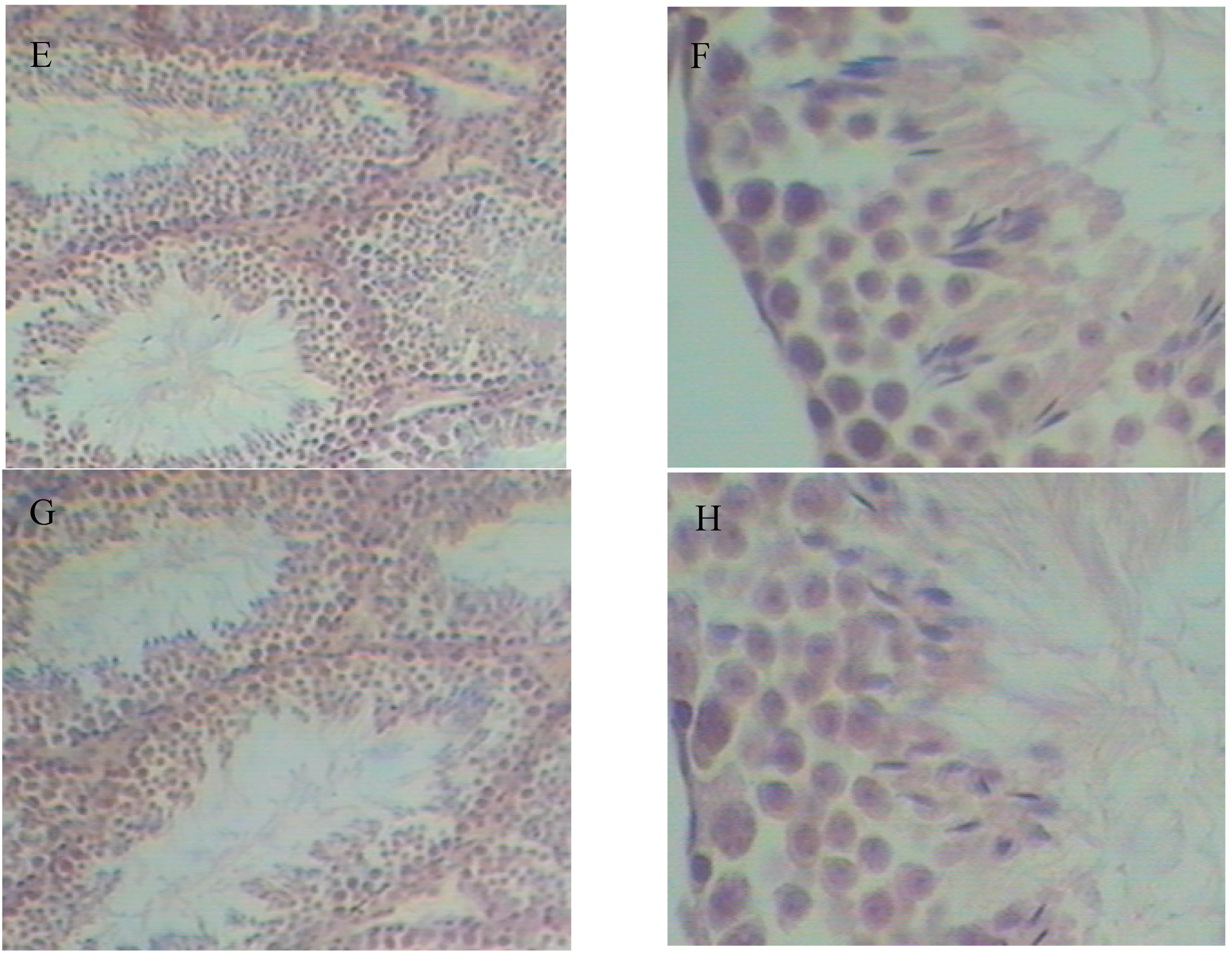
2.5. Histopathological Variation
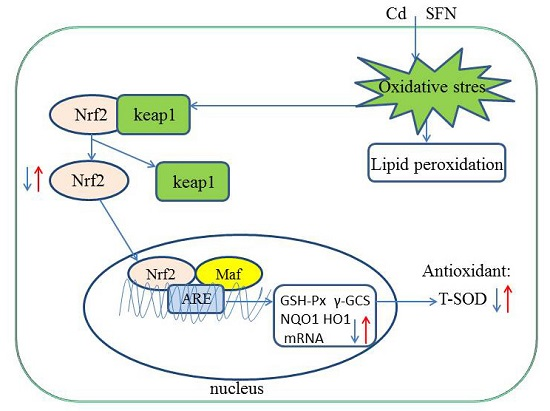
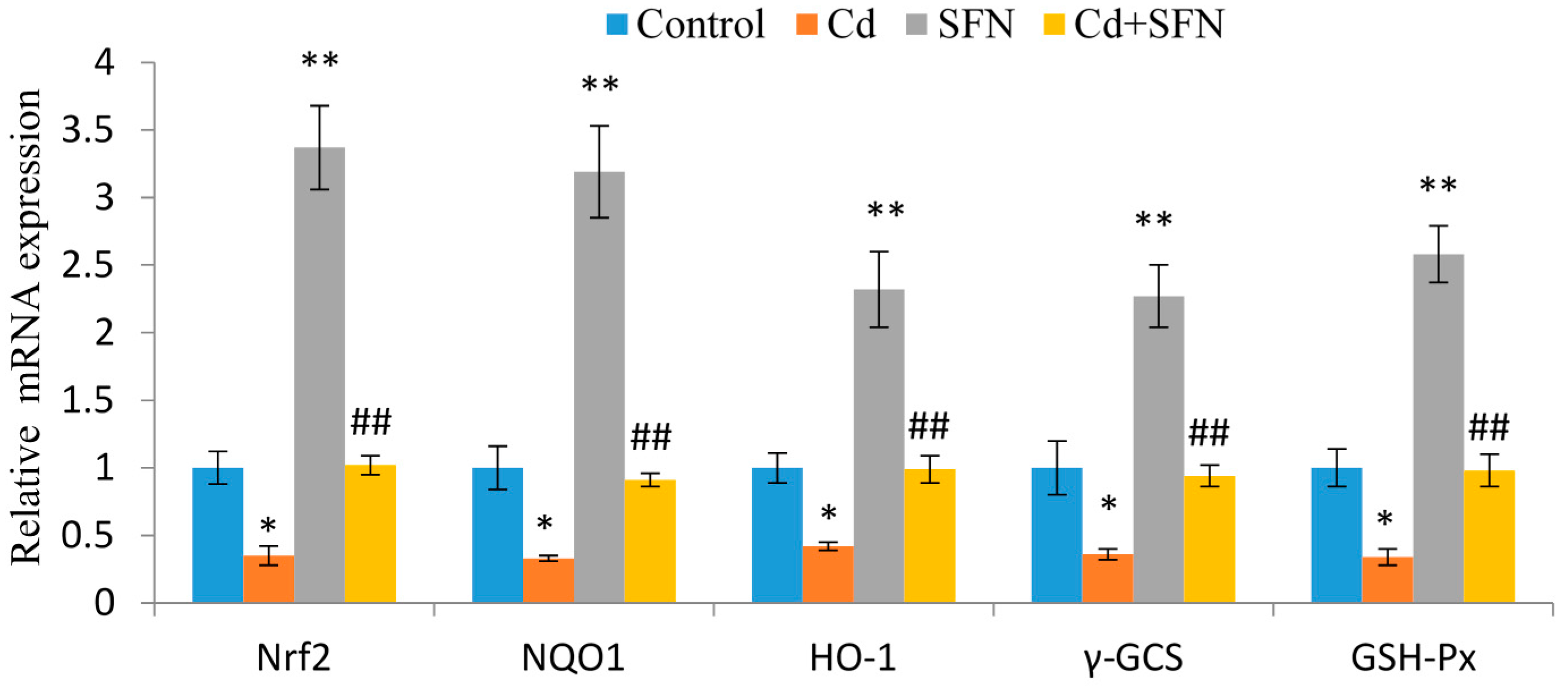
2.6. SFN and Nrf2 Signaling
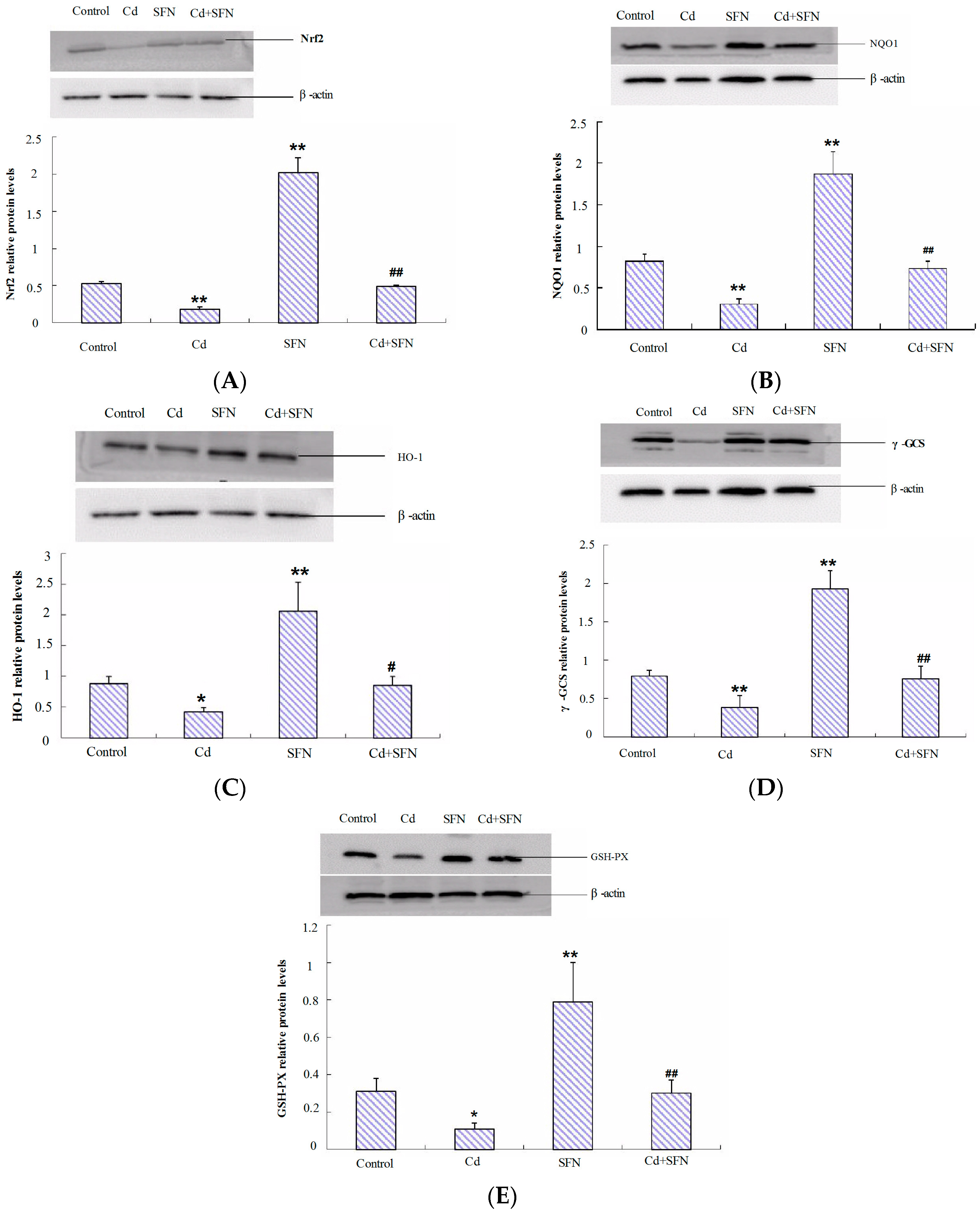
2.7. Nrf2-Related Protein Expression
3. Discussion
4. Experimental Section
4.1. Animals
4.2. Reagents
4.3. Experimental Design and Treatment
4.4. Sperm Parameters
4.5. Determination of Serum Testosterone
4.6. Antioxidant Parameters
4.7. Histopathology
4.8. Gene Expression
4.9. Western Blot
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- IARC. Cadmium and cadmium compounds. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 58, 119–237. [Google Scholar]
- De Souza Predes, F.; Diamante, M.A.; Dolder, H. Testis response to low doses of cadmium in Wistar rats. Int. J. Exp. Pathol. 2010, 91, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Siu, E.R.; Mruk, D.D.; Porto, C.S.; Yan Cheng, C. Cadmium-induced testicular injury. Toxicol. Appl. Pharmacol. 2009, 3, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Alkhedaid, A.; Alshehri, Z.S.; Sabry, A.; Abdel-Ghaffar, T.; Soliman, M.M.; Attia, H. Protective effect of grape seed extract against cadmium-induced testicular dysfunction. Mol. Med. Rep. 2016, 13, 3101–3109. [Google Scholar] [CrossRef] [PubMed]
- Ola-Mudathir, K.F.; Suru, S.M.; Fafunso, M.A.; Obioha, U.E.; Faremi, T.Y. Protective roles of onion and garlic extracts on cadmiuminduced changes in sperm characteristics and testicular oxidative damage in rats. Food Chem. Toxicol. 2008, 46, 3604–3611. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Xia, Y.; Hu, W.; Zhang, H.; Shen, Z. Cadmium induced oxidative damage and protective effects of N-acetyl-l-cysteine against cadmium toxicity in Solanum nigrum L. J. Hazard. Mater. 2010, 180, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.D.; Lin, J.X.; Mi, Y.L.; Zhang, C.Q. Quercetin attenuates cadmium-induced oxidative damage and apoptosis in granulosa cells from chicken ovarian follicles. Reprod. Toxicol. 2011, 31, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Aktas, C.; Kanter, M.; Erboga, M.; Ozturk, S. Anti-apoptotic of curcumin on effects cadmium-induced apoptosis in rat testes. Toxicol. Ind. Health 2012, 28, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Xv, R.Q. Protective effect of grape seed oil on oxidative damage by cadmium in testicular germ cell of rat. Chin. J. Ind. Med. 2013, 26. [Google Scholar]
- Asadi, M.H.; Zafari, F.; Sarveazad, A.; Abbasi, M.; Safa, M.; Koruji, M.; Yari, A.; Alizadeh Miran, R. Saffron improves epididymal sperm parameters in rats exposed to cadmium. Nephrourol. Mon. 2014, 6, e12125. [Google Scholar] [CrossRef] [PubMed]
- Yari, A.; Sarveazad, A.; Asadi, E.; Raouf Sarshoori, J.; Babahajian, A.; Amini, N.; Amidi, F.; Bahadoran, H.; Joghataei, M.; Asadi, M.; et al. Efficacy of Crocus sativus L. on reduction of cadmium-induced toxicity on spermatogenesis in adult rats. Andrologia 2016. [Google Scholar] [CrossRef]
- Lai, R.H.; Keck, A.S.; Walling, M.A.; West, L.G.; Jeffery, E.H. Evaluation of the safety and bioactivity of purified and semi-purified glucoraphanin. Food Chem. Toxicol. 2008, 46, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Zhao, X.J.; Wu, K.; Yuan, L.H. Determination of sulforaphane in vegetables of north diet. Chin. J. Dis. Control Prev. 2005, 9, 209–211. [Google Scholar]
- Fahey, J.W.; Talalay, P. Antioxidant functions of sulforaphane: A potent inducer of Phase II detoxication enzymes. Food Chem. Toxicol. 1999, 37, 973–979. [Google Scholar] [CrossRef]
- Campbell, L.; Howie, F.; Arthur, J.R.; Nicol, F.; Beckett, G. Selenium and sulforaphane modify the expression of selenoenzymes in the human endothelial cell line EAhy926 and protect cells from oxidative damag. Nutrition 2007, 23, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Banning, A. Part of the series: From dietary antioxidants to regulators in cellular signaling and gene regulation. Sulforaphane and selenium, partners in adaptive response and prevention of cancer. Free Radic. Res. 2006, 40, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, S.M.; Royce, S.G.; Licciardi, P.V.; Karagiannis, T.C. Dietary sulforaphane in cancer chemoprevention: The role of epigenetic regulation and HDAC inhibition. Antioxid. Redox Signal. 2015, 22, 1382–1424. [Google Scholar] [CrossRef] [PubMed]
- Fimognari, C.; Hrelia, P. Sulforaphaneas a promising molecule for fighting caneer. Mutat. Res. 2007, 635, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Haristoy, X.; Angioi-Duprez, K.; Duprez, A.; Lozniewski, A. Efficacy of sulforaphane in eradicating Helicobacter pylori in human gastric xenografts implanted in nude mice. Antimicrob. Agents Chemother. 2003, 47, 3982–3984. [Google Scholar] [CrossRef] [PubMed]
- Thejass, P.; Kuttan, G. Augmentation of natural killer cell and antibody-dependent cellular cytotoxicity in BALB/c mice by sulforaphane, a naturally occurring isothiocyanate from broccoli through enhanced production of cytokines IL-2 and IFN-γ. Immunopharmacol. Immunotoxicol. 2006, 28, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane: Translational research from laboratory bench to clinic. Nutr. Rev. 2013, 71, 709–726. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; McMahon, M. NRF2 and KEAP1 mutations: Permanent activation of an adaptive response in cancer. Trends Biochem. Sci. 2009, 34, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, Y.; Piao, Y.; Nagaoka, K.; Watanabe, G.; Taya, K.; Li, C. Protective effects of nuclear factor erythroid 2-related factor 2 on whole body heat stress-induced oxidative damage in the mouse testis. Reprod. Biol. Endocrinol. 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, B.N.; Lawson, G.; Chan, J.Y.; Banuelos, J.; Cortes, M.M.; Hoang, Y.D.; Ortiz, L.; Rau, B.A.; Luderer, U. Knockout of the transcription factor Nrf2 disrupts spermatogenesis in an age-dependent manner. Free Radic. Biol. Med. 2010, 49, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Mai, Z.; Zhou, Y.; Gao, X.; Yu, B. Low Nrf2 mRNA expression in spermatozoa from men with low sperm motility. Tohoku J. Exp. Med. 2012, 228, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Cui, W.; Xin, Y.; Miao, X.; Barati, M.T.; Zhang, C.; Chen, Q.; Tan, Y.; Cui, T.; Zheng, Y.; et al. Prevention by sulforaphane of diabetic cardiomyopathy is associated with up-regulation of Nrf2 expression and transcription activation. J. Mol. Cell. Cardiol. 2013, 57, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Bai, Y.; Miao, X.; Luo, P.; Chen, Q.; Tan, Y.; Rane, M.J.; Miao, L.; Cai, L. Prevention of diabetic nephropathy by sulforaphane: Possible role of Nrf2 upregulation and activation. Oxid. Med. Cell. Longev. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Z.; Guo, W.; Sun, W.; Miao, X.; Wu, H.; Cong, X.; Wintergerst, K.A.; Kong, X.; Cai, L. Sulforaphane reduction of testicular apoptotic cell death in diabetic mice is associated with the up-regulation of Nrf2 expression and function. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E14–E23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Su, Z.Y.; Khor, T.O.; Shu, L.; Kong, A.N. Sulforaphane enhances Nrf2 expression in prostate cancer TRAMP C1 cells through epigenetic regulation. Biochem. Pharmacol. 2013, 85, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.T.; Lysiak, J.J. Oxidative stress: A commonfactor in testicular dysfunction. J. Androl. 2008, 29, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Renugadevi, J.; Prabu, S.M. Naringenin protects against cadmium-induced oxidative renal dysfunction in rats. Toxicology 2002, 56, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Said, L.; Banni, M.; Kerkeni, A.; Said, K.; Messaoudi, I. Influence of combined treatment with zinc and selenium on cadmium induced testicular pathophysiology in rat. Food Chem. Toxicol. 2010, 48, 2759–2765. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, T.; Lei, W.W.; Liu, D.M.; Li, Y.J.; Xuan, R.J.; Ma, J.J. Cadmium-induced oxidative stress and apoptotic changes in the testis of freshwater crab, Sinopotamon henanense. PLoS ONE 2011, 6, e27853. [Google Scholar] [CrossRef] [PubMed]
- Vaca, C.E.; Wilhelm, J.; Hartwig, A.; Harms-Ringgdahl, M. Interaction of lipid peroxidation products with DNA: A review. Mutat. Res. 1988, 195, 137–149. [Google Scholar] [CrossRef]
- Ognjanovic, B.I.; Markovic, S.D.; Ethordevic, N.Z.; Trbojevic, I.S.; Stajn, A.S.; Saicic, Z.S. Cadmium-induced lipid peroxidation and changes in antioxidant defense system in the rat testes: Protective role of coenzyme Q10 and vitamin E. Reprod. Toxicol. 2010, 29, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Luo, X.; Li, L.; Peng, Q.; Yang, Y.; Zhao, L.; Ma, M.; Hou, Z. The protective effects of melatonin against oxidative stress and inflammation induced by acute cadmium exposure in mice testis. Biol. Trace Elem. Res. 2016, 170, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Townsend, D.M.; Tew, K.D.; Tapiero, H. The importance of glutathione in human disease. Biomed. Pharmacother. 2003, 57, 145–155. [Google Scholar] [CrossRef]
- Uruno, A.; Motohashi, H. The Keap1-Nrf2 system as an in vivo sensor for electrophiles. Nitric Oxide 2011, 25, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.F.; Tan, C.B.; Shi, L.L.; Zhang, J.; Xu, W.R.; Tang, L.D. Advances in study on Nrf2/ARE signal pathway and its relevant drugs. Drug Eval. Res. 2013, 36, 54–59. [Google Scholar]
- Kwak, M.K.; Itoh, K.; Yamamoto, M.; Kensler, T.W. Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: Role of antioxidant response element-like sequences in the Nrf2 promoter. Mol. Cell. Biol. 2002, 22, 2883–2892. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, N.; Daggett, M.A.; Chu, W.A.; Bittel, D.; Johnson, J.A.; Andrews, G.K. Participation of upstream stimulator factor (USF) in cadmium-induction of the mouse metallothionein-I gene. Nucleic Acids Res. 1998, 26, 5182–5189. [Google Scholar] [CrossRef] [PubMed]
- Nazimabashir; Manoharan, V.; Miltonprabu, S. Cadmium induced cardiac oxidative stress in rats and its attenuation by GSP through the activation of Nrf2 signaling pathway. Chem. Biol. Interact. 2015, 242, 179–193. [Google Scholar]
- Ciftci, O.; Aydin, M.; Ozdemir, I.; Vardi, N. Quercetin prevents 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced testicular damage in rats. Andrologia 2012, 44, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, O.; Ozdemir, I.; Aydin, M.; Beytur, A. Beneficial effects of chrysin on the reproductive system of adult male rats. Andrologia 2012, 44, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Yang, S.H.; Han, J.X.; Li, P.; Zhang, Y.; Dong, S.; Chen, X.; Guo, J.; Wang, J.; He, J.B. The protective effect of grape-seed proanthocyanidin extract on oxidative damage induced by zearalenone in Kunming mice liver. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (g) | Testis Weight (mg) | Testis/Body Weight Ratio (mg/g) |
|---|---|---|---|
| Control | 45.57 ± 1.69 | 304.47 ± 10.04 | 6.68 ± 0.10 |
| CdCl2 (2.3 mg/kg) | 42.75 ± 1.42 | 193.65 ± 20.30 ** | 4.60 ± 0.64 ** |
| SFN (10 mg/kg) | 40.43 ± 0.89 * | 278.27 ± 11.84 | 6.88 ± 0.33 |
| CdCl2 (2.3 mg/kg) + SFN (10 mg/kg) | 40.35 ± 1.89 | 223.12 ± 28.75 | 5.48 ± 0.53 |
| Group | T-SOD (U/mgprot) | GSH (mg/gprot) | MDA (nmol/mgprot) |
|---|---|---|---|
| Control | 86.67 ± 3.43 | 142.47 ± 2.93 | 12.69 ± 1.07 |
| CdCl2 (2.3 mg/kg) | 43.23 ± 12.76 ** | 33.63 ± 10.15 ** | 65.83 ± 1.07 ** |
| SFN (10 mg/kg) | 119.92 ± 6.41 * | 162.14 ± 7.68 * | 11.21 ± 0.69 * |
| CdCl2 (2.3 mg/kg)+ SFN (10 mg/kg) | 104.61 ± 6.10 ## | 118.09 ± 10.15 ## | 19.17 ± 4.17 ## |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-H.; Long, M.; Yu, L.-H.; Li, L.; Li, P.; Zhang, Y.; Guo, Y.; Gao, F.; Liu, M.-D.; He, J.-B. Sulforaphane Prevents Testicular Damage in Kunming Mice Exposed to Cadmium via Activation of Nrf2/ARE Signaling Pathways. Int. J. Mol. Sci. 2016, 17, 1703. https://doi.org/10.3390/ijms17101703
Yang S-H, Long M, Yu L-H, Li L, Li P, Zhang Y, Guo Y, Gao F, Liu M-D, He J-B. Sulforaphane Prevents Testicular Damage in Kunming Mice Exposed to Cadmium via Activation of Nrf2/ARE Signaling Pathways. International Journal of Molecular Sciences. 2016; 17(10):1703. https://doi.org/10.3390/ijms17101703
Chicago/Turabian StyleYang, Shu-Hua, Miao Long, Li-Hui Yu, Lin Li, Peng Li, Yi Zhang, Yang Guo, Feng Gao, Ming-Da Liu, and Jian-Bin He. 2016. "Sulforaphane Prevents Testicular Damage in Kunming Mice Exposed to Cadmium via Activation of Nrf2/ARE Signaling Pathways" International Journal of Molecular Sciences 17, no. 10: 1703. https://doi.org/10.3390/ijms17101703