
Translationally Controlled Tumor Protein Stimulates Dopamine Release from PC12 Cells via Ca2+-Independent Phospholipase A2 Pathways


Abstract
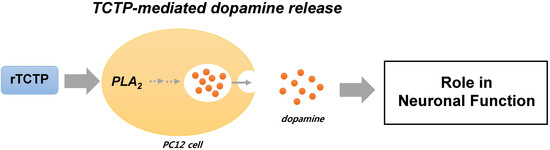
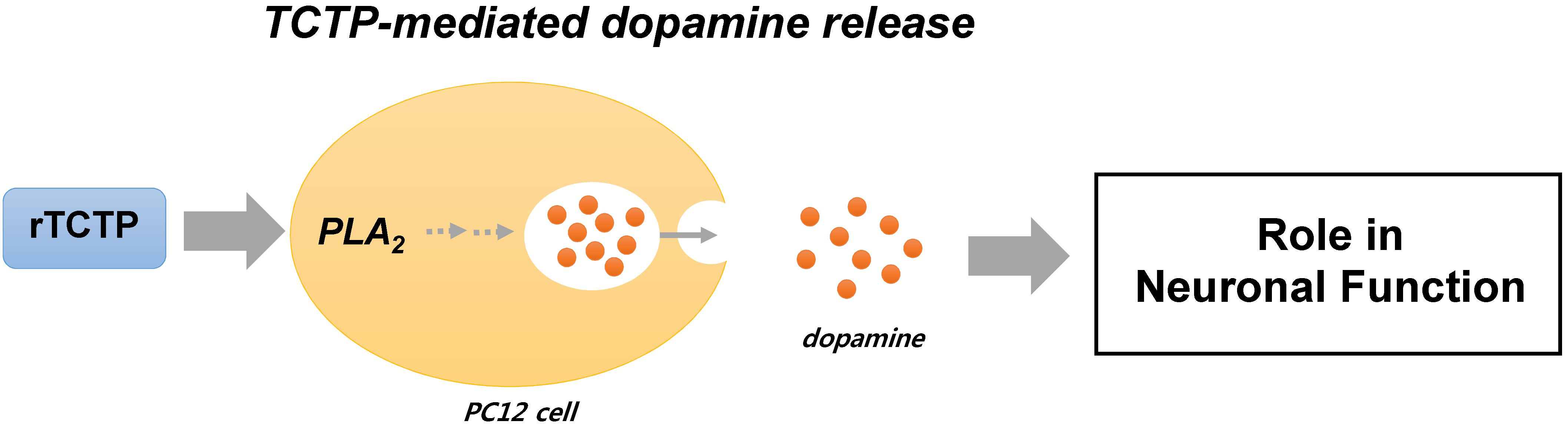
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Expression of Endogenous Translationally Controlled Tumor Protein (TCTP) in PC12 Cells and Rat Brain Tissues
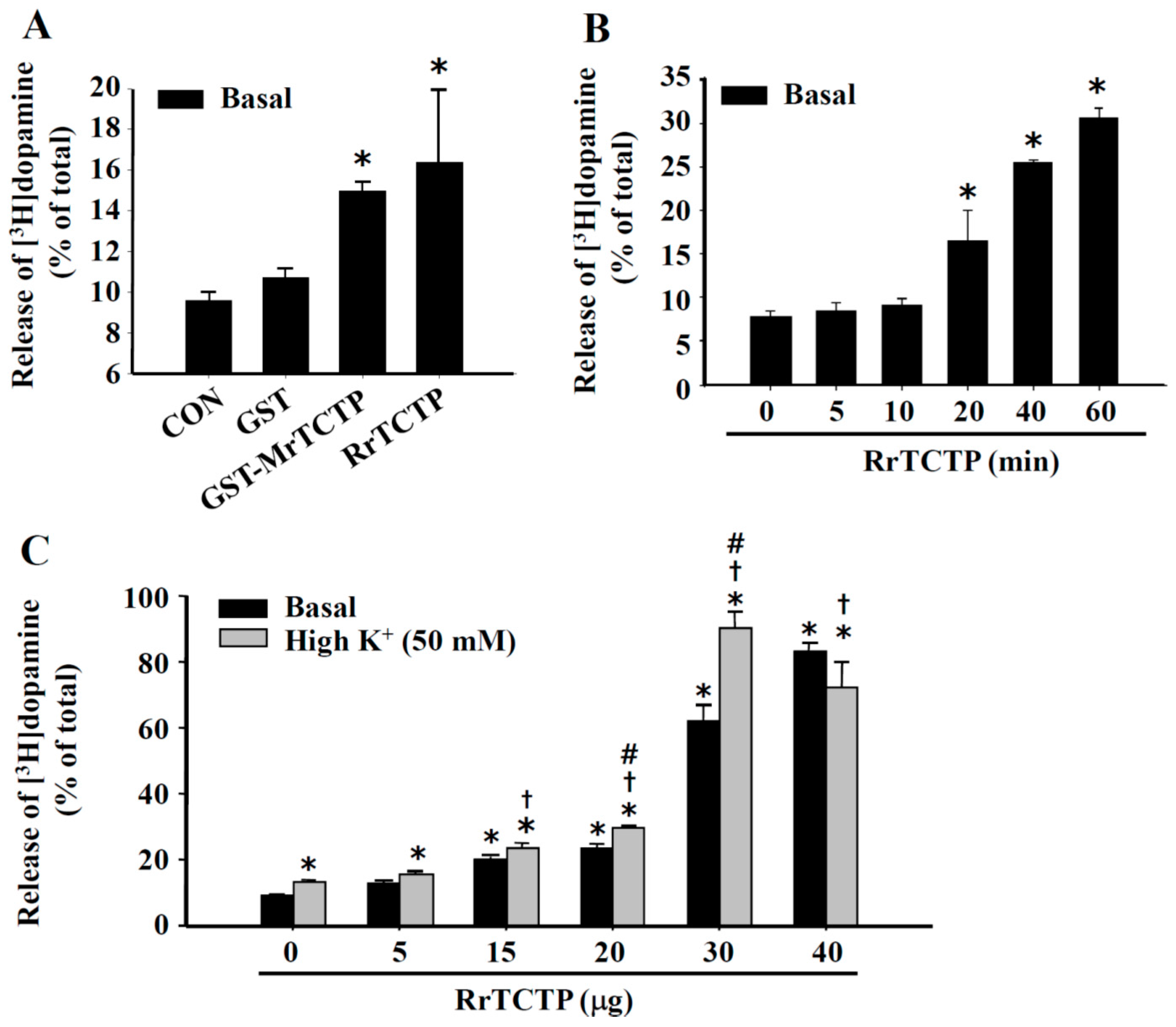
2.2. Stimulatory Effect of TCTP on Basal and Depolarization-Induced Release of Dopamine from PC12 Cells
2.3. Regulation of Intracellular Ca2+ Levels by TCTP in PC12 Cells
2.4. Ca2+-Independent Release of Dopamine by TCTP from PC12 Cells
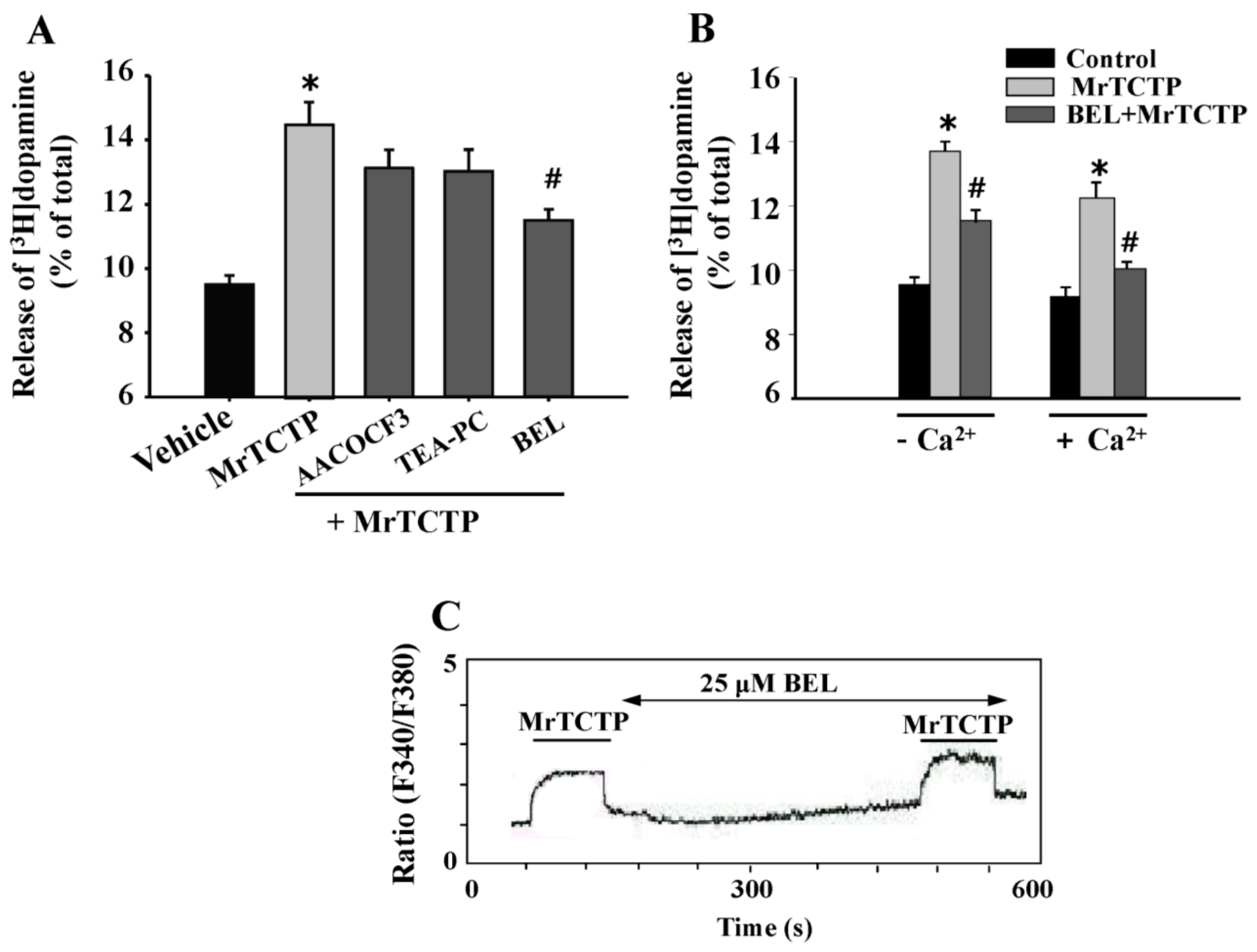
2.5. Involvement of Ca2+-Independent Phospholipase A2 (PLA2) in TCTP-Induced Dopamine Release from PC12 Cells
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Expression and Purification of TCTP
4.3. Neuronal Cell Culture
4.4. SDS-Polyacrylamide Gel Electrophoresis (PAGE) and Western Blotting
4.5. [3H]Dopamine Release
4.6. Measurement of Changes in [Ca2+]i
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bommer, U.A.; Thiele, B.J. The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol. 2004, 36, 379–385. [Google Scholar] [CrossRef]
- Bohm, H.; Benndorf, R.; Gaestel, M.; Gross, B.; Nurnberg, P.; Kraft, R.; Otto, A.; Bielka, H. The growth-related protein P23 of the Ehrlich ascites tumor: Translational control, cloning and primary structure. Biochem. Int. 1989, 19, 277–286. [Google Scholar] [PubMed]
- Yenofsky, R.; Bergmann, I.; Brawerman, G. Messenger RNA species partially in a repressed state in mouse sarcoma ascites cells. Proc. Natl. Acad. Sci. USA 1982, 79, 5876–5880. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, S.M.; Rafnar, T.; Langdon, J.; Lichtenstein, L.M. Molecular identification of an IgE-dependent histamine-releasing factor. Science 1995, 269, 688–690. [Google Scholar] [CrossRef] [PubMed]
- Nagano-Ito, M.; Ichikawa, S. Biological effects of Mammalian translationally controlled tumor protein (TCTP) on cell death, proliferation, and tumorigenesis. Biochem. Res. Int. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Thiele, H.; Berger, M.; Skalweit, A.; Thiele, B.J. Expression of the gene and processed pseudogenes encoding the human and rabbit translationally controlled tumour protein (TCTP). Eur. J. Biochem. 2000, 267, 5473–5481. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Cairns, N.; Fountoulakisc, M.; Lubec, G. Decreased brain histamine-releasing factor protein in patients with Down syndrome and Alzheimer’s disease. Neurosci. Lett. 2001, 300, 41–44. [Google Scholar] [CrossRef]
- Chung, C.; Tallerico, T.; Seeman, P. Schizophrenia hippocampus has elevated expression of chondrex glycoprotein gene. Synapse 2003, 50, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Andreassi, C.; Zimmermann, C.; Mitter, R.; Fusco, S.; de Vita, S.; Saiardi, A.; Riccio, A. An NGF-responsive element targets myo-inositol monophosphatase-1 mRNA to sympathetic neuron axons. Nat. Neurosci. 2010, 13, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Gumy, L.F.; Yeo, G.S.; Tung, Y.C.; Zivraj, K.H.; Willis, D.; Coppola, G.; Lam, B.Y.; Twiss, J.L.; Holt, C.E.; Fawcett, J.W. Transcriptome analysis of embryonic and adult sensory axons reveals changes in mRNA repertoire localization. RNA 2011, 17, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Roque, C.G.; Wong, H.H.; Lin, J.Q.; Holt, C.E. Tumor protein TCTP regulates axon development in the embryonic visual system. Development 2016, 143, 1134–1148. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Berchtold, N.C.; Perreau, V.M.; Tu, C.H.; Li Jeon, N.; Cotman, C.W. Axonal mRNA in uninjured and regenerating cortical mammalian axons. J. Neurosci. 2009, 29, 4697–4707. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Milani, D.; Leon, A.; Meldolesi, J.; Pozzan, T. Voltage-dependent activation and inactivation of calcium channels in PC12 cells. Correlation with neurotransmitter release. J. Biol. Chem. 1987, 262, 9189–9195. [Google Scholar] [PubMed]
- Greene, L.A.; Rein, G. Release of [3H]norepinephrine from a clonal line of pheochromocytoma cells (PC12) by nicotinic cholinergic stimulation. Brain Res. 1977, 138, 521–528. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, S.; Jung, J.E.; Roh, E.J.; Kim, H.J. Capacitative Ca2+ entry is involved in regulating soluble amyloid precursor protein (sAPPα) release mediated by muscarinic acetylcholine receptor activation in neuroblastoma SH-SY5Y cells. J. Neurochem. 2006, 97, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Thastrup, O.; Cullen, P.J.; Drobak, B.K.; Hanley, M.R.; Dawson, A.P. Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2+-ATPase. Proc. Natl. Acad. Sci. USA 1990, 87, 2466–2470. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raya, S.; Bloch-Shilderman, E.; Lelkes, P.I.; Trembovler, V.; Shohami, E.; Gutman, Y.; Lazarovici, P. Characterization of pardaxin-induced dopamine release from pheochromocytoma cells: Role of calcium and eicosanoids. J. Pharmacol. Exp. Ther. 1999, 288, 399–406. [Google Scholar] [PubMed]
- Lazarovici, P.; Lelkes, P.I. Pardaxin induces exocytosis in bovine adrenal medullary chromaffin cells independent of calcium. J. Pharmacol. Exp. Ther. 1992, 263, 1317–1326. [Google Scholar] [PubMed]
- Goetzl, E.J.; An, S.; Smith, W.L. Specificity of expression and effects of eicosanoid mediators in normal physiology and human diseases. FASEB J. 1995, 9, 1051–1058. [Google Scholar] [PubMed]
- Hermann, P.M.; Watson, S.N.; Wildering, W.C. Phospholipase A2—Nexus of aging, oxidative stress, neuronal excitability, and functional decline of the aging nervous system? Insights from a snail model system of neuronal aging and age-associated memory impairment. Front. Genet. 2014, 5, 419. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Balboa, M.A.; Insel, P.A.; Dennis, E.A. Regulation and inhibition of phospholipase A2. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Ramanadham, S.; Ali, T.; Ashley, J.W.; Bone, R.N.; Hancock, W.D.; Lei, X. Calcium-independent phospholipases A2 and their roles in biological processes and diseases. J. Lipid Res. 2015, 56, 1643–1668. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Singaravelu, K.; Lohr, C.; Deitmer, J.W. Regulation of store-operated calcium entry by calcium-independent phospholipase A2 in rat cerebellar astrocytes. J. Neurosci. 2006, 26, 9579–9592. [Google Scholar] [CrossRef] [PubMed]
- Vaudry, D.; Stork, P.J.; Lazarovici, P.; Eiden, L.E. Signaling pathways for PC12 cell differentiation: Making the right connections. Science 2002, 296, 1648–1649. [Google Scholar] [CrossRef] [PubMed]
- Yermolaieva, O.; Brot, N.; Weissbach, H.; Heinemann, S.H.; Hoshi, T. Reactive oxygen species and nitric oxide mediate plasticity of neuronal calcium signaling. Proc. Natl. Acad. Sci. USA 2000, 97, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Fasolato, C.; Pandiella, A.; Meldolesi, J.; Pozzan, T. Generation of inositol phosphates, cytosolic Ca2+, and ionic fluxes in PC12 cells treated with bradykinin. J. Biol. Chem. 1988, 263, 17350–17359. [Google Scholar] [PubMed]
- Weiss, C.; Atlas, D. The bradykinin receptor—A putative receptor-operated channel in PC12 cells: Studies of neurotransmitter release and inositol phosphate accumulation. Brain Res. 1991, 543, 102–110. [Google Scholar] [CrossRef]
- Adam-Vizi, V.; Ligeti, E. Release of acetylcholine from rat brain synaptosomes by various agents in the absence of external calcium ions. J. Physiol. 1984, 353, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.A. Depolarization without calcium can release γ-aminobutyric acid from a retinal neuron. Science 1987, 238, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.G.; Sihra, T.S.; Sanchez-Prieto, J. Calcium-dependent and -independent release of glutamate from synaptosomes monitored by continuous fluorometry. J. Neurochem. 1987, 49, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Lonart, G.; Zigmond, M.J. High glutamate concentrations evoke Ca(++)-independent dopamine release from striatal slices: A possible role of reverse dopamine transport. J. Pharmacol. Exp. Ther. 1991, 256, 1132–1138. [Google Scholar] [PubMed]
- Hochner, B.; Parnas, H.; Parnas, I. Membrane depolarization evokes neurotransmitter release in the absence of calcium entry. Nature 1989, 342, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Bloch-Shilderman, E.; Abu-Raya, S.; Trembovler, V.; Boschwitz, H.; Gruzman, A.; Linial, M.; Lazarovici, P. Pardaxin stimulation of phospholipases A2 and their involvement in exocytosis in PC-12 cells. J. Pharmacol. Exp. Ther. 2002, 301, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Bonventre, J.V. Roles of phospholipases A2 in brain cell and tissue injury associated with ischemia and excitotoxicity. J. Lipid Mediat. Cell Signal. 1997, 16, 199–208. [Google Scholar] [CrossRef]
- Matsuzawa, A.; Murakami, M.; Atsumi, G.; Imai, K.; Prados, P.; Inoue, K.; Kudo, I. Release of secretory phospholipase A2 from rat neuronal cells and its possible function in the regulation of catecholamine secretion. Biochem. J. 1996, 318, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Berman, J.D.; Middleton, W.; Brendle, J. Botulinum toxin inhibits arachidonic acid release associated with acetylcholine release from PC12 cells. J. Biol. Chem. 1993, 268, 11057–11064. [Google Scholar] [PubMed]
- Song, H.S.; Ko, M.S.; Jo, Y.S.; Whang, W.K.; Sim, S.S. Inhibitory effect of acteoside on melittin-induced catecholamine exocytosis through inhibition of Ca2+-dependent phospholipase A2 and extracellular Ca2+ influx in PC12 cells. Arch. Pharm. Res. 2015, 38, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Rathinavelu, A.; Borowitz, J.L.; Isom, G.E. Activation of a calcium- and pH-dependent phospholipase A2 by cyanide in PC12 cells. Toxicol. Appl. Pharmacol. 1994, 124, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Ong, W.Y.; Horrocks, L.A. Inhibitors of brain phospholipase A2 activity: Their neuropharmacological effects and therapeutic importance for the treatment of neurologic disorders. Pharmacol. Rev. 2006, 58, 591–620. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.C.; Mosior, M.; Ni, B.; Dennis, E.A. Regional distribution, ontogeny, purification, and characterization of the Ca2+-independent phospholipase A2 from rat brain. J. Neurochem. 1999, 73, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Smani, T.; Dominguez-Rodriguez, A.; Callejo-Garcia, P.; Rosado, J.A.; Avila-Medina, J. Phospholipase A2 as a Molecular Determinant of Store-Operated Calcium Entry. Adv. Exp. Med. Biol. 2016, 898, 111–131. [Google Scholar] [PubMed]
- Vonakis, B.M.; Macglashan, D.W., Jr.; Vilarino, N.; Langdon, J.M.; Scott, R.S.; MacDonald, S.M. Distinct characteristics of signal transduction events by histamine-releasing factor/translationally controlled tumor protein (HRF/TCTP)-induced priming and activation of human basophils. Blood 2008, 111, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.T.; Lichtenstein, L.M.; MacDonald, S.M. An immunoglobulin E-dependent recombinant histamine-releasing factor induces interleukin-4 secretion from human basophils. J. Exp. Med. 1996, 183, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- MacGlashan, D., Jr.; Botana, L.M. Biphasic Ca2+ responses in human basophils. Evidence that the initial transient elevation associated with the mobilization of intracellular calcium is an insufficient signal for degranulation. J. Immunol. 1993, 150, 980–991. [Google Scholar] [PubMed]
- Balboa, M.A.; Balsinde, J.; Dennis, E.A. Involvement of phosphatidate phosphohydrolase in arachidonic acid mobilization in human amnionic WISH cells. J. Biol. Chem. 1998, 273, 7684–7690. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, J.; Maeng, J.; Kim, H.-J. Translationally Controlled Tumor Protein Stimulates Dopamine Release from PC12 Cells via Ca2+-Independent Phospholipase A2 Pathways. Int. J. Mol. Sci. 2016, 17, 1774. https://doi.org/10.3390/ijms17101774
Seo J, Maeng J, Kim H-J. Translationally Controlled Tumor Protein Stimulates Dopamine Release from PC12 Cells via Ca2+-Independent Phospholipase A2 Pathways. International Journal of Molecular Sciences. 2016; 17(10):1774. https://doi.org/10.3390/ijms17101774
Chicago/Turabian StyleSeo, Jihui, Jeehye Maeng, and Hwa-Jung Kim. 2016. "Translationally Controlled Tumor Protein Stimulates Dopamine Release from PC12 Cells via Ca2+-Independent Phospholipase A2 Pathways" International Journal of Molecular Sciences 17, no. 10: 1774. https://doi.org/10.3390/ijms17101774
APA StyleSeo, J., Maeng, J., & Kim, H.-J. (2016). Translationally Controlled Tumor Protein Stimulates Dopamine Release from PC12 Cells via Ca2+-Independent Phospholipase A2 Pathways. International Journal of Molecular Sciences, 17(10), 1774. https://doi.org/10.3390/ijms17101774