
Neuroprotection via Reduction in Stress: Altered Menstrual Patterns as a Marker for Stress and Implications for Long-Term Neurologic Health in Women

Abstract
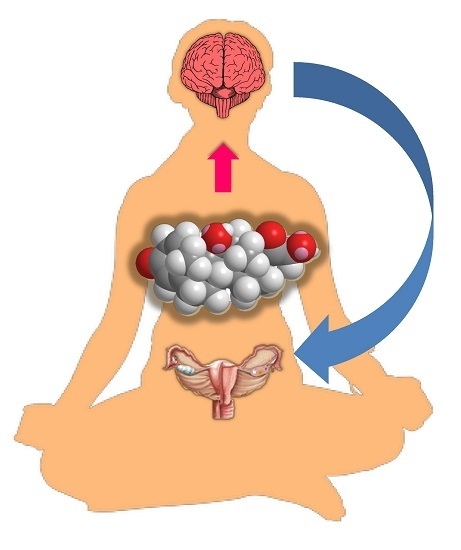
:
1. Introduction
2. Role of Stress in Functional Hypothalamic Amenorrhea (FHA)
3. Glucocorticoid Effects upon Reproductive Tissues
4. Neuroprotection via Reduction in Stress
5. Molecular Consequences of Stress
6. Stress and Neurodegenerative Diseases
7. Conclusions
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| CBT | Cognitive behavioral therapy |
| CNS | Central nervous system |
| CRH | Corticotrophin-releasing hormone |
| CSF | Cerebrospinal fluid |
| FHA | Functional hypothalamic amenorrhea |
| GnRH | Gonadotropin-releasing hormone |
| HPA | Hypothalamus–pituitary–adrenal |
| TSH | Thyroid-stimulating hormone |
References
- Schneiderman, N.; Ironson, G.S.; Siegel, S.D. Stress and health: Psychological, behavioral, and biological determinants. Annu. Rev. Clin. Psychol. 2005, 1, 607–628. [Google Scholar] [CrossRef] [PubMed]
- Berga, S.L. Functional hypothalamic chronic anovulation. In Reproductive Endocrinology, Surgery, and Technology; Adashi, E.Y., Rock, J.A., Rosenwaks, Z., Eds.; Lippincott–Raven: Philadelphia, PA, USA, 1996; pp. 1061–1075. [Google Scholar]
- Berga, S.L. Stress and reprodution: A tale of false dichotomy? Endocrinology 2008, 149, 867–868. [Google Scholar] [CrossRef] [PubMed]
- Reame, N.E.; Sauder, S.E.; Case, G.D.; Kelch, R.P.; Marshall, J.C. Pulsatile gonadotropin secretion in women with hypothalamic amenorrhea: Evidence that reduced frequency of gonadotropin-releasing hormone secretion is the mechanism of persistent anovulation. J. Clin. Endocrinol. Metab. 1985, 61, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Crowley, W.F., Jr.; Filicori, M.; Spratt, D.I.; Santoro, N. The physiology of gonadotropin-releasing hormone (GnRH) secretion in men and women. Recent Prog. Horm. Res. 1985, 41, 473–526. [Google Scholar] [PubMed]
- Berga, S.L.; Mortola, J.F.; Girton, L.; Suh, B.; Laughlin, G.; Pham, P.; Yen, S.S.C. Neuroendocrine aberrations in women with hypothalamic amenorrhea. J. Clin. Endocrinol. Metab. 1989, 68, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Reindollar, R.H.; Novak, M.; Tho, S.P.; McDonough, P.G. Adult-onset amenorrhea: A study of 262 patients. Am. J. Obstet. Gynecol. 1986, 155, 531–543. [Google Scholar] [CrossRef]
- McEwen, B.S. The neurobiology of stress: From serendipity to clinical relevance. Brain Res. 2000, 886, 172–189. [Google Scholar] [CrossRef]
- Berga, S.L.; Daniels, T.L.; Giles, D.E. Women with functional hypothalamic amenorrhea but not other forms of anovulation display amplified cortisol concentrations. Fertil. Steril. 1997, 67, 1024–1030. [Google Scholar] [CrossRef]
- Brundu, B.; Loucks, T.L.; Adler, L.J.; Cameron, J.L.; Berga, S.L. Increased cortisol in the cerebrospinal fluid of women with functional hypothalamic amenorrhea. J. Clin. Endocrinol. Metab. 2006, 91, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Marcus, M.D.; Loucks, T.L.; Berga, S.L. Psychological correlates of functional hypothalamic amenorrhea. Fertil. Steril. 2001, 76, 310–316. [Google Scholar] [CrossRef]
- Wuttke, W.; Pitzel, L.; Seidlova-Wuttke, D.; Hinney, B. LH pulses and the corpus luteum: The luteal phase deficiency (LPD). Vitam. Horm. 2001, 63, 131–158. [Google Scholar] [PubMed]
- Jain, A.; Polotsky, A.J.; Rochester, D.; Berga, S.L.; Loucks, T.; Zeitlian, G.; Gibbs, K.; Polotsky, H.N.; Feng, S.; Isaac, B.; et al. Pulsatile luteinizing hormone amplitude and progesterone metabolite excretion are reduced in obese women. J. Clin. Endocrinol. Metab. 2007, 92, 2468–2473. [Google Scholar] [CrossRef] [PubMed]
- Epel, E.S. Psychological and metabolic stress: A recipe for accelerated cellular aging? Hormones 2009, 8, 8–22. [Google Scholar] [CrossRef]
- Hanzel, C.E.; Pichet-Binette, A.; Pimentel, L.S.; Iulita, M.F.; Allard, S.; Ducatenzeiler, A.; do Carmoa, S.; Cuelloa, A.C. Neuronal driven pre-plaque inflammation in a transgenic rat model of Alzheimer’s disease. Neurobiol. Aging 2014, 35, 2249–2262. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, S.M.; Matzuk, M.M. The menstrual cycle—Basic biology. Ann. N. Y. Acad. Sci. 2008, 1135, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.C.; Roseboom, T.J.; de Rooij, S.R. Long-term effects of prenatal stress and glucocorticoid exposure. Birth Defects Res. Part C Embryo Today 2012, 96, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Dunkel Schetter, C.; Glynn, L.M. Stress in pregnancy: Empirical evidence and theoretical issues to guide interdisciplinary researchers. In Handbook of Stress; Contrada, R., Baum, A., Eds.; Springer: New York, NY, USA, 2011; pp. 321–343. [Google Scholar]
- Jones, S.A.; Challis, J.R. Steroid, corticotrophin-releasing hormone, ACTH and prostaglandin interactions in the amnion and placenta of early pregnancy in man. J. Endocrinol. 1990, 125, 153–159. [Google Scholar] [CrossRef] [PubMed]
- McLean, M.; Thompson, D.; Zhang, H.P.; Brinsmead, M.; Smith, R. Corticotrophin-releasing hormone and β-endorphin in labour. Eur. J. Endocrinol. 1994, 131, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.A.; Challism, J.R. Local stimulation of prostaglandin production by corticotropin-releasing hormone in human fetal membranes and placenta. Biochem. Biophys. Res. Commun. 1989, 159, 192–199. [Google Scholar] [CrossRef]
- Lockwood, C.J. Stress-associated preterm delivery: The role of corticotropin-releasing hormone. Am. J. Obstet. Gynecol. 1999, 180, S264–S266. [Google Scholar] [CrossRef]
- McLean, M.; Bisits, A.; Davies, J.; Waltres, W.; Hackshaw, A.; de Voss, K.; Smith, R. Predicting risk of preterm delivery by second-trimester measurement of maternal plasma corticotropin-releasing hormone and α-fetoprotein concentrations. Am. J. Obstet. Gynecol. 1999, 181, 207–215. [Google Scholar] [CrossRef]
- Wadhwa, P.D.; Porto, M.; Garite, T.J.; Chicz-DeMet, A.; Sandman, C.A. Maternal corticotropin-releasing hormone levels in the early third trimester predict length of gestation in human pregnancy. Am. J. Obstet. Gynecol. 1998, 179, 1079–1085. [Google Scholar] [CrossRef]
- Hobel, C.J.; Dunkel-Schetter, C.; Roesch, S.C.; Castro, L.C.; Chander, P.A. Maternal plasma corticotropin-releasing hormone associated with stress at 20 weeks’ gestation in pregnancies ending in preterm delivery. Am. J. Obstet. Gynecol. 1999, 180, S257–S263. [Google Scholar] [CrossRef]
- Korebrits, C.; Ramirez, M.M.; Watson, L.; Brinkman, E.; Bocking, A.D.; Challis, J.R.G. Maternal corticotropin-releasing hormone is increased with impending preterm birth. J. Clin. Endocrinol. Metab. 1998, 83, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Klima, C.S. Centering pregnancy: A model for pregnant adolescents. J. Midwifery Women’s Health 2003, 48, 220–225. [Google Scholar] [CrossRef]
- Picklesimer, A.H.; Billings, D.; Hale, N.; Blackhurst, D.; Covington-Kolb, S. The effect of Centering Pregnancy group prenatal care on preterm birth in a low-income population. Am. J. Obstet. Gynecol. 2012, 206, e1–e7. [Google Scholar] [CrossRef]
- Tandon, S.; Colon, L.; Vega, P.; Murphy, J.; Alonso, A. Birth outcomes associated with receipt of group prenatal care among low-income Hispanic women. J. Midwifery Women’s Health 2012, 57, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Ickovics, J.R.; Kershaw, T.S.; Westdahl, C.; Rising, S.S.; Klima, C.; Reynolds, H.; Magriples, U. Group prenatal care and preterm birth weight: Results from a matched cohort study at public clinics. Obstet. Gynecol. 2003, 102, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Yu, J.; Basanta-Henry, P.; Brou, L.; Berga, S.L.; Fortunato, S.J.; Taylor, R. Short fetal leukocyte telomere length and preterm prelabor rupture of the membranes. PLoS ONE 2012, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ballard, P.L.; Ballard, R.A. Scientific basis and therapeutic regimens for use of antenatal glucocorticoids. Am. J. Obstet. Gynecol. 1995, 173, 254–262. [Google Scholar] [CrossRef]
- Paganelli, S.; Soncini, E.; Gargano, G.; Capodanno, F.; Vezzani, C.; La Sala, G.B. Retrospective analysis on the efficacy of corticosteroid prophylaxis prior to elective caesarean section to reduce neonatal respiratory complications at term of pregnancy: Review of literature. Arch. Gynecol. Obstet. 2013, 288, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Seckl, J. Glucocorticoids, prenatal stress and the programming of disease. Horm. Behav. 2011, 59, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Seckl, J.R.; Meaney, M.J. Glucocorticoid “programming” and PTSD risk. Ann. N. Y. Acad. Sci. 2006, 1071, 351–378. [Google Scholar] [CrossRef] [PubMed]
- Seckl, J.R. Glucocorticoids, developmental “programming” and the risk of affective dysfunction. Prog. Brain Res. 2008, 167, 17–34. [Google Scholar] [PubMed]
- Seckl, J.R.; Holmes, M.C. Mechanisms of disease: Glucocorticoids, their placental metabolism and fetal “programming” of adult pathophysiology. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, J.P.; Acuna, G.; Rose, J.C.; Massmann, G.A. Maternal antenatal steroid administration at 0.55 gestation increases arterial blood pressure in young adult sheep offspring. J. Soc. Gynecol. Investig. 2004, 11, 358A. [Google Scholar]
- Contag, S.A.; Bi, J.; Chappell, M.C.; Rose, J.C. Developmental effect of antenatal exposure to betamethasone on renal angiotensin II activity in the young adult sheep. Am. J. Physiol. Ren. Physiol. 2010, 298, F847–F856. [Google Scholar] [CrossRef] [PubMed]
- Shaltout, H.A.; Rose, J.C.; Chappell, M.C.; Diz, D.I. Angiotensin-(1–7) deficiency and baroreflex impairment precede the antenatal β-methasone exposure-induced elevation in blood pressure. Hypertension 2012, 59, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Peña, C.J.; Monk, C.; Champagne, F.A. Epigenetic effects of prenatal stress on 11β-hydroxysteroid dehydrogenase-2 in the placenta and fetal brain. PLoS ONE 2012, 7, e39791. [Google Scholar]
- Hofmann, S.G.; Asnaani, A.; Vonk, I.J.J.; Sawyer, A.T.; Fang, A. The efficacy of cognitive behavioral therapy: A review of meta-analyses. Cogn. Ther. Res. 2012, 36, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Berga, S.L.; Loucks, T.L. Use of cognitive behavior therapy for functional hypothalamic amenorrhea. Ann. N. Y. Acad. Sci. 2006, 1092, 114–129. [Google Scholar] [CrossRef] [PubMed]
- Sterling, P.; Eyer, J. Allostasis: A new paradigm to explain arousal pathology. In Handbook of Life Stress, Cognition and Health; Fisher, S., Reason, J., Eds.; John Wiley & Sons: New York, NY, USA, 1988; pp. 629–649. [Google Scholar]
- Berga, S.L.; Marcus, M.D.; Loucks, T.L.; Hlastala, S.; Ringham, R.; Krohn, M.A. Recovery of ovarian activity in women with functional hypothalamic amenorrhea who were treated with cognitive behavior therapy. Fertil. Steril. 2003, 80, 976–981. [Google Scholar] [CrossRef]
- Michopoulos, V.; Mancini, F.; Loucks, T.L.; Berga, S.L. Neuroendocrine recovery initiated by cognitive behavioral therapy in women with functional hypothalamic amenorrhea: A randomized, controlled trial. Fertil. Steril. 2013, 99, 2084–2091. [Google Scholar] [CrossRef] [PubMed]
- Mohammadkhani, R.; Darbandi, N.; Vafaei, A.A.; Ahmadalipour, A.; Rashidy-Pour, A. Glucocorticoid-induced impairment of long-term memory retrieval in female rats: Influences of estrous cycle and estrogen. Neurobiol. Learn. Mem. 2015, 118, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Bolt, M.J.; Stossi, F.; Newberg, J.Y.; Orjalo, A.; Johansson, H.E.; Mancini, M.A. Coactivators enable glucocorticoid receptor recruitment to fine-tune estrogen receptor transcriptional responses. Nucleic Acids Res. 2013, 41, 4036–4048. [Google Scholar] [CrossRef] [PubMed]
- Whirledge, S.; Cidlowski, J.A. Estradiol antagonism of glucocorticoid-induced GILZ expression in human uterine epithelial cells and murine uterus. Endocrinology 2013, 154, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Whirledge, S.; Xiaojiang, X.; Cidlowski, J.A. Global gene expression analysis in human uterine epithelial cells defines new targets of glucocorticoid and estradiol antagonism. Biol. Reprod. 2013, 89, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Whirledge, S.; Cidlowski, J.A. A role for glucocorticoids in stress-impaired reproduction: Beyond the hypothalamus and pituitary. Endocrinology 2013, 154, 4450–4468. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. The neuroendocrinology of stress and aging: The glucocorticoid cascade hypothesis. Endocr. Rev. 1986, 7, 284–301. [Google Scholar] [CrossRef] [PubMed]
- Samuels, M.H.; Luther, M.; Henry, P.; Ridgway, E.C. Effects of hydrocortisone on pulsatile pituitary glycoprotein secretion. J. Clin. Endocrinol. Metab. 1994, 78, 211–215. [Google Scholar] [PubMed]
- Rozanski, A.; Kubzansky, L.D. Psychologic functioning and physical health: A paradigm of flexibility. Psychosom. Med. 2005, 67, S47–S53. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Protective and damaging effects of stress mediators. N. Engl. J. Med. 1998, 338, 171–179. [Google Scholar] [PubMed]
- Villa, A.; Vegeto, E.; Poletti, A.; Maggi, A. Estrogens, neuroinflammation, and neurodegeneration. Endocr. Rev. 2016, 37, 372–402. [Google Scholar] [CrossRef] [PubMed]
- Sierra, A.; Gottfried-Blackmore, A.; Milner, T.A.; McEwen, B.S.; Bulloch, K. Steroid hormone receptor expression and function in microglia. Glia 2008, 56, 659–674. [Google Scholar] [CrossRef] [PubMed]
- Vegeto, E.; Pollio, G.; Ciana, P.; Maggi, A. Estrogen blocks inducible nitric oxide synthase accumulation in LPS-activated microglia cells. Exp. Gerontol. 2000, 35, 1309–1316. [Google Scholar] [CrossRef]
- Soucy, G.; Boivin, G.; Labrie, F.; Rivest, S. Estradiol is required for a proper immune response to bacterial and viral pathogens in the female brain. J. Immunol. 2005, 174, 6391–6398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Nair, A.; Krady, K.; Corpe, C.; Bonneau, R.H.; Simpson, I.A.; Vannucci, S.J. Estrogen stimulates microglia and brain recovery from hypoxia-ischemia in normoglycemic but not diabetic female mice. J. Clin. Investig. 2004, 113, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.I.; Berga, S.L.; Cameron, J.L. Synergism between psychosocial and metabolic stressors: Impact on reproductive function in cynomolgus monkeys. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E270–E276. [Google Scholar] [CrossRef] [PubMed]
- Ekdahl, C.T.; Claasen, J.H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13632–13637. [Google Scholar] [CrossRef] [PubMed]
- Taupin, P. Adult neurogenesis, neuroinflammation and therapeutic potential of adult neural stem cells. Int. J. Med. Sci. 2008, 5, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L. Role of inflammation in neurodegenerative diseases. Curr. Opin. Neurol. 2005, 18, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Eikelenboom, P.; Veerhuis, R.; Scheper, W.; Rozemuller, A.J.; van Gool, W.A.; Hoozemans, J.J. The significance of neuroinflammation in understanding Alzheimer’s disease. J. Neural Transm. 2006, 113, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Latta, C.H.; Brothers, H.M.; Wilcock, D.M. Neuroinflammation in Alzheimer’s disease; a source of heterogeneity and target for personalized therapy. Neuroscience 2015, 302, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Vasto, S.; Candore, G.; Duro, G.; Lio, D.; Grimaldi, M.P.; Caruso, C. Alzheimer’s disease and genetics of inflammation: A pharmacogenomic vision. Pharmacogenomics 2007, 8, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, G.; van Heijningen, S.; Reijne, A.C.; Nyakas, C.; van der Zee, E.A.; Eisel, U.L. Integrative neurobiology of metabolic diseases, neuroinflammation, and neurodegeneration. Front. Neurosci. 2015. [Google Scholar] [CrossRef] [PubMed]

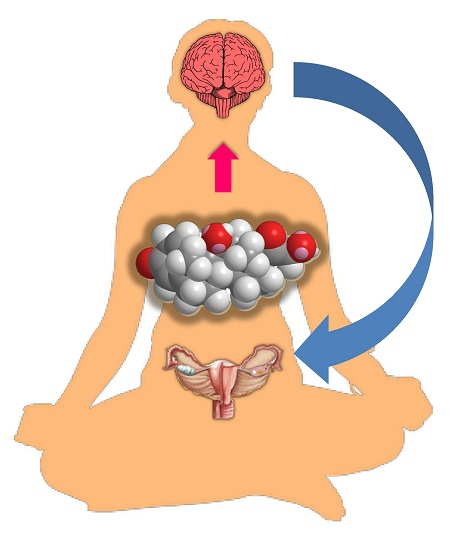{kind=link}
| Metabolic Aberrations in FHA that May Contribute to Long-Term Neurodegeneration |
|---|
| Elevated cortisol [9,10] |
| Hypoestrogenism [2] |
| Catabolism [51] |
| Hypothalamic hypothyroidism [6] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prokai, D.; Berga, S.L. Neuroprotection via Reduction in Stress: Altered Menstrual Patterns as a Marker for Stress and Implications for Long-Term Neurologic Health in Women. Int. J. Mol. Sci. 2016, 17, 2147. https://doi.org/10.3390/ijms17122147
Prokai D, Berga SL. Neuroprotection via Reduction in Stress: Altered Menstrual Patterns as a Marker for Stress and Implications for Long-Term Neurologic Health in Women. International Journal of Molecular Sciences. 2016; 17(12):2147. https://doi.org/10.3390/ijms17122147
Chicago/Turabian StyleProkai, David, and Sarah L. Berga. 2016. "Neuroprotection via Reduction in Stress: Altered Menstrual Patterns as a Marker for Stress and Implications for Long-Term Neurologic Health in Women" International Journal of Molecular Sciences 17, no. 12: 2147. https://doi.org/10.3390/ijms17122147
APA StyleProkai, D., & Berga, S. L. (2016). Neuroprotection via Reduction in Stress: Altered Menstrual Patterns as a Marker for Stress and Implications for Long-Term Neurologic Health in Women. International Journal of Molecular Sciences, 17(12), 2147. https://doi.org/10.3390/ijms17122147