
The Role of KV7.3 in Regulating Osteoblast Maturation and Mineralization


Abstract
:1. Introduction
2. Results
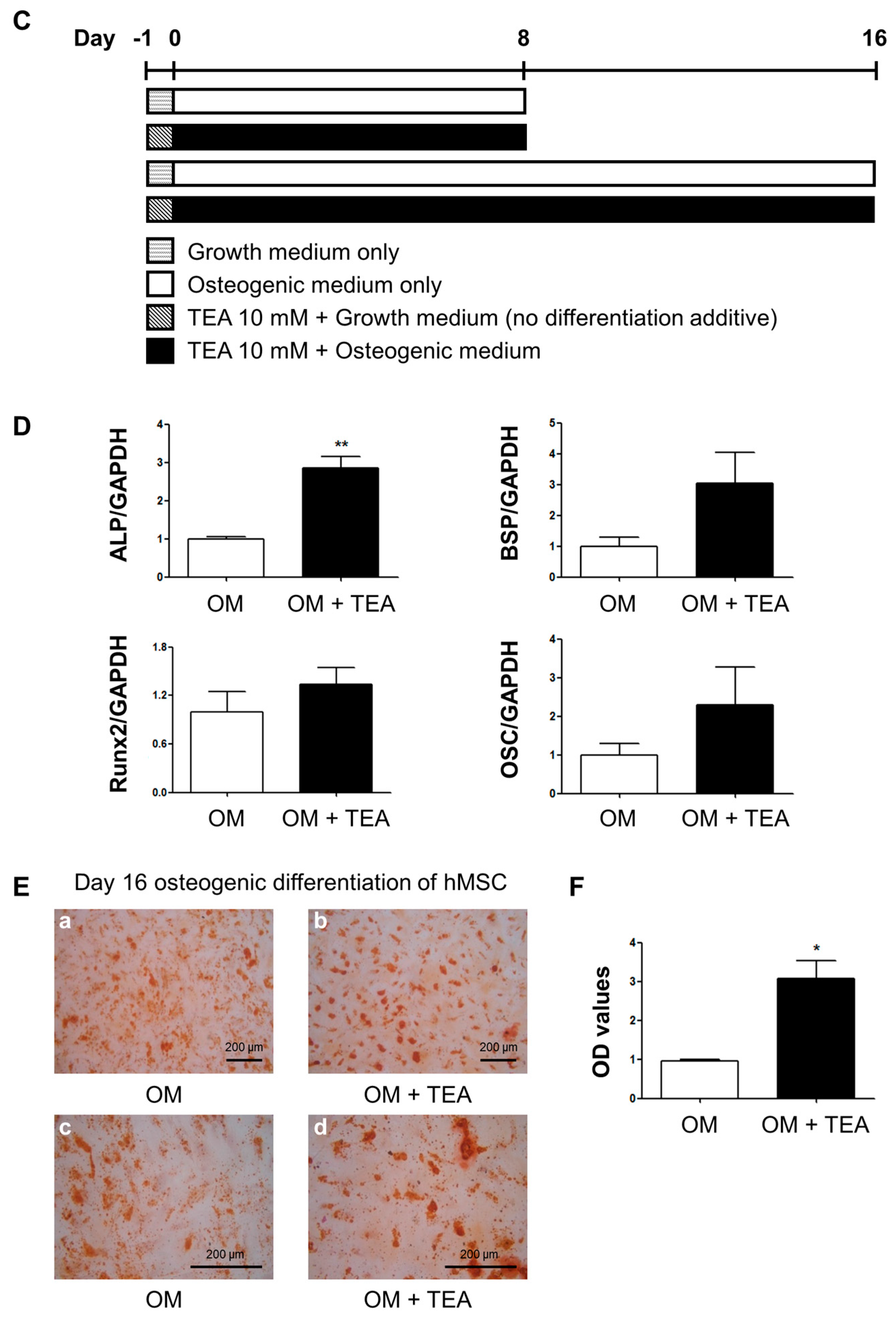
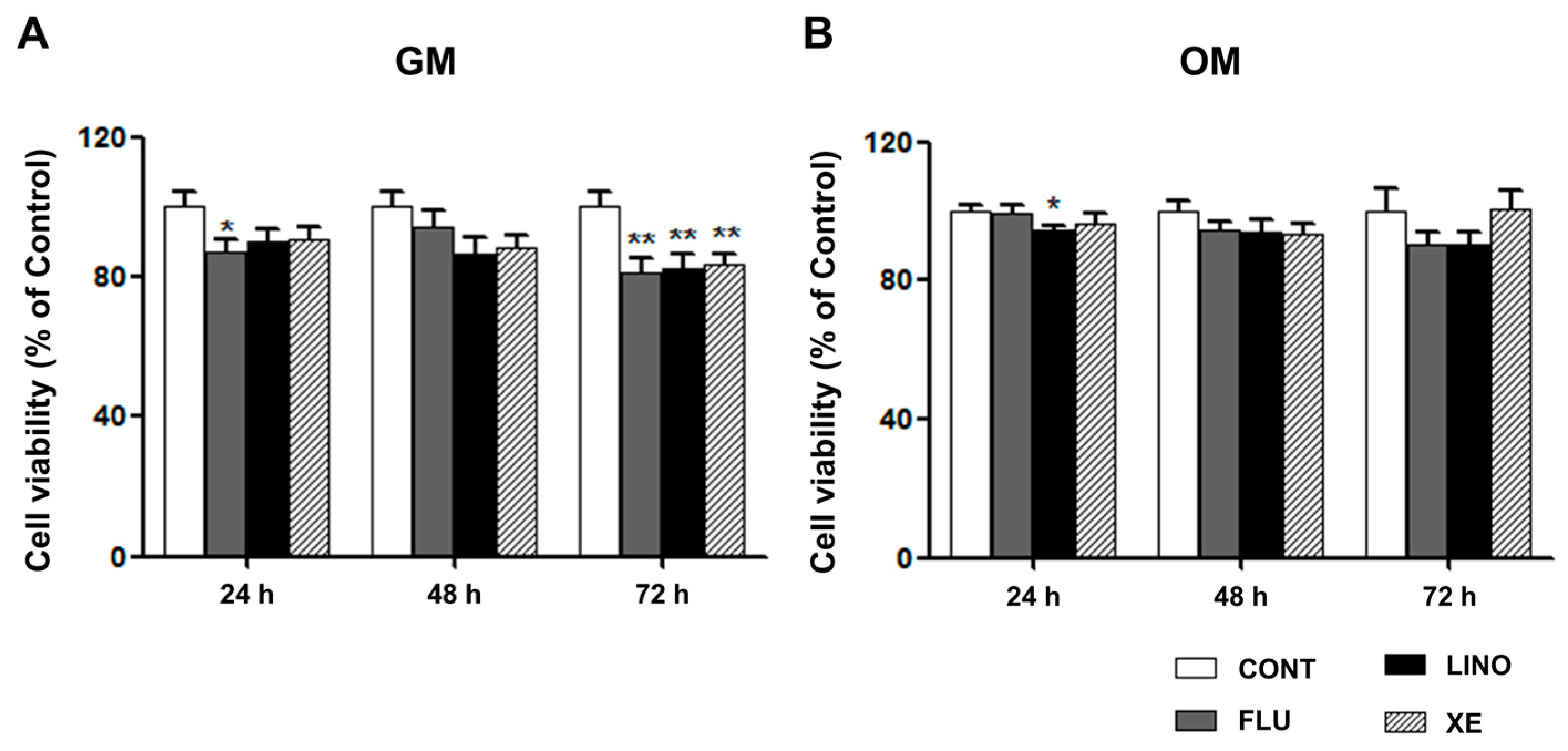
2.1. Effects of Tetraethylammonium, A Non-Selective Potassium Channel Blocker, on Human Mesenchymal Stem Cell Osteogenic Differentiation
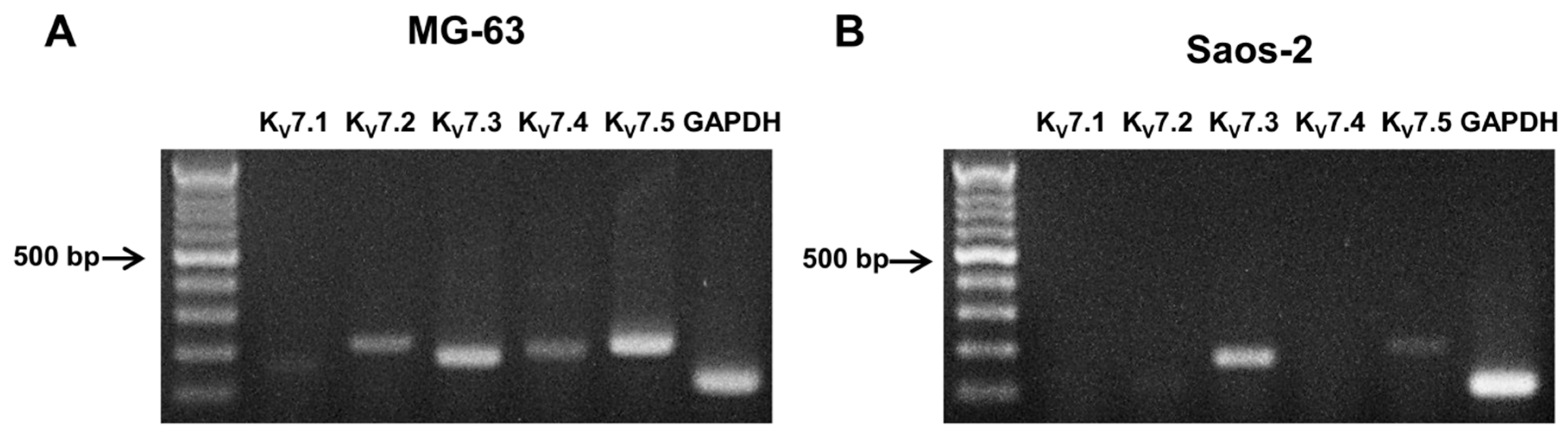
2.2. Expression of KV7 Channels in Osteoblast-Like Cell Lines, MG-63 and Saos-2
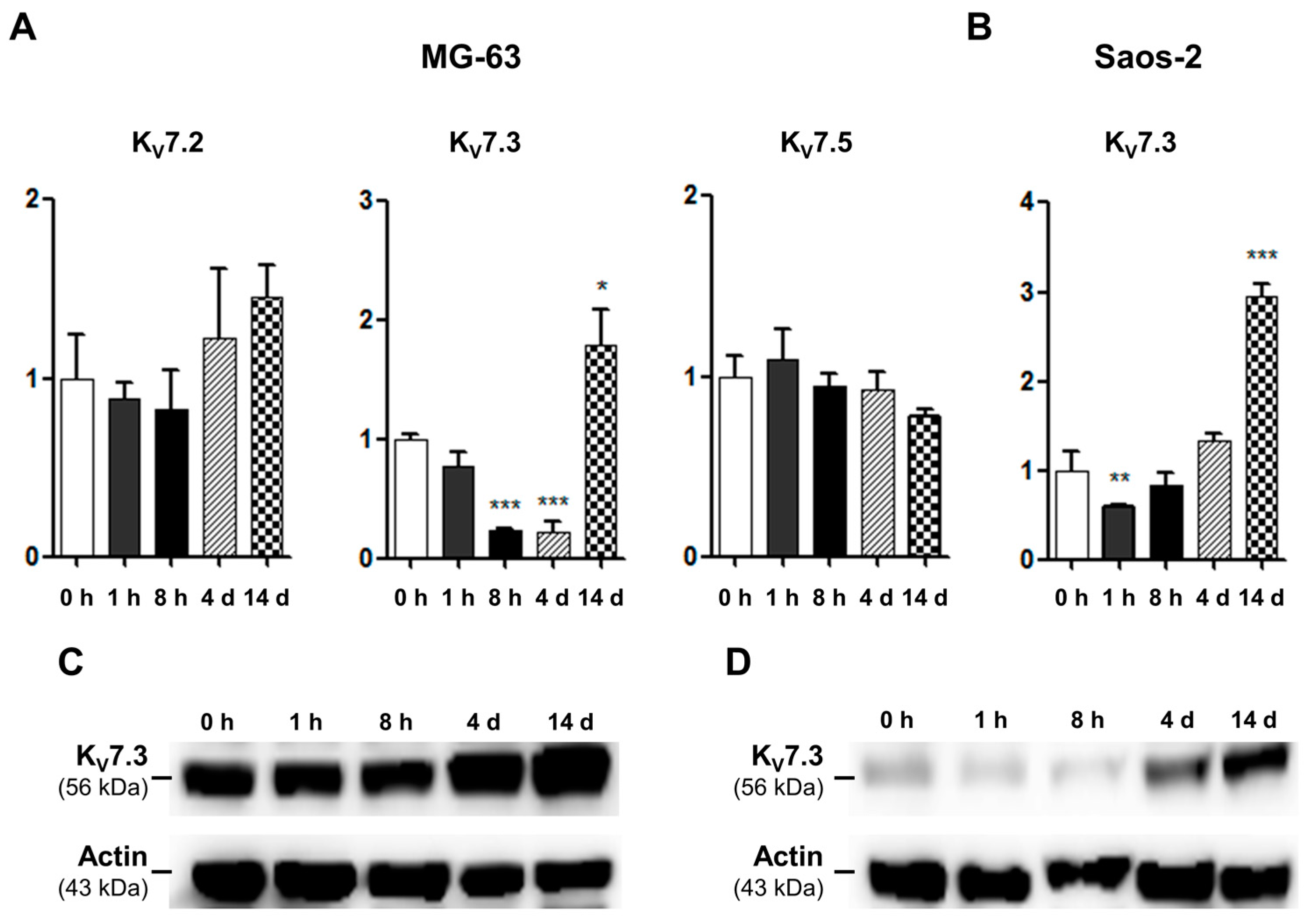
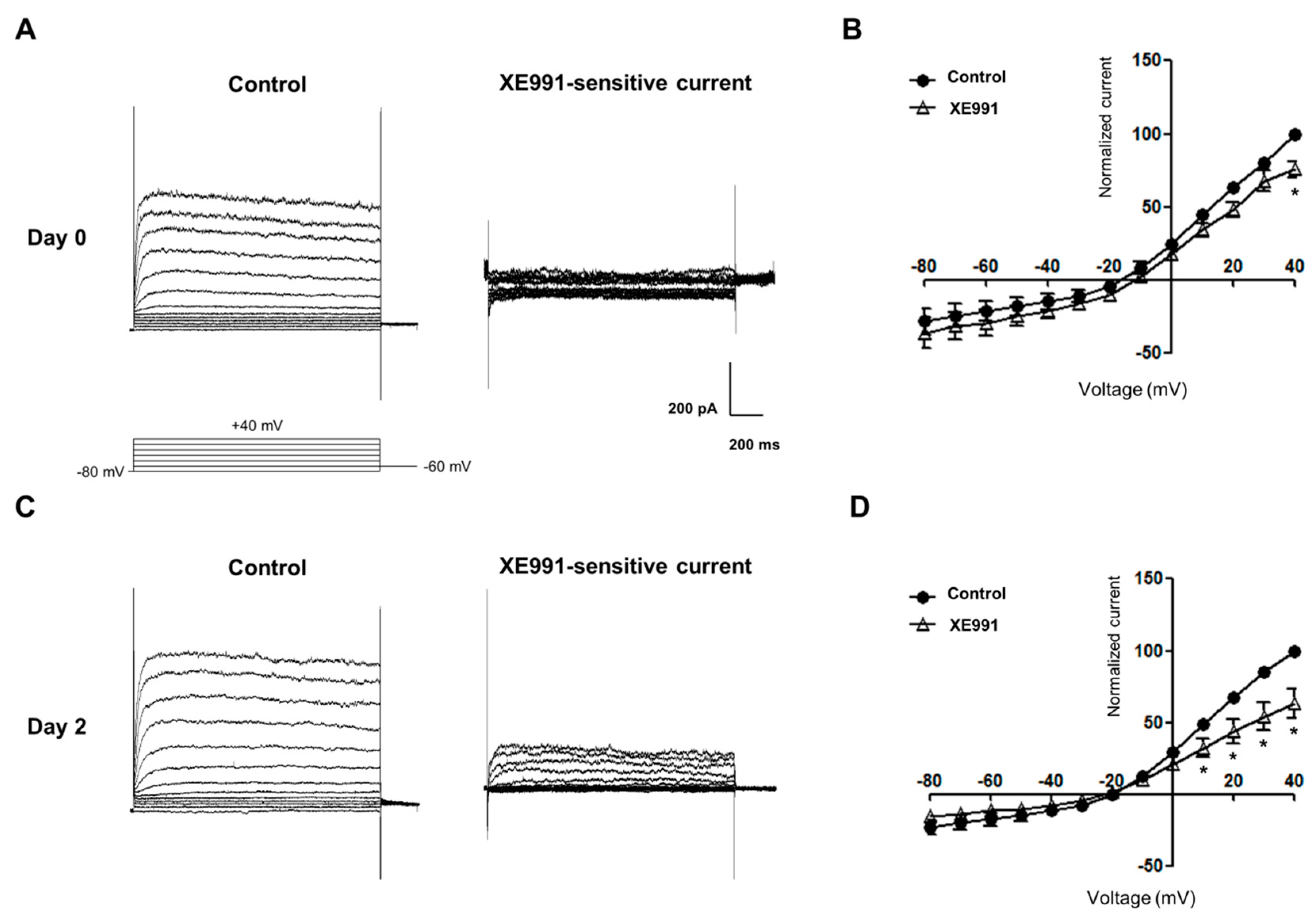
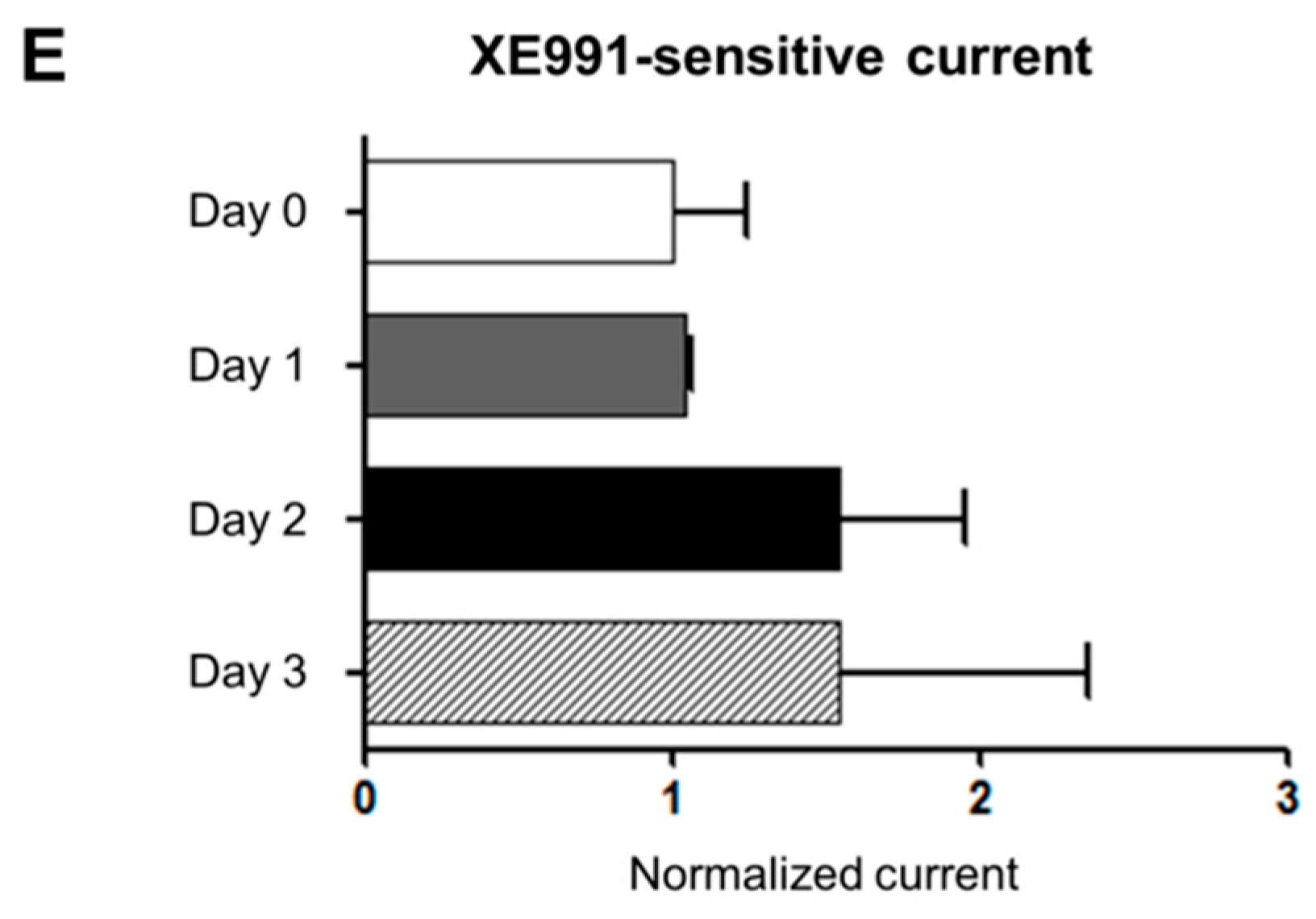
2.3. mRNA and Protein Expression and Functional Activities of Kv7 Channels during Osteoblastic Differentiation
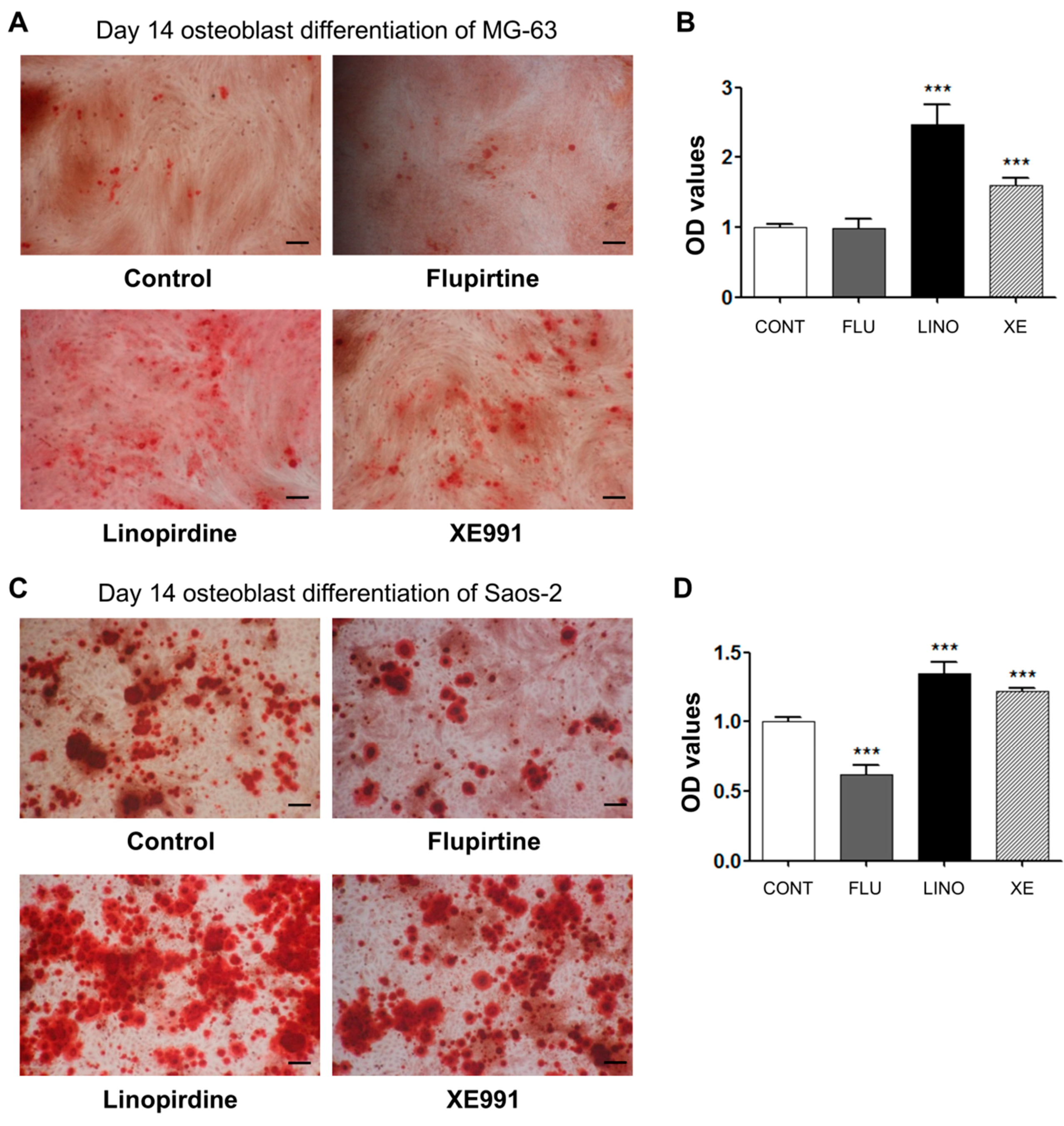
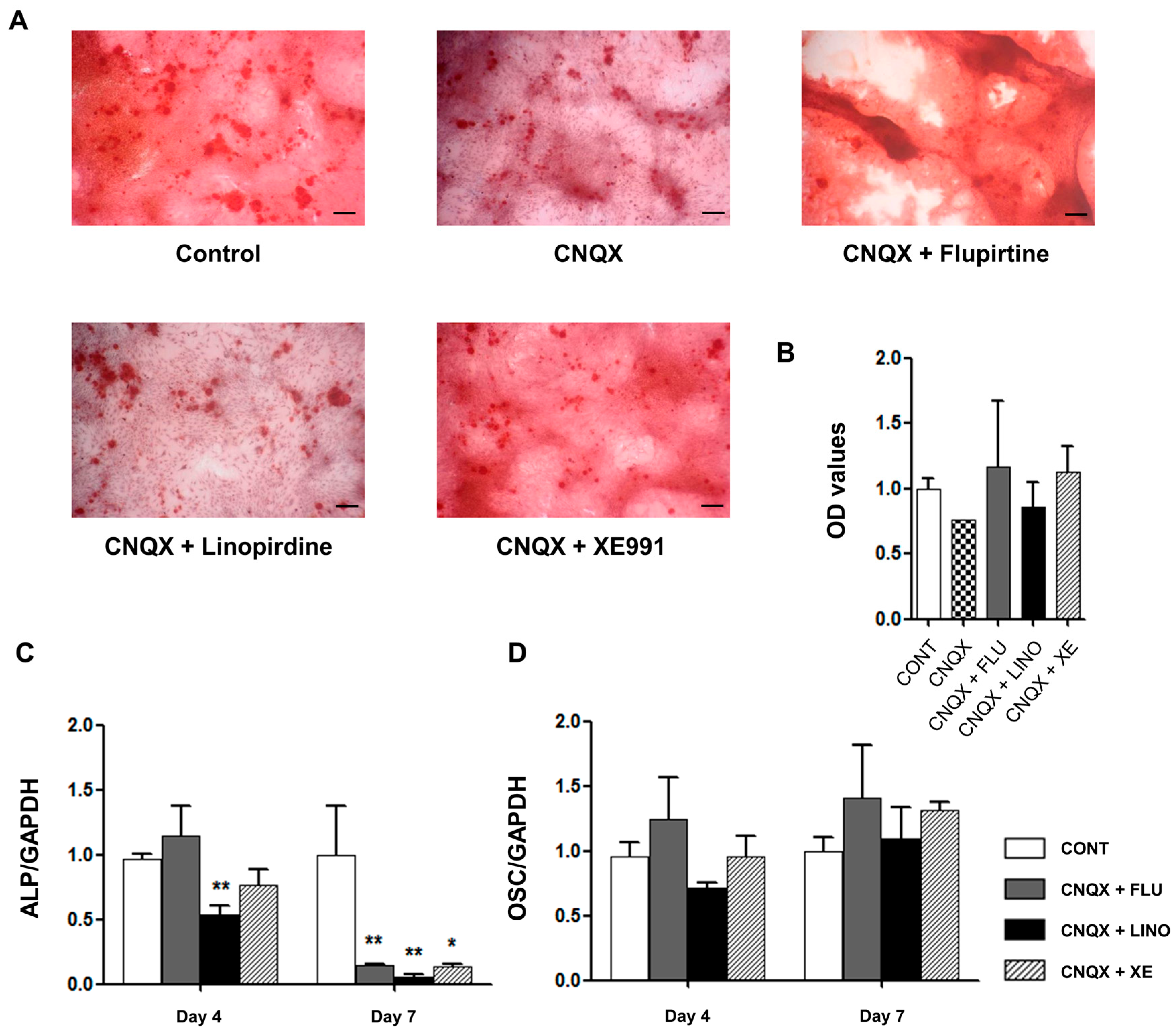
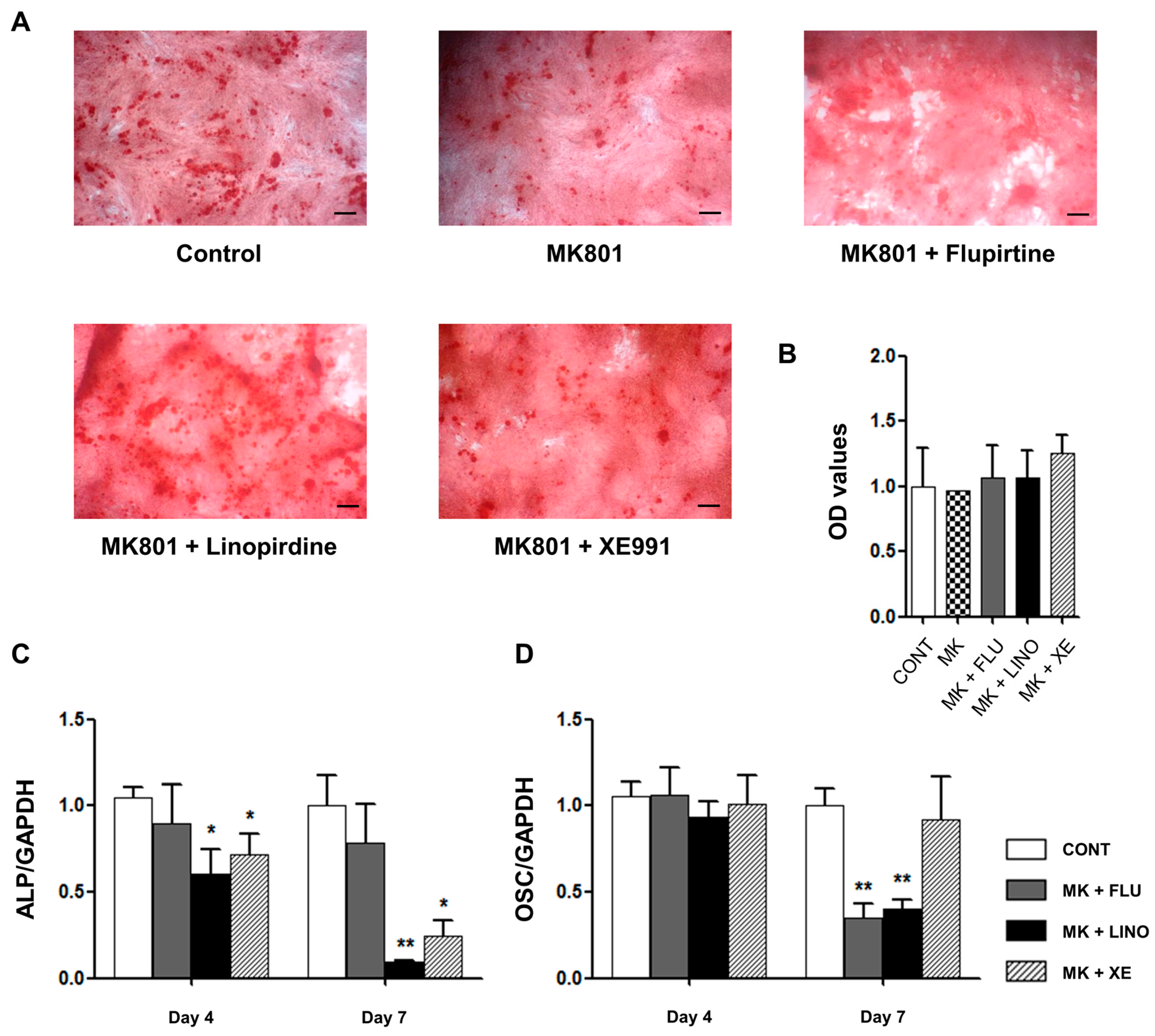
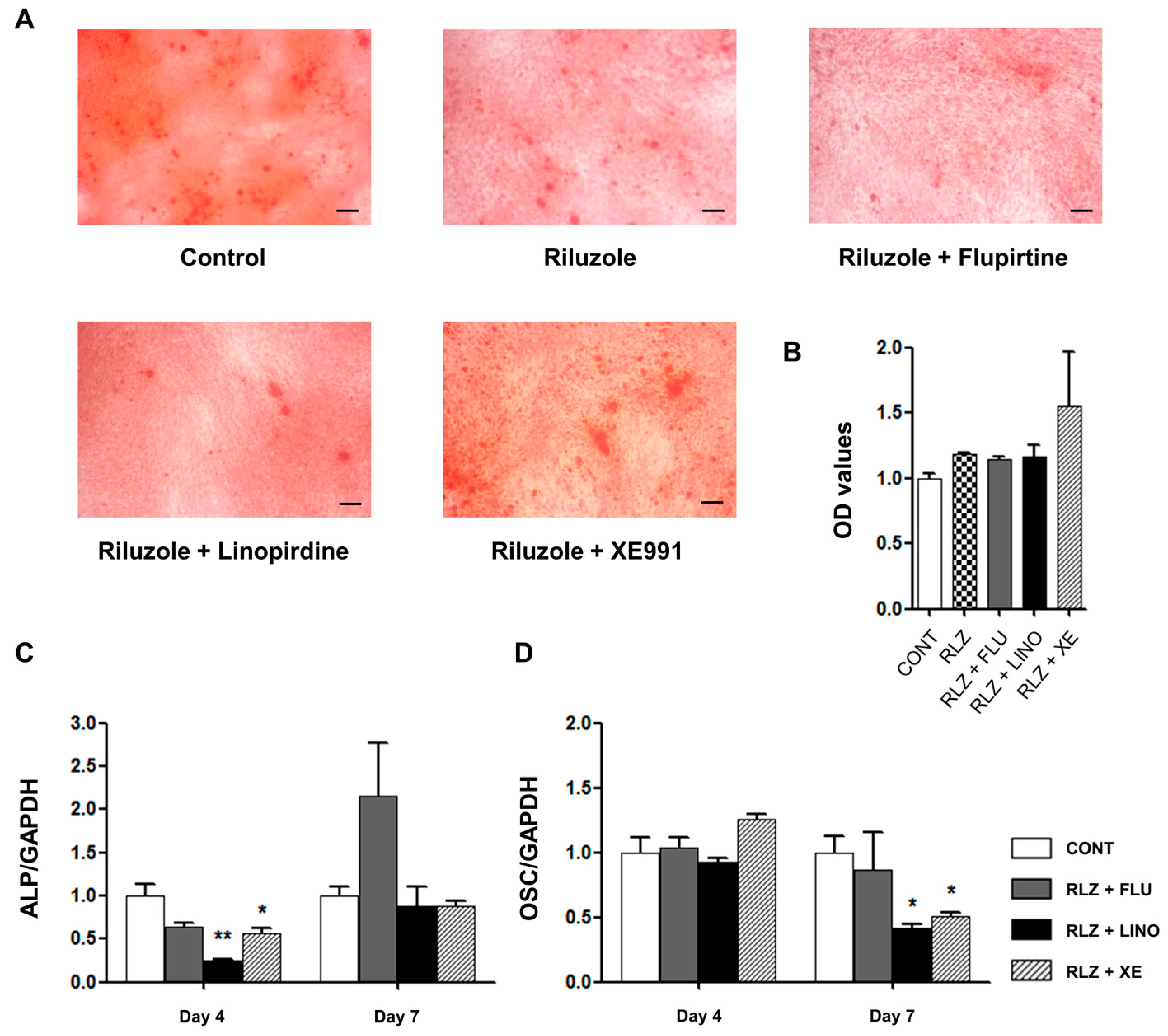
2.4. Regulation of Mineralization by KV7.3 Blockers or the KV7 Opener in MG-63 and Saos-2 Cells
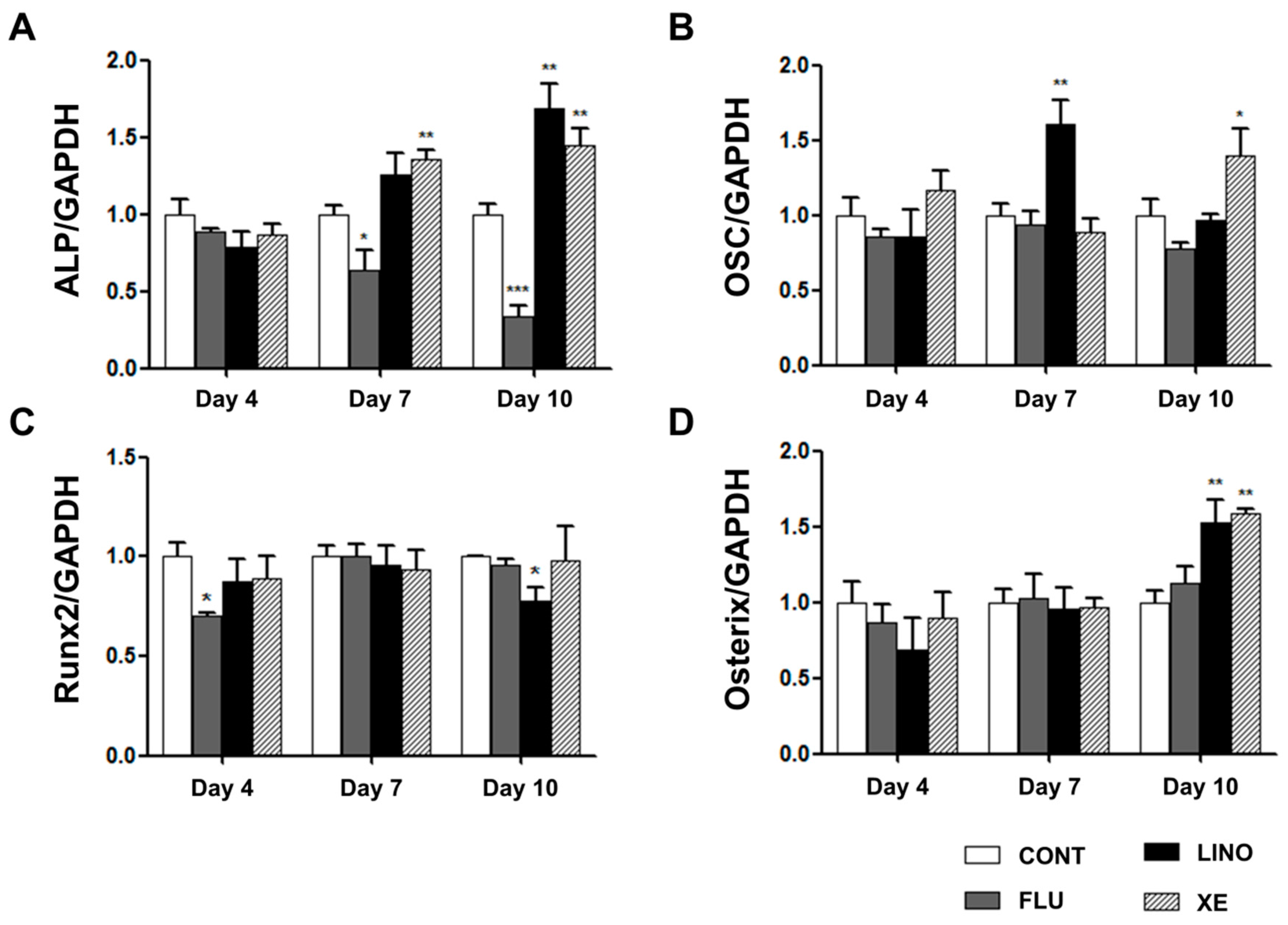
2.5. Regulation of Osteoblast Differentiation Markers by KV7 Channels in MG-63 Cells
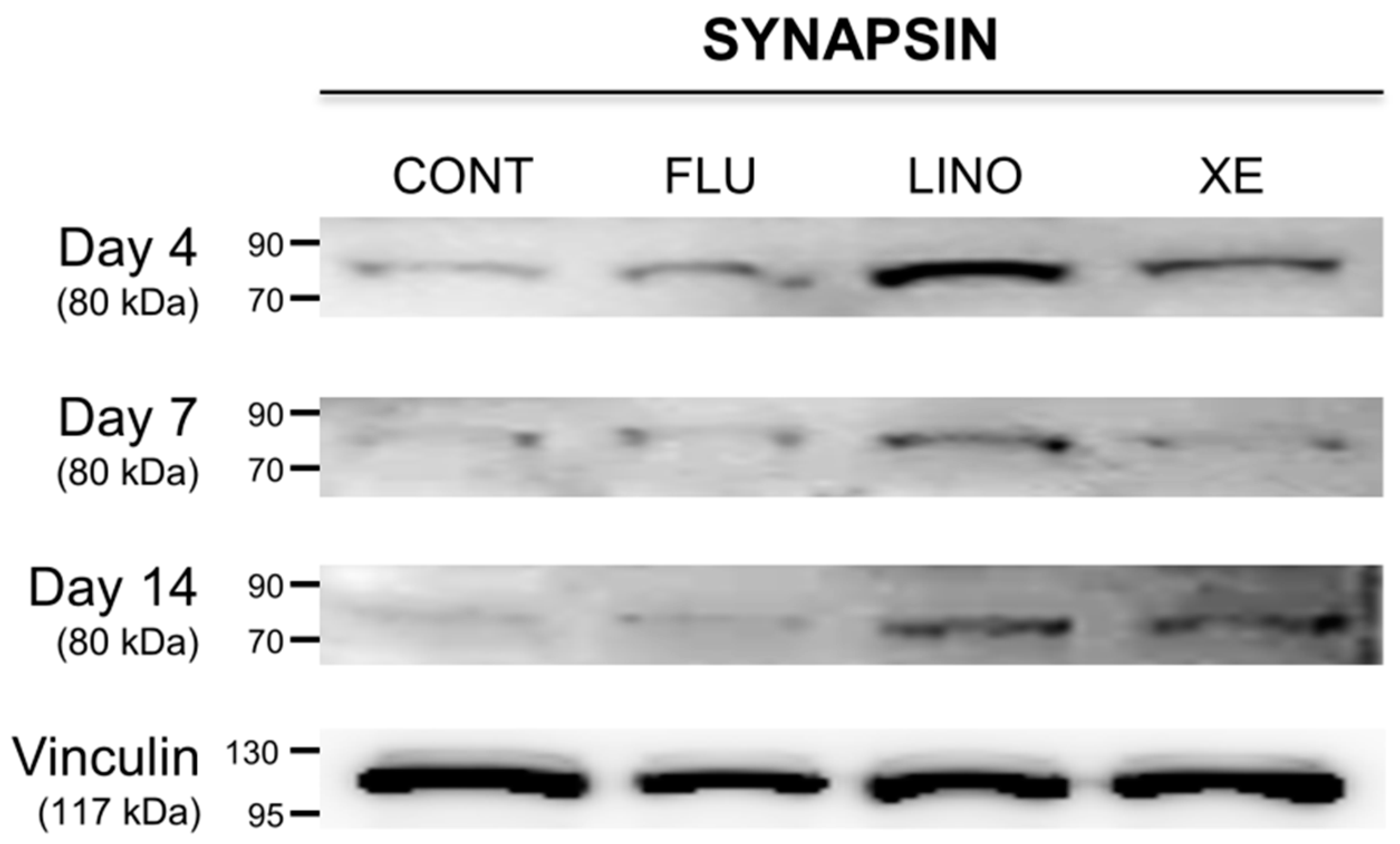
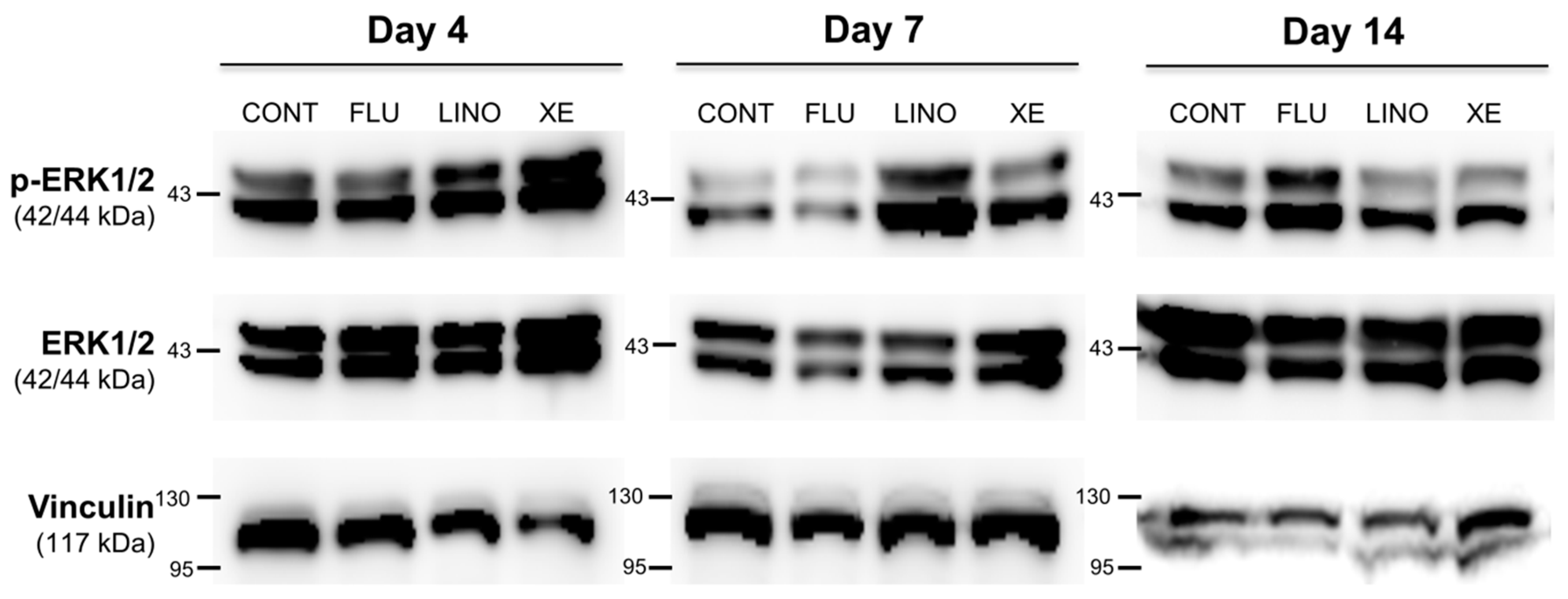
2.6. Effect of KV7 Channel Modulations on Synaptic Vesicle-Related Synapsin and the Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway
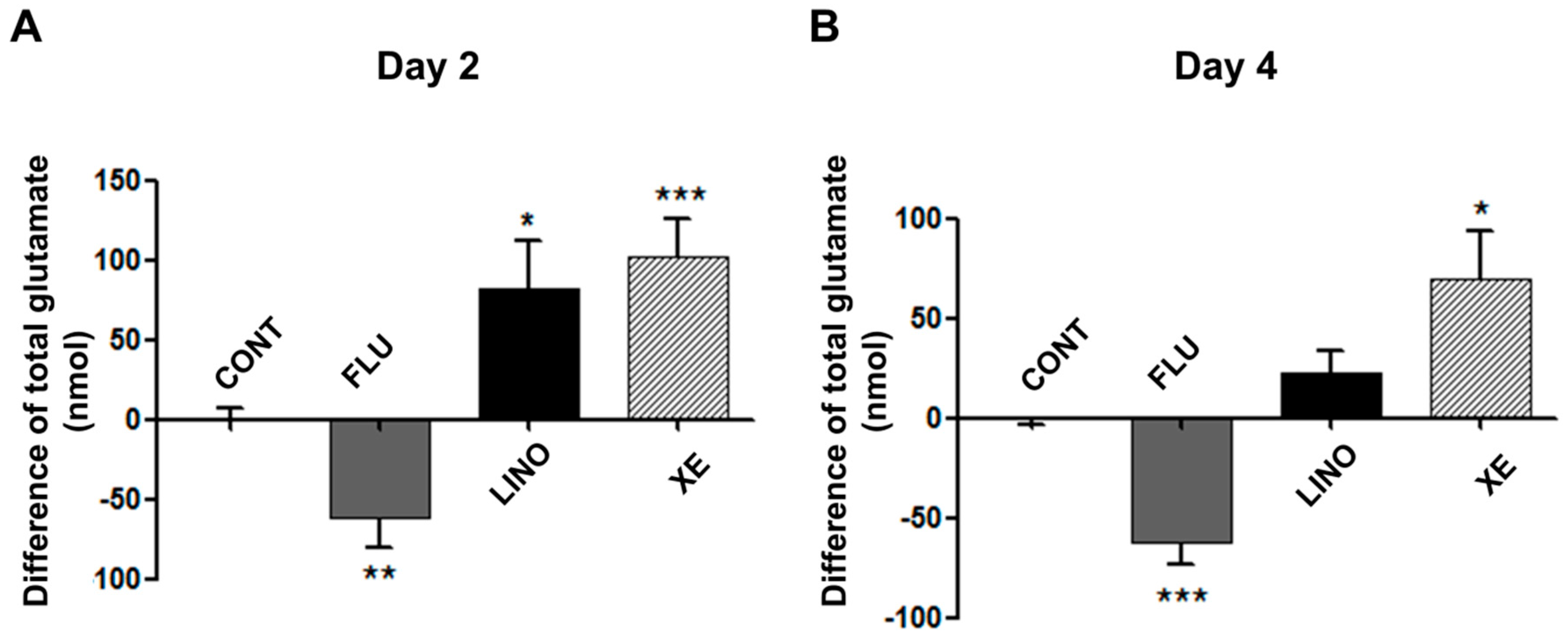
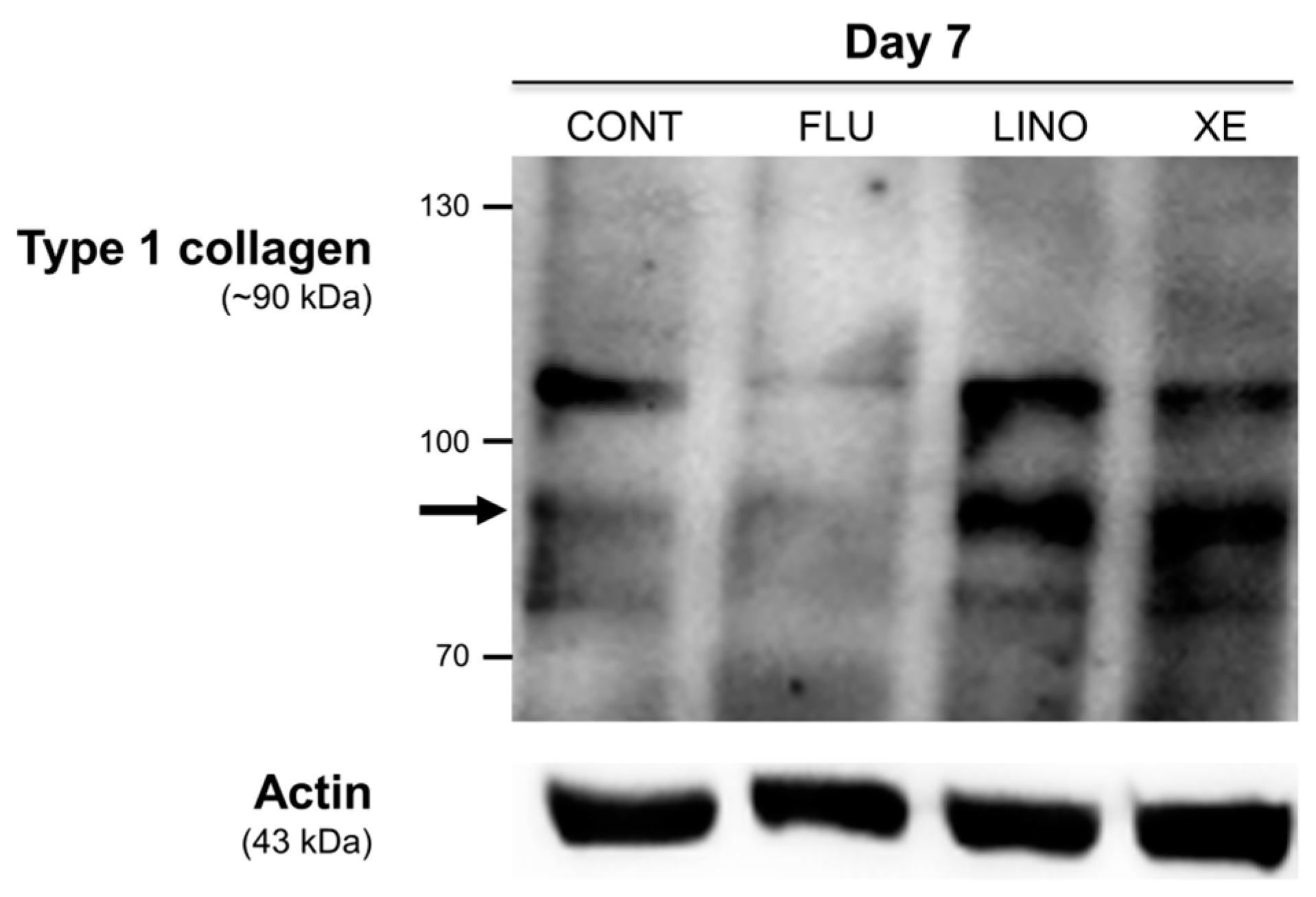
2.7. Induction of Glutamate Release and Type 1 Collagen by KV7.3 Channels during MG-63 Osteoblast Differentiation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Osteoblast Induction
4.3. Cell Viability Assay
4.4. RNA Extraction, RT-PCR and qRT-PCR
4.5. Alizarin Red S Staining and Quantification
4.6. Western Blot Analysis
4.7. Extracellular Glutamate Assay
4.9. Electrophysiological Recordings
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Misonou, H.; Mohapatra, D.P.; Trimmer, J.S. Kv2.1: A voltage-gated K+ channel critical to dynamic control of neuronal excitability. Neurotoxicology 2005, 26, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.K.; Newcombe, J.; Pryke, J.; Dolly, J.O. Subunit composition of Kv1 channels in human CNS. J. Neurochem. 1999, 73, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Nashmi, R.; Fehlings, M.G. Mechanisms of axonal dysfunction after spinal cord injury: With an emphasis on the role of voltage-gated potassium channels. Brain Res. Brain Res. Rev. 2001, 38, 165–191. [Google Scholar] [CrossRef]
- Bijlenga, P.; Occhiodoro, T.; Liu, J.H.; Bader, C.R.; Bernheim, L.; Fischer-Lougheed, J. An ether-a-go-go K+ current, Ih-eag, contributes to the hyperpolarization of human fusion-competent myoblasts. J. Physiol. 1998, 512, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Hancox, J.C.; McPate, M.J.; El Harchi, A.; Zhang, Y.H. The herg potassium channel and herg screening for drug-induced torsades de pointes. Pharmacol. Ther. 2008, 119, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Grunnet, M.; Hansen, R.S.; Olesen, S.P. Herg1 channel activators: A new anti-arrhythmic principle. Prog. Biophys. Mol. Biol. 2008, 98, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Martire, M.; Castaldo, P.; D’Amico, M.; Preziosi, P.; Annunziato, L.; Taglialatela, M. M channels containing KCNQ2 subunits modulate norepinephrine, aspartate, and gaba release from hippocampal nerve terminals. J. Neurosci. 2004, 24, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Martire, M.; D’Amico, M.; Panza, E.; Miceli, F.; Viggiano, D.; Lavergata, F.; Iannotti, F.A.; Barrese, V.; Preziosi, P.; Annunziato, L.; et al. Involvement of KCNQ2 subunits in [3H] dopamine release triggered by depolarization and pre-synaptic muscarinic receptor activation from rat striatal synaptosomes. J. Neurochem. 2007, 102, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Pillozzi, S.; Brizzi, M.F.; Balzi, M.; Crociani, O.; Cherubini, A.; Guasti, L.; Bartolozzi, B.; Becchetti, A.; Wanke, E.; Bernabei, P.A.; et al. Herg potassium channels are constitutively expressed in primary human acute myeloid leukemias and regulate cell proliferation of normal and leukemic hemopoietic progenitors. Leukemia 2002, 16, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Abdul, M.; Santo, A.; Hoosein, N. Activity of potassium channel-blockers in breast cancer. Anticancer Res. 2003, 23, 3347–3352. [Google Scholar] [PubMed]
- Jang, S.H.; Kang, K.S.; Ryu, P.D.; Lee, S.Y. Kv1.3 voltage-gated K+ channel subunit as a potential diagnostic marker and therapeutic target for breast cancer. BMB Rep. 2009, 42, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Ouadid-Ahidouch, H.; Chaussade, F.; Roudbaraki, M.; Slomianny, C.; Dewailly, E.; Delcourt, P.; Prevarskaya, N. Kv1.1 K+ channels identification in human breast carcinoma cells: Involvement in cell proliferation. Biochem. Biophys. Res. Commun. 2000, 278, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Vantol, B.; Cowley, E.; Blay, J.; Linsdell, P. Pharmacological separation of heag and herg K+ channel function in the human mammary carcinoma cell line MCF-7. Oncol. Rep. 2008, 19, 1511–1516. [Google Scholar] [PubMed]
- Spitzner, M.; Ousingsawat, J.; Scheidt, K.; Kunzelmann, K.; Schreiber, R. Voltage-gated K+ channels support proliferation of colonic carcinoma cells. FASEB J. 2007, 21, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Jiraporn, O.; Melanie, S.; Supaporn, P.; Luigi, T.; Luigi, T.; Lukas, B.; Karl, K.; Rainer, S. Expression of voltage-gated potassium channels in human and mouse colonic carcinoma. Clin. Cancer Res. 2007, 13, 824–831. [Google Scholar]
- Lastraioli, E.; Guasti, L.; Crociani, O.; Polvani, S.; Hofmann, G.; Witchel, H.; Bencini, L.; Calistri, M.; Messerini, L.; Scatizzi, M.; et al. Herg1 gene and herg1 protein are overexpressed in colorectal cancers and regulate cell invasion of tumor cells. Cancer Res. 2004, 64, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Fujii, T.; Takahashi, Y.; Takahashi, Y.; Suzuki, T.; Ukai, M.; Tauchi, K.; Horikawa, N.; Tsukada, K.; Sakai, H. Up-regulation of Kv7.1 channels in thromboxane A2-induced colonic cancer cell proliferation. Pflug. Arch. Eur.J. Physiol. 2014, 466, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Yongquan, S.; Zheyi, H.; Zhiming, H.; Yanglin, P.; Na, L.; Changcun, G.; Liu, H.; Jun, W.; Taidong, Q.; et al. Expression of delayed rectifier potassium channels and their possible roles in proliferation of human gastric cancer cells. Cancer Biol. Ther. 2005, 4, 1342–1347. [Google Scholar]
- Conforti, L.; Petrovic, M.; Mohammad, D.; Lee, S.; Ma, Q.; Barone, S.; Filipovich, A.H. Hypoxia regulates expression and activity of Kv1.3 channels in T lymphocytes: A possible role in T cell proliferation. J. Immunol. 2003, 170, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Wulff, H.; Calabresi, P.A.; Allie, R.; Yun, S.; Pennington, M.; Beeton, C.; Chandy, K.G. The voltage-gated Kv1.3 K+ channel in effector memory T cells as new target for ms. J. Clin. Investig. 2003, 111, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.H.; Choi, S.Y.; Ryu, P.D.; Lee, S.Y. Anti-proliferative effect of Kv1.3 blockers in A549 human lung adenocarcinoma in vitro and in vivo. Eur. J. Pharmacol. 2011, 651, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Panza, E.; Barrese, V.; Viggiano, D.; Soldovieri, M.V.; Taglialatela, M. Expression, localization, and pharmacological role of Kv7 potassium channels in skeletal muscle proliferation, differentiation, and survival after myotoxic insults. J. Pharmacol. Exp. Ther. 2010, 332, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Ryu, P.D.; Lee, S.Y. Serum starvation-induced voltage-gated potassium channel Kv7.5 expression and its regulation by Sp1 in canine osteosarcoma cells. Int. J. Mol. Sci. 2014, 15, 977–993. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, A.; Hofmann, G.; Pillozzi, S.; Guasti, L.; Crociani, O.; Cilia, E.; Di Stefano, P.; Degani, S.; Balzi, M.; Olivotto, M.; et al. Human ether-a-go-go-related gene 1 channels are physically linked to β1 integrins and modulate adhesion-dependent signaling. Mol. Biol. Cell 2005, 16, 2972–2983. [Google Scholar] [CrossRef] [PubMed]
- Levite, M.; Cahalon, L.; Peretz, A.; Hershkoviz, R.; Sobko, A.; Ariel, A.; Desai, R.; Attali, B.; Lider, O. Extracellular K+ and opening of voltage-gated potassium channels activate T cell integrin function: Physical and functional association between Kv1.3 channels and β1 integrins. J. Exp. Med. 2000, 191, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- You, M.H.; Song, M.S.; Lee, S.K.; Ryu, P.D.; Lee, S.Y.; Kim, D.Y. Voltage-gated K+ channels in adipogenic differentiation of bone marrow-derived human mesenchymal stem cells. Acta Pharmacol. Sin. 2013, 34, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Song, M.; Chen, D.; Wei, L.; Yu, S.P. Potential role of KCNQ/M-channels in regulating neuronal differentiation in mouse hippocampal and embryonic stem cell-derived neuronal cultures. Exp. Neurol. 2011, 229, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Jespersen, T.; Grunnet, M.; Olesen, S.P. The KCNQ1 potassium channel: From gene to physiological function. Physiology 2005, 20, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.M.; Huang, C.F.; Chao, C.C.; Lu, D.Y.; Kuo, C.S.; Cheng, T.H.; Chang, L.Y.; Chou, C.H. Voltage-gated K+ channels play a role in camp-stimulated neuritogenesis in mouse neuroblastoma N2A cells. J. Cell. Physiol. 2011, 226, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Soldovieri, M.V.; Miceli, F.; Taglialatela, M. Driving with no brakes: Molecular pathophysiology of Kv7 potassium channels. Physiology 2011, 26, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Flores, M.C.; Sihn, C.R.; Kim, H.J.; Zhang, Y.; Doyle, K.J.; Chiamvimonvat, N.; Zhang, X.D.; Yamoah, E.N. Identification of a key residue in Kv7.1 potassium channel essential for sensing external potassium ions. J. Gen. Physiol. 2015, 145, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; Osipenko, O.N.; Gurney, A.M. Properties of a novel K+ current that is active at resting potential in rabbit pulmonary artery smooth muscle cells. J. Physiol. 1996, 496, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Heitzmann, D.; Warth, R. No potassium, no acid: K+ channels and gastric acid secretion. Physiology 2007, 22, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Kao, T.; Horvath, Z.; Lemos, J.; Sul, J.Y.; Cranstoun, S.D.; Bennett, V.; Scherer, S.S.; Cooper, E.C. A common ankyrin-G-based mechanism retains KCNQ and Nav channels at electrically active domains of the axon. J. Neurosci. 2006, 26, 2599–2613. [Google Scholar] [CrossRef] [PubMed]
- Miceli, F.; Soldovieri, M.V.; Ambrosino, P.; Barrese, V.; Migliore, M.; Cilio, M.R.; Taglialatela, M. Genotype-phenotype correlations in neonatal epilepsies caused by mutations in the voltage sensor of Kv7.2 potassium channel subunits. Proc. Natl. Acd. Sci. USA 2013, 110, 4386–4391. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J. KCNQ potassium channels: Physiology, pathophysiology, and pharmacology. Pharmacol. Ther. 2001, 90, 1–19. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Barrese, V.; Formisano, L.; Miceli, F.; Taglialatela, M. Specification of skeletal muscle differentiation by repressor element-1 silencing transcription factor REST-regulated Kv7.4 potassium channels. Mol. Biol. Cell 2013, 24, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.C.; Hechenberger, M.; Weinreich, F.; Kubisch, C.; Jentsch, T.J. KCNQ5, a novel potassium channel broadly expressed in brain, mediates M-type currents. J. Biol. Chem. 2000, 275, 24089–24095. [Google Scholar] [CrossRef] [PubMed]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 2012, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Iniguez-Ariza, N.M.; Clarke, B.L. Bone biology, signaling pathways, and therapeutic targets for osteoporosis. Maturitas 2015, 82, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.; Li, Y.P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Kida, K.; Yamaguchi, A.; Hata, K.; Ichida, F.; Meguro, H.; Aburatani, H.; Nishimura, R.; Yoneda, T. BMP2 regulates osterix through Msx2 and Runx2 during osteoblast differentiation. J. Biol. Chem. 2008, 283, 29119–29125. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Stein, G.S.; Lian, J.B.; van Wijnen, A.J.; Stein, J.L.; Montecino, M.; Javed, A.; Zaidi, S.K.; Young, D.W.; Choi, J.Y.; Pockwinse, S.M. Runx2 control of organization, assembly and activity of the regulatory machinery for skeletal gene expression. Oncogene 2004, 23, 4315–4329. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell. Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Golub, E.E.; Boesze-Battaglia, K. The role of alkaline phosphatase in mineralization. Curr. Opin. Orthop. 2007, 18, 444–448. [Google Scholar] [CrossRef]
- Rohde, M.; Mayer, H. Exocytotic process as a novel model for mineralization by osteoblasts in vitro and in vivo determined by electron microscopic analysis. Calcif. Tissue Int. 2007, 80, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Marrion, N.V. Control of M-current. Annu. Rev. Physiol. 1997, 59, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Marrion, N.V.; Zucker, R.S.; Marsh, S.J.; Adams, P.R. Modulation of M-current by intracellular Ca2+. Neuron 1991, 6, 533–545. [Google Scholar] [CrossRef]
- Kosenko, A.; Hoshi, N. A change in configuration of the calmodulin-KCNQ channel complex underlies Ca2+-dependent modulation of KCNQ channel activity. PLoS ONE 2013, 8, e82290. [Google Scholar] [CrossRef] [PubMed]
- Haitin, Y.; Attali, B. The C-terminus of Kv7 channels: A multifunctional module. J. Physiol. 2008, 586, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.C.; Zaika, O.; Tolstykh, G.P.; Shapiro, M.S. Regulation of neural KCNQ channels: Signalling pathways, structural motifs and functional implications. J. Physiol. 2008, 586, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Barry, E.L. Expression of mRNAs for the α1 subunit of voltage-gated calcium channels in human osteoblast-like cell lines and in normal human osteoblasts. Calcif. Tissue Int. 2000, 66, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhong, D.; Lin, B.; Zhai, W.; Ding, Z.; Wu, J. p38 MAPK regulates the expression of ether-a-go-go potassium channel in human osteosarcoma cells. Radiol. Oncol. 2013, 47, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhong, D.; Gao, Q.; Zhai, W.; Ding, Z.; Wu, J. MicroRNA-34a inhibits human osteosarcoma proliferation by downregulating ether-a-go-go 1 expression. Int. J. Med. Sci. 2013, 10, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhong, D.; Fu, X.; Liu, Q.; Kang, L.; Ding, Z. Silencing of ether-a-go-go 1 by shRNA inhibits osteosarcoma growth and cell cycle progression. Int. J. Mol. Sci. 2014, 15, 5570–5581. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mao, Y.; Zhang, B.; Wang, T.; Li, F.; Fu, S.; Xue, Y.; Yang, T.; Wen, X.; Ding, Y.; et al. Chloride channel ClC-3 promotion of osteogenic differentiation through Runx2. J. Cell. Biochem. 2010, 111, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Jung, J.Y.; Cho, S.W.; Choi, H.J.; Kim, S.W.; Kim, S.Y.; Kim, H.J.; Jang, C.H.; Lee, M.G.; Han, J.; et al. Chloride intracellular channel 1 regulates osteoblast differentiation. Bone 2009, 45, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Sacco, S.; Giuliano, S.; Sacconi, S.; Desnuelle, C.; Barhanin, J.; Amri, E.Z.; Bendahhou, S. The inward rectifier potassium channel Kir2.1 is required for osteoblastogenesis. Hum. Mol. Genet. 2015, 24, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Henney, N.C.; Bo, L.; Elford, C.; Reviriego, P.; Campbell, A.K.; Wann, K.T.; Evans, B.A.J. A large-conductance BK potassium channel subtype affects both growth and mineralization of human osteoblasts. Am. J. Physiol. Cell. Physiol. 2009, 297, C1397–C1408. [Google Scholar] [CrossRef] [PubMed]
- Rezzonico, R.; Cayatte, C.; Bourget-Ponzio, I.; Romey, G.; Belhacene, N.; Loubat, A.; Rocchi, S.; van Obberghen, E.; Girault, J.A.; Rossi, B.; et al. Focal adhesion kinase pp125FAK interacts with the large conductance calcium-activated hSlo potassium channel in human osteoblasts: Potential role in mechanotransduction. J. Bone Miner. Res. 2003, 18, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [PubMed]
- Pautke, C.; Schieker, M.; Tischer, T.; Kolk, A.; Neth, P.; Mutschler, W.; Milz, S. Characterization of osteosarcoma cell lines MG-63, Saos-2 and U-2 OS in comparison to human osteoblasts. Anticancer Res. 2004, 24, 3743–3748. [Google Scholar] [PubMed]
- Prideaux, M.; Wijenayaka, A.R.; Kumarasinghe, D.D.; Ormsby, R.T.; Evdokiou, A.; Findlay, D.M.; Atkins, G.J. Saos2 osteosarcoma cells as an in vitro model for studying the transition of human osteoblasts to osteocytes. Calcif. Tissue Int. 2014, 95, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar] [PubMed]
- Yellowley, C.E.; Hancox, J.C.; Skerry, T.M.; Levi, A.J. Whole-cell membrane currents from human osteoblast-like cells. Calcif. Tissue Int. 1998, 62, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, S.; Dong, X.; Xiao, J. 17β-estradiol inhibits outward voltage-gated K+ currents in human osteoblast-like MG63 cells. J. Membr. Biol. 2013, 246, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sanchez, A.; Rubio, M.E.; Kohl, T.; Pardo, L.A.; Stuhmer, W. Functional Kv10.1 channels localize to the inner nuclear membrane. PLoS ONE 2011, 6, e19257. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.H.; Byun, J.K.; Jeon, W.I.; Choi, S.Y.; Park, J.; Lee, B.H.; Yang, J.E.; Park, J.B.; O’Grady, S.M.; Kim, D.Y.; et al. Nuclear localization and functional characteristics of voltage-gated potassium channel Kv1.3. J. Biol. Chem. 2015, 290, 12547–12557. [Google Scholar] [CrossRef] [PubMed]
- Szabo, I.; Bock, J.; Grassme, H.; Soddemann, M.; Wilker, B.; Lang, F.; Zoratti, M.; Gulbins, E. Mitochondrial potassium channel Kv1.3 mediates Bax-induced apoptosis in lymphocytes. Proc. Natl. Acad. Sci. USA 2008, 105, 14861–14866. [Google Scholar] [CrossRef] [PubMed]
- Swayne, L.A.; Wicki-Stordeur, L. Ion channels in postnatal neurogenesis: Potential targets for brain repair. Channels 2012, 6, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Sekiguchi, A.; Iwasaki, Y.K.; Sagara, K.; Iinuma, H.; Hatano, S.; Fu, L.T.; Watanabe, H. Circadian variation of cardiac K+ channel gene expression. Circulation 2003, 107, 1917–1922. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Hadley, J.K.; Noda, M.; Selyanko, A.A.; Wood, I.C.; Abogadie, F.C.; Brown, D.A. Differential tetraethylammonium sensitivity of KCNQ1-4 potassium channels. Br. J. Pharmacol. 2000, 129, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, Z.; Yoshida, C.A.; Furuichi, T.; Amizuka, N.; Ito, M.; Fukuyama, R.; Miyazaki, T.; Kitaura, H.; Nakamura, K.; Fujita, T.; et al. Runx2 determines bone maturity and turnover rate in postnatal bone development and is involved in bone loss in estrogen deficiency. Dev. Dyn. 2007, 236, 1876–1890. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Toyosawa, S.; Furuichi, T.; Kanatani, N.; Yoshida, C.; Liu, Y.; Himeno, M.; Narai, S.; Yamaguchi, A.; Komori, T. Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures. J. Cell Biol. 2001, 155, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Kanatani, N.; Fujita, T.; Fukuyama, R.; Liu, W.; Yoshida, C.A.; Moriishi, T.; Yamana, K.; Miyazaki, T.; Toyosawa, S.; Komori, T. Cbf β regulates Runx2 function isoform-dependently in postnatal bone development. Dev. Biol. 2006, 296, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor Osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.; Feng, J.Q.; Dusevich, V.M.; Sinha, K.; Zhang, H.; Darnay, B.G.; de Crombrugghe, B. Multiple functions of osterix are required for bone growth and homeostasis in postnatal mice. Proc. Natl. Acad. Sci. USA 2010, 107, 12919–12924. [Google Scholar] [CrossRef] [PubMed]
- Stein, G.S.; Lian, J.B.; Owen, T.A. Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. FASEB J. 1990, 4, 3111–3123. [Google Scholar] [PubMed]
- Said, S.I.; Berisha, H.I.; Pakbaz, H. Excitotoxicity in the lung: N-methyl-d-aspartate-induced, nitric oxide-dependent, pulmonary edema is attenuated by vasoactive intestinal peptide and by inhibitors of poly(ADP-ribose) polymerase. Proc. Natl. Acad. Sci. USA 1996, 14, 4688–4692. [Google Scholar] [CrossRef]
- Genever, P.G.; Wilkinson, D.J.; Patton, A.J.; Peet, N.M.; Hong, Y.; Mathur, A.; Erusalimsky, J.D.; Skerry, T. Expression of a functional N-methyl-d-aspartate-type glutamate receptor by bone marrow megakaryocytes. Blood 1999, 93, 2876–2883. [Google Scholar] [PubMed]
- Inagaki, N.; Kuromi, H.; Gonoi, T.; Okamoto, Y.; Ishida, H.; Seino, Y.; Kaneko, T.; Iwanaga, T.; Seino, S. Expression and role of ionotropic glutamate receptors in pancreatic islet cells. FASEB J. 1995, 9, 686–691. [Google Scholar] [PubMed]
- Weaver, C.D.; Yao, T.L.; Powers, A.C.; Verdoorn, T.A. Differential expression of glutamate receptor subtypes in rat pancreatic islets. J. Biol. Chem. 1996, 271, 2977–2984. [Google Scholar]
- Molnár, E.; Váradi, A.; McIlhinney, R.A.; Ashcroft, S.J. Identification of functional ionotropic glutamate receptor proteins in pancreatic β-cells and in islets of langerhans. FEBS Lett. 1995, 371, 253–257. [Google Scholar] [PubMed]
- Lin, T.H.; Yang, R.S.; Tang, C.H.; Wu, M.Y.; Fu, W.M. Regulation of the maturation of osteoblasts and osteoclastogenesis by glutamate. Eur. J. Pharmacol. 2008, 589, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Seidlitz, E.P.; Sharma, M.K.; Singh, G. Extracellular glutamate alters mature osteoclast and osteoblast functions. Cad. J. Physiol. Pharmacol. 2010, 88, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Chenu, C. Glutamatergic innervation in bone. Microsc Res. Tech. 2002, 58, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Brakspear, K.S.; Mason, D.J. Glutamate signaling in bone. Front. Endocrinol. 2012, 3, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Genever, P.G.; Skerry, T.M. Regulation of spontaneous glutamate release activity in osteoblastic cells and its role in differentiation and survival: Evidence for intrinsic glutamatergic signaling in bone. FASEB J. 2001, 15, 1586–1588. [Google Scholar] [CrossRef] [PubMed]
- Cowan, R.W.; Seidlitz, E.P.; Singh, G. Glutamate signaling in healthy and diseased bone. Front. Endocrinol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Kalariti, N.; Lembessis, P.; Koutsilieris, M. Characterization of the glutametergic system in MG-63 osteoblast-like osteosarcoma cells. Anticancer Res. 2004, 24, 3923–3929. [Google Scholar] [PubMed]
- Blum, R.; Konnerth, A. Neurotrophin-mediated rapid signaling in the central nervous system: Mechanisms and functions. Physiology 2005, 20, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Jie-Li, L.; Bin, C.; Li, Q.; Xiao-Ying, L.; Lian-Fu, D.; Guang, N.; Jian-Min, L. NMDA enhances stretching-induced differentiation of osteoblasts through the ERK1/2 signaling pathway. Bone 2008, 43, 469–475. [Google Scholar]
- Bhangu, P.S.; Genever, P.G.; Spencer, G.J.; Grewal, T.S.; Skerry, T.M. Evidence for targeted vesicular glutamate exocytosis in osteoblasts. Bone 2001, 29, 16–23. [Google Scholar] [CrossRef]
- Zhou, X.; Wei, J.; Song, M.; Francis, K.; Yu, S.P. Novel role of KCNQ2/3 channels in regulating neuronal cell viability. Cell. Death Differ. 2011, 18, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Kihara, T.; Hirose, M.; Oshima, A.; Ohgushi, H. Exogenous type I collagen facilitates osteogenic differentiation and acts as a substrate for mineralization of rat marrow mesenchymal stem cells in vitro. Biochem. Biophys. Res. Commun. 2006, 341, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Kuboki, Y. Osteoblast-related gene expression of bone marrow cells during the osteoblastic differentiation induced by type I collagen. J. Biochem. 2001, 129, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Salasznyk, R.M.; Williams, W.A.; Boskey, A.; Batorsky, A.; Plopper, G.E. Adhesion to vitronectin and collagen I promotes osteogenic differentiation of human mesenchymal stem cells. J. Biomed. Biotechnol. 2004, 2004, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Viguet-Carrin, S.; Garnero, P.; Delmas, P.D. The role of collagen in bone strength. Osteoporos. Int. 2006, 17, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Mathews, S.; Bhonde, R.; Gupta, P.K.; Totey, S. A novel tripolymer coating demonstrating the synergistic effect of chitosan, collagen type I and hyaluronic acid on osteogenic differentiation of human bone marrow derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2011, 414, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Gajko-Galicka, A. Mutations in type I collagen genes resulting in osteogenesis imperfecta in humans. Acta Biochim. Pol. 2002, 49, 433–441. [Google Scholar] [PubMed]
- Basel, D.; Steiner, R.D. Osteogenesis imperfecta: Recent findings shed new light on this once well-understood condition. Genet. Med. 2009, 11, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Desiderio, V.; Tirino, V.; Papaccio, G.; Paino, F. Bone defects: Molecular and cellular therapeutic targets. Int. J. Biochem. Cell. Biol. 2014, 51, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Maraka, S.; Kennel, K.A. Bisphosphonates for the prevention and treatment of osteoporosis. BMJ 2015, 351, h3783. [Google Scholar] [CrossRef] [PubMed]
- Drake, M.T.; Clarke, B.L.; Khosla, S. Bisphosphonates: Mechanism of action and role in clinical practice. Mayo Clin. Proc. 2008, 83, 1032–1045. [Google Scholar] [CrossRef] [PubMed]
- Komarova, S.V.; Dixon, S.J.; Sims, S.M. Osteoclast ion channels: Potential targets for antiresorptive drugs. Curr. Pharm. Des. 2001, 7, 637–654. [Google Scholar] [CrossRef] [PubMed]
- Nardone, V.; D’Asta, F.; Brandi, M.L. Pharmacological management of osteogenesis. Clinics 2014, 69, 438–446. [Google Scholar] [CrossRef]
- Davies, G.C.; Huster, W.J.; Lu, Y.; Plouffe, L., Jr.; Lakshmanan, M. Adverse events reported by postmenopausal women in controlled trials with raloxifene. Obstet. Gynecol. 1999, 93, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Long- and short-term side effects and safety of calcitonin in man: A prospective study. Calcif. Tissue Int. 1993, 52, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, C.; Bale, H.; Gludovatz, B.; Wat, A.; Tang, S.Y.; Wang, M.; Busse, B.; Zimmermann, E.A.; Schaible, E.; Allen, M.R.; et al. Alendronate treatment alters bone tissues at multiple structural levels in healthy canine cortical bone. Bone 2015, 81, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Watts, N.B. Long-term risks of bisphosphonate therapy. Arq. Bras. Endocrinol. Metabol. 2014, 58, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Demonaco, H.J. Patient- and physician-oriented web sites and drug surveillance: Bisphosphonates and severe bone, joint, and muscle pain. Arch. Intern. Med. 2009, 169, 1164–1166. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.H.; Yoon, C.; Park, S.M. Bisphosphonate use and gastrointestinal tract cancer risk: Meta-analysis of observational studies. World J. Gastroenterol. 2012, 18, 5779–5788. [Google Scholar] [CrossRef] [PubMed]
- Andrici, J.; Tio, M.; Eslick, G.D. Meta-analysis: Oral bisphosphonates and the risk of oesophageal cancer. Aliment. Pharmacol. Ther. 2012, 36, 708–716. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence | Product Size (bp) | Accession Numbers |
|---|---|---|---|---|
| KV7.1 | Forward | CCCAAGAAGTCTGTGGTGGT | 154 | NM_000218 |
| Reverse | TGTCATAGCCGTCGACAGAG | |||
| KV7.2 | Forward | GCAAGCTGCAGAATTTCCTC | 201 | NM_004518 |
| Reverse | AGTACTCCACGCCAAACACC | |||
| KV7.3 | Forward | GGTGCAGGTCACGGAGTATT | 174 | NM_001204824 |
| Reverse | GGGCTGACTTTGTCAATGGT | |||
| KV7.4 | Forward | CTGGGCATCTCTTTCTTTGC | 160 | AH007377 |
| Reverse | GTACCAGGTGGCTGTCAGGT | |||
| KV7.5 | Forward | CGCTTTCGTTTTTCTCCTTG | 207 | NM_001160134 |
| Reverse | CGAGCAAACCTCAGTCTTCC | |||
| ALP | Forward | CCTCCTCGGAAGACACTCTG | 139 | NM_000478 |
| Reverse | GCAGTGAAGGGCTTCTTGTC | |||
| OSC | Forward | GACTGTGACGAGTTGGCTGA | 119 | NM_001199662 |
| Reverse | CTGGAGAGGAGCAGAACTGG | |||
| Runx2 | Forward | CACCGAGACCAACAGAGTCA | 95 | NM_001015051 |
| Reverse | TGATGCCATAGTCCCTCCTT | |||
| Osterix | Forward | GCCAGAAGCTGTGAAACCTC | 161 | AF477981 |
| Reverse | GCTGCAAGCTCTCCATAACC | |||
| GAPDH | Forward | CTCTGCTCCTCCTGTTCGAC | 112 | NM_002046 |
| Reverse | ACGACCAAATCCGTTGACTC |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.E.; Song, M.S.; Shen, Y.; Ryu, P.D.; Lee, S.Y. The Role of KV7.3 in Regulating Osteoblast Maturation and Mineralization. Int. J. Mol. Sci. 2016, 17, 407. https://doi.org/10.3390/ijms17030407
Yang JE, Song MS, Shen Y, Ryu PD, Lee SY. The Role of KV7.3 in Regulating Osteoblast Maturation and Mineralization. International Journal of Molecular Sciences. 2016; 17(3):407. https://doi.org/10.3390/ijms17030407
Chicago/Turabian StyleYang, Ji Eun, Min Seok Song, Yiming Shen, Pan Dong Ryu, and So Yeong Lee. 2016. "The Role of KV7.3 in Regulating Osteoblast Maturation and Mineralization" International Journal of Molecular Sciences 17, no. 3: 407. https://doi.org/10.3390/ijms17030407
APA StyleYang, J. E., Song, M. S., Shen, Y., Ryu, P. D., & Lee, S. Y. (2016). The Role of KV7.3 in Regulating Osteoblast Maturation and Mineralization. International Journal of Molecular Sciences, 17(3), 407. https://doi.org/10.3390/ijms17030407