
The Role of Galectin-3 in the Kidneys


{kind=link}
{kind=link}
Abstract
:1. Introduction
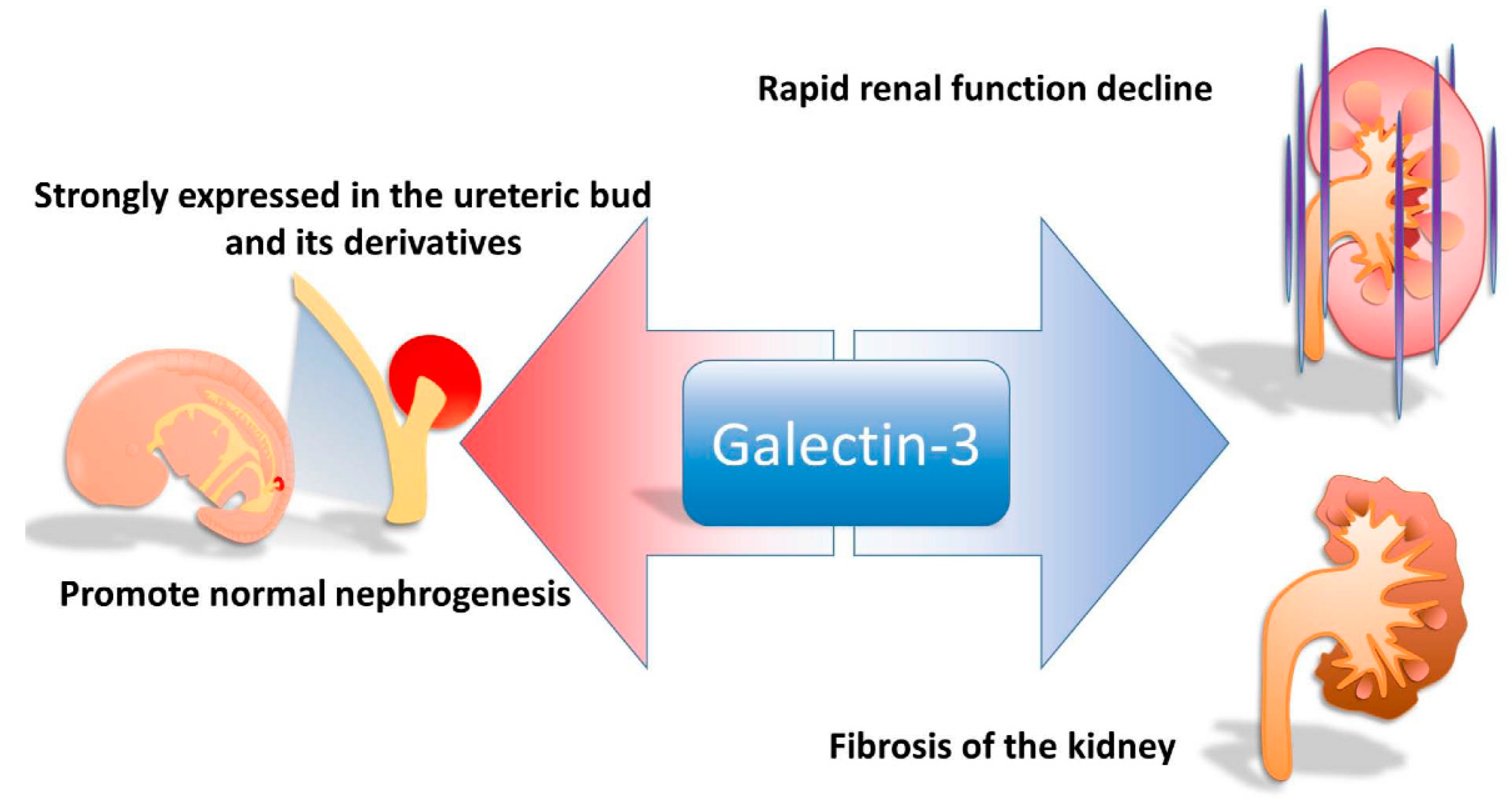
2. The Role of Galectin-3 in Nephrogenesis
3. Presentation of Galctin-3 in Animal Models of Renal Failure
4. Galctin-3 and Advanced Glycation End-Products (AGEs) in Animal Models of Diabetic Nephropathy
5. Galctin-3 Presentation in Animal Models of Ischemia/Reperfusion Injury (IRI)
6. Galectin-3 in Human Diabetic Nephropathy
7. Galectin-3 in Systemic Lupus Erythematosus (SLE) Nephritis
8. Elevated Galectin-3 in Chronic Heart Failure
9. Galectin-3 Is Associated with Renal Outcomes in Chronic Kidney Disease (CKD)
10. Galectin-3 Is Associated with Clinical Outcomes in CKD
11. Galectin-3 Is Associated with Renal Transplantation
12. Galectin-3 as a New Treatment Opportunity for Renal Disease
Acknowledgments
Conflict of Interest
References
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [PubMed]
- Cummings, R.D.; Liu, F.T. Galectins. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Kasai, K.; Hirabayashi, J. Galectins: A family of animal lectins that decipher glycocodes. J. Biochem. 1996, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Davidson, P.J.; Davis, M.J.; Patterson, R.J.; Ripoche, M.A.; Poirier, F.; Wang, J.L. Shuttling of galectin-3 between the nucleus and cytoplasm. Glycobiology 2002, 12, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.K.; Liu, F.T. Regulation of cellular homeostasis by galectins. Glycoconj. J. 2004, 19, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, J.; Furtak, V.; Lukyanov, P. Extracellular functions of galectin-3. Glycoconj. J. 2004, 19, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Dumic, J.; Dabelic, S.; Flogel, M. Galectin-3: An open-ended story. Biochim. Biophys. Acta 2006, 1760, 616–635. [Google Scholar] [CrossRef] [PubMed]
- De Boer, R.A.; Voors, A.A.; Muntendam, P.; van Gilst, W.H.; van Veldhuisen, D.J. Galectin-3: A novel mediator of heart failure development and progression. Eur. J. Heart Fail. 2009, 11, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N.C.; Sethi, T. The regulation of inflammation by galectin-3. Immunol. Rev. 2009, 230, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, M.; Fukumori, T.; Fukawa, T.; Elsamman, E.; Shiirevnyamba, A.; Nakatsuji, H.; Kanayama, H.O. Clinical significance of Galectin-3 in clear cell renal cell carcinoma. J. Med. Investig. 2010, 57, 152–157. [Google Scholar] [CrossRef]
- Straube, T.; Elli, A.F.; Greb, C.; Hegele, A.; Elsasser, H.P.; Delacour, D.; Jacob, R. Changes in the expression and subcellular distribution of galectin-3 in clear cell renal cell carcinoma. J. Exp. Clin. Cancer Res. 2011, 30, 89. [Google Scholar] [CrossRef] [PubMed]
- Menini, S.; Iacobini, C.; Blasetti Fantauzzi, C.; Pesce, C.M.; Pugliese, G. Role of galectin-3 in obesity and impaired glucose homeostasis. Oxid. Med. Cell. Longev. 2016, 2016, 9618092. [Google Scholar] [CrossRef] [PubMed]
- Dhirapong, A.; Lleo, A.; Leung, P.; Gershwin, M.E.; Liu, F.T. The immunological potential of galectin-1 and -3. Autoimmun. Rev. 2009, 8, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Feldman, C. Clinical relevance of antimicrobial resistance in the management of pneumococcal community-acquired pneumonia. J. Lab. Clin. Med. 2004, 143, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Bichara, M.; Attmane-Elakeb, A.; Brown, D.; Essig, M.; Karim, Z.; Muffat-Joly, M.; Micheli, L.; Eude-Le Parco, I.; Cluzeaud, F.; Peuchmaur, M.; et al. Exploring the role of galectin 3 in kidney function: A genetic approach. Glycobiology 2006, 16, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Winyard, P.J.; Bao, Q.; Hughes, R.C.; Woolf, A.S. Epithelial galectin-3 during human nephrogenesis and childhood cystic diseases. J. Am. Soc. Nephrol. 1997, 8, 1647–1657. [Google Scholar] [PubMed]
- Bullock, S.L.; Johnson, T.M.; Bao, Q.; Hughes, R.C.; Winyard, P.J.; Woolf, A.S. Galectin-3 modulates ureteric bud branching in organ culture of the developing mouse kidney. J. Am. Soc. Nephrol. 2001, 12, 515–523. [Google Scholar] [PubMed]
- Iacobini, C.; Menini, S.; Oddi, G.; Ricci, C.; Amadio, L.; Pricci, F.; Olivieri, A.; Sorcini, M.; di Mario, U.; Pesce, C.; Pugliese, G. Galectin-3/AGE-receptor 3 knockout mice show accelerated AGE-induced glomerular injury: Evidence for a protective role of galectin-3 as an AGE receptor. FASEB J. 2004, 18, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, J.; Kobayashi, S.; Ishida, A.; Nakabayashi, I.; Tajima, O.; Miura, S.; Katayama, M.; Nogami, H. Up-regulation of galectin-3 in acute renal failure of the rat. Am. J. Pathol. 2000, 157, 815–823. [Google Scholar] [CrossRef]
- Lohr, M.; Kaltner, H.; Lensch, M.; Andre, S.; Sinowatz, F.; Gabius, H.J. Cell-type-specific expression of murine multifunctional galectin-3 and its association with follicular atresia/luteolysis in contrast to pro-apoptotic galectins-1 and -7. Histochem. Cell Biol. 2008, 130, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.R. Macrophages and progressive renal disease in experimental hydronephrosis. Am. J. Kidney Dis. 1995, 26, 133–140. [Google Scholar] [CrossRef]
- Henderson, N.C.; Mackinnon, A.C.; Farnworth, S.L.; Kipari, T.; Haslett, C.; Iredale, J.P.; Liu, F.T.; Hughes, J.; Sethi, T. Galectin-3 expression and secretion links macrophages to the promotion of renal fibrosis. Am. J. Pathol. 2008, 172, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Okamura, D.M.; Pasichnyk, K.; Lopez-Guisa, J.M.; Collins, S.; Hsu, D.K.; Liu, F.T.; Eddy, A.A. Galectin-3 preserves renal tubules and modulates extracellular matrix remodeling in progressive fibrosis. Am. J. Physiol. Ren. Physiol. 2011, 300, F245–F253. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, G.; Pricci, F.; Iacobini, C.; Leto, G.; Amadio, L.; Barsotti, P.; Frigeri, L.; Hsu, D.K.; Vlassara, H.; Liu, F.T.; et al. Accelerated diabetic glomerulopathy in galectin-3/AGE receptor 3 knockout mice. FASEB J. 2001, 15, 2471–2479. [Google Scholar] [CrossRef] [PubMed]
- Vansthertem, D.; Cludts, S.; Nonclercq, D.; Gossiaux, A.; Saussez, S.; Legrand, A.; Gabius, H.J.; Toubeau, G. Immunohistochemical localization of galectins-1 and -3 and monitoring of tissue galectin-binding sites during tubular regeneration after renal ischemia reperfusion in the rat. Histol. Histopathol. 2010, 25, 1417–1429. [Google Scholar] [PubMed]
- Fernandes Bertocchi, A.P.; Campanhole, G.; Wang, P.H.; Goncalves, G.M.; Damiao, M.J.; Cenedeze, M.A.; Beraldo, F.C.; de Paula Antunes Teixeira, V.; dos Reis, M.A.; Mazzali, M.; et al. A Role for galectin-3 in renal tissue damage triggered by ischemia and reperfusion injury. Transpl. Int. 2008, 21, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Kobayashi, S.; Hemmi, N.; Ikee, R.; Hyodo, N.; Saigusa, T.; Namikoshi, T.; Yamada, M.; Suzuki, S.; Miura, S. Galectin-3-positive cell infiltration in human diabetic nephropathy. Nephrol. Dial. Transplant. 2004, 19, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.H.; Moon, K.C.; Lee, E.Y.; Lee, Y.J.; Lee, E.B.; Ahn, C.; Song, Y.W. Renal expression of galectin-3 in systemic lupus erythematosus patients with nephritis. Lupus 2009, 18, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.C.; Pokharel, S.; van Brakel, T.J.; van Berlo, J.H.; Cleutjens, J.P.; Schroen, B.; Andre, S.; Crijns, H.J.; Gabius, H.J.; Maessen, J.; et al. Galectin-3 marks activated macrophages in failure-prone hypertrophied hearts and contributes to cardiac dysfunction. Circulation 2004, 110, 3121–3128. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Ruifrok, W.P.; Meissner, M.; Bos, E.M.; van Goor, H.; Sanjabi, B.; van der Harst, P.; Pitt, B.; Goldstein, I.J.; Koerts, J.A.; et al. Genetic and pharmacological inhibition of galectin-3 prevents cardiac remodeling by interfering with myocardial fibrogenesis. Circ. Heart Fail. 2013, 6, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Schroen, B.; Heymans, S.; Sharma, U.; Blankesteijn, W.M.; Pokharel, S.; Cleutjens, J.P.; Porter, J.G.; Evelo, C.T.; Duisters, R.; van Leeuwen, R.E.; et al. Thrombospondin-2 is essential for myocardial matrix integrity: Increased expression identifies failure-prone cardiac hypertrophy. Circ. Res. 2004, 95, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Miana, M.; Reboul, P.; Cachofeiro, V.; Martinez-Martinez, E.; de Boer, R.A.; Poirier, F.; Lacolley, P.; Zannad, F.; Rossignol, P.; et al. Galectin-3 mediates aldosterone-induced vascular fibrosis. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Shrestha, K.; Shao, Z.; Borowski, A.G.; Troughton, R.W.; Thomas, J.D.; Klein, A.L. Usefulness of plasma galectin-3 levels in systolic heart failure to predict renal insufficiency and survival. Am. J. Cardiol. 2011, 108, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Lok, D.J.; van der Meer, P.; de la Porte, P.W.; Lipsic, E.; van Wijngaarden, J.; Hillege, H.L.; van Veldhuisen, D.J. Prognostic value of galectin-3, a novel marker of fibrosis, in patients with chronic heart failure: Data from the DEAL-HF study. Clin. Res. Cardiol. 2010, 99, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Zamora, E.; Lupon, J.; de Antonio, M.; Galan, A.; Domingo, M.; Urrutia, A.; Troya, M.; Bayes-Genis, A. Renal function largely influences Galectin-3 prognostic value in heart failure. Int. J. Cardiol. 2014, 177, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Gopal, D.M.; Kommineni, M.; Ayalon, N.; Koelbl, C.; Ayalon, R.; Biolo, A.; Dember, L.M.; Downing, J.; Siwik, D.A.; Liang, C.S.; et al. Relationship of plasma galectin-3 to renal function in patients with heart failure: Effects of clinical status, pathophysiology of heart failure, and presence or absence of heart failure. J. Am. Heart Assoc. 2012, 1, e000760. [Google Scholar] [CrossRef] [PubMed]
- Meijers, W.C.; van der Velde, A.R.; Ruifrok, W.P.; Schroten, N.F.; Dokter, M.M.; Damman, K.; Assa, S.; Franssen, C.F.; Gansevoort, R.T.; van Gilst, W.H.; et al. Renal handling of galectin-3 in the general population, chronic heart failure, and hemodialysis. J. Am. Heart Assoc. 2014, 3, e000962. [Google Scholar] [CrossRef] [PubMed]
- Ensrud, K.E.; Lui, L.Y.; Taylor, B.C.; Ishani, A.; Shlipak, M.G.; Stone, K.L.; Cauley, J.A.; Jamal, S.A.; Antoniucci, D.M.; Cummings, S.R. Renal function and risk of hip and vertebral fractures in older women. Arch. Intern. Med. 2007, 167, 133–139. [Google Scholar] [CrossRef] [PubMed]
- McClellan, W.M.; Flanders, W.D. Risk factors for progressive chronic kidney disease. J. Am. Soc. Nephrol. 2003, 14, S65–S70. [Google Scholar] [CrossRef] [PubMed]
- Perkovic, V.; Cass, A.; Patel, A.A.; Suriyawongpaisal, P.; Barzi, F.; Chadban, S.; Macmahon, S.; Neal, B. High prevalence of chronic kidney disease in Thailand. Kidney Int. 2008, 73, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Shiohira, Y.; Uezu, Y.; Higa, A.; Iseki, K. Metabolic syndrome and chronic kidney disease in Okinawa, Japan. Kidney. Int. 2006, 69, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Weiner, D.E.; Tighiouart, H.; Amin, M.G.; Stark, P.C.; MacLeod, B.; Griffith, J.L.; Salem, D.N.; Levey, A.S.; Sarnak, M.J. Chronic kidney disease as a risk factor for cardiovascular disease and all-cause mortality: A pooled analysis of community-based studies. J. Am. Soc. Nephrol. 2004, 15, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, D.; Ramjit, D.; Zhang, Z.; Ribeiro, A.B.; Kurokawa, K.; Lash, J.P.; Chan, J.; Remuzzi, G.; Brenner, B.M.; Shahinfar, S. Renal risk and renoprotection among ethnic groups with type 2 diabetic nephropathy: A post hoc analysis of RENAAL. Kidney Int. 2006, 69, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- O'Seaghdha, C.M.; Hwang, S.J.; Ho, J.E.; Vasan, R.S.; Levy, D.; Fox, C.S. Elevated galectin-3 precedes the development of CKD. J. Am. Soc. Nephrol. 2013, 24, 1470–1477. [Google Scholar] [CrossRef] [PubMed]
- Iacobini, C.; Amadio, L.; Oddi, G.; Ricci, C.; Barsotti, P.; Missori, S.; Sorcini, M.; di Mario, U.; Pricci, F.; Pugliese, G. Role of galectin-3 in diabetic nephropathy. J. Am. Soc. Nephrol. 2003, 14, S264–S270. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, C.; Delgado, G.; Wanner, C.; Blouin, K.; Pilz, S.; Tomaschitz, A.; Kleber, M.E.; Dressel, A.; Willmes, C.; Krane, V.; et al. Galectin-3, renal function, and clinical outcomes: Results from the LURIC and 4D studies. J. Am. Soc. Nephrol. 2015, 26, 2213–2221. [Google Scholar] [CrossRef] [PubMed]
- De Boer, R.A.; Lok, D.J.; Jaarsma, T.; van der Meer, P.; Voors, A.A.; Hillege, H.L.; van Veldhuisen, D.J. Predictive value of plasma galectin-3 levels in heart failure with reduced and preserved ejection fraction. Ann. Med. 2011, 43, 60–68. [Google Scholar] [CrossRef] [PubMed]
- De Boer, R.A.; van Veldhuisen, D.J.; Gansevoort, R.T.; Muller Kobold, A.C.; van Gilst, W.H.; Hillege, H.L.; Bakker, S.J.; van der Harst, P. The fibrosis marker galectin-3 and outcome in the general population. J. Intern. Med. 2012, 272, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Arend, S.M.; Mallat, M.J.; Westendorp, R.J.; van der Woude, F.J.; van Es, L.A. Patient survival after renal transplantation; more than 25 years follow-up. Nephrol. Dial. Transplant. 1997, 12, 1672–1679. [Google Scholar] [PubMed]
- Tan, R.; Liu, X.; Wang, J.; Lu, P.; Han, Z.; Tao, J.; Yin, C.; Gu, M. Alternations of galectin levels after renal transplantation. Clin. Biochem. 2014, 47, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Dang, Z.; MacKinnon, A.; Marson, L.P.; Sethi, T. Tubular atrophy and interstitial fibrosis after renal transplantation is dependent on galectin-3. Transplantation 2012, 93, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Bao, Q.; Hughes, R.C. Galectin-3 modulates rat mesangial cell proliferation and matrix synthesis during experimental glomerulonephritis induced by anti-Thy1.1 antibodies. J. Pathol. 1999, 187, 481–489. [Google Scholar] [CrossRef]
- Sun, Z.L.; Ma, C.J.; Jin, H.; Yuan, Y.; Liu, N.F. Effects of advanced glycosylation end products and rosiglitazone on the expression and secretion of galectin-3 in human renal mesangial cells. Chin. Med. J. 2009, 122, 1067–1071. [Google Scholar] [PubMed]
- Kolatsi-Joannou, M.; Price, K.L.; Winyard, P.J.; Long, D.A. Modified citrus pectin reduces galectin-3 expression and disease severity in experimental acute kidney injury. PLoS ONE 2011, 6, e18683. [Google Scholar] [CrossRef] [PubMed]
- La Jolla begins Phase IIb trial of GCS-100 to treat advanced CKD. Available online: http://www.drugdevelopment-technology.com/news/newsla-jolla-begins-phase-iib-trial-of-gcs-100-to-treat-advanced-ckd-4535972 (accessed on 12 April 2016).


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-C.; Kuo, P.-L. The Role of Galectin-3 in the Kidneys. Int. J. Mol. Sci. 2016, 17, 565. https://doi.org/10.3390/ijms17040565
Chen S-C, Kuo P-L. The Role of Galectin-3 in the Kidneys. International Journal of Molecular Sciences. 2016; 17(4):565. https://doi.org/10.3390/ijms17040565
Chicago/Turabian StyleChen, Szu-Chia, and Po-Lin Kuo. 2016. "The Role of Galectin-3 in the Kidneys" International Journal of Molecular Sciences 17, no. 4: 565. https://doi.org/10.3390/ijms17040565