
Cypermethrin Induces Macrophages Death through Cell Cycle Arrest and Oxidative Stress-Mediated JNK/ERK Signaling Regulated Apoptosis

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
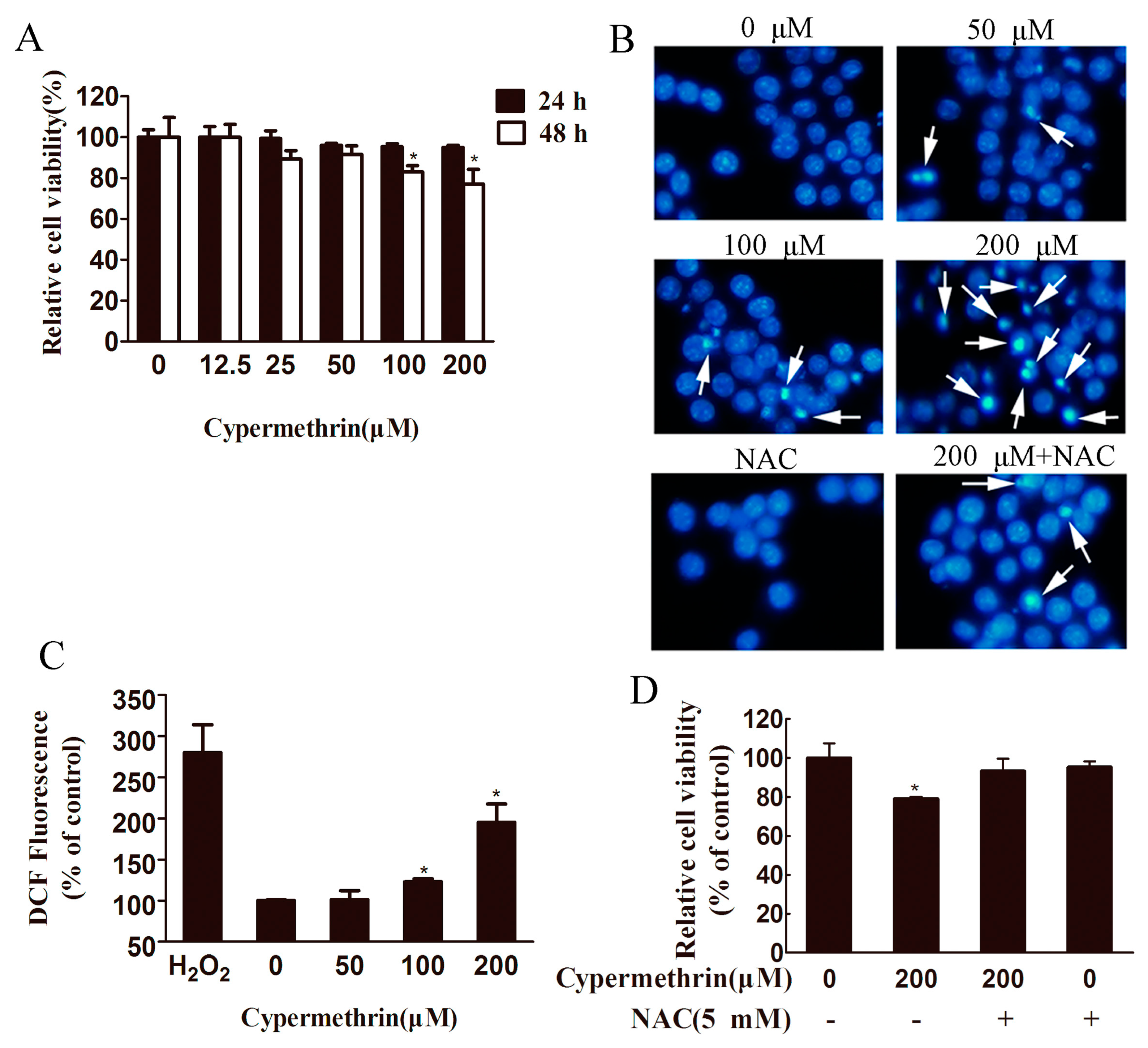
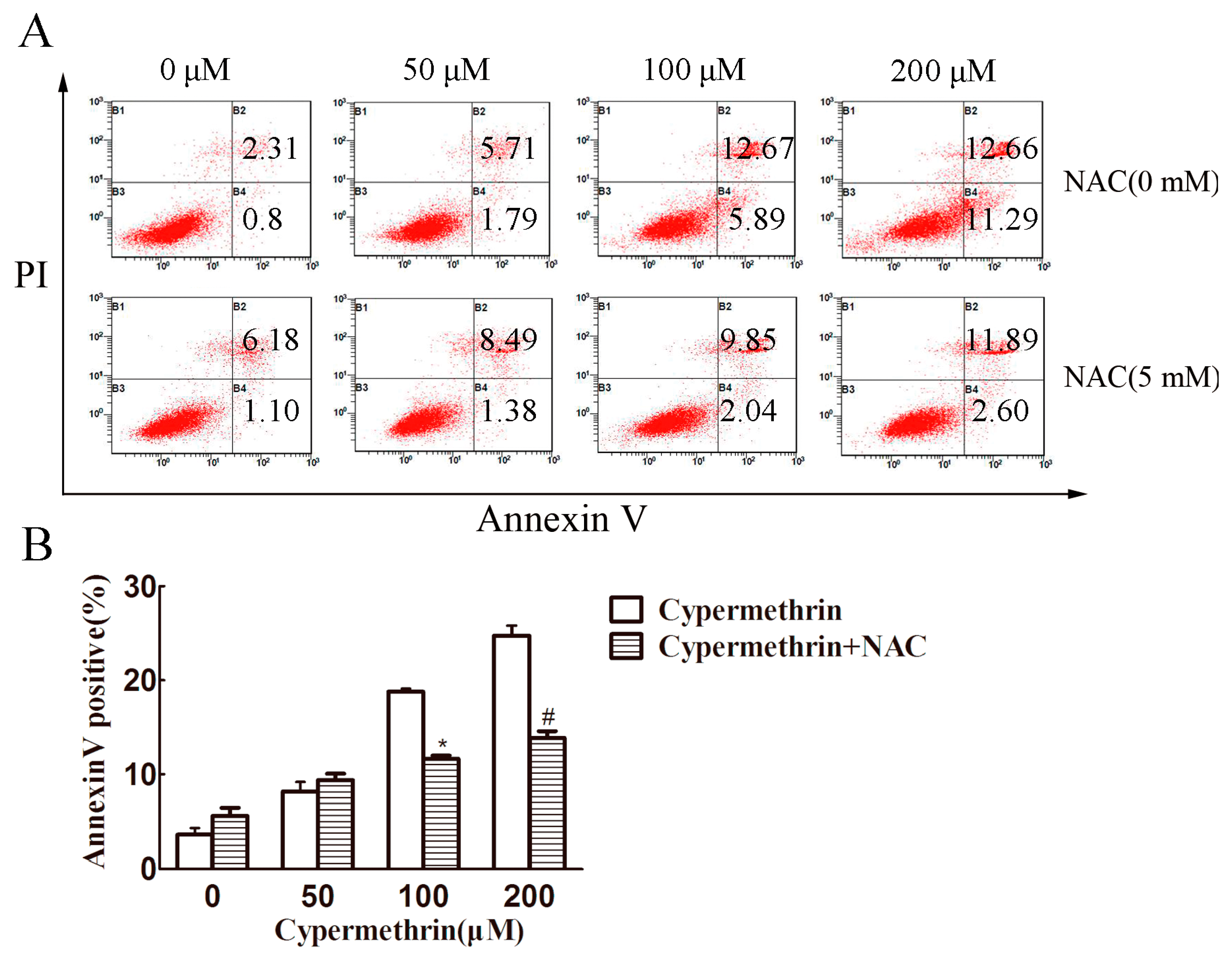
2.1. Cytotoxic Effect of Cypermethrin, Reactive Oxygen Species (ROS) Production and Apoptosis Induction in RAW 264.7 Cells
2.2. Induction of G1 Cell Cycle Arrest in Raw 264.7 Cells by Cypermethrin
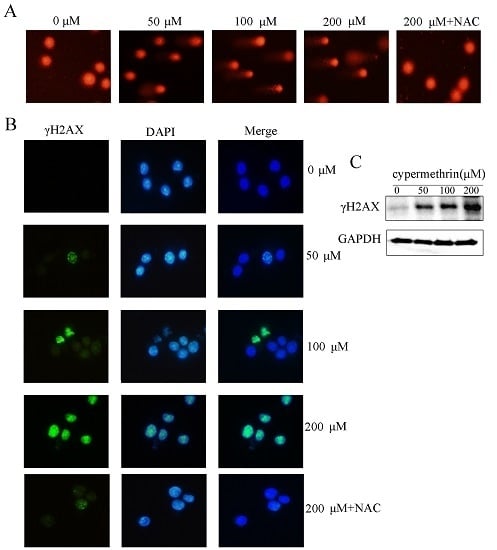
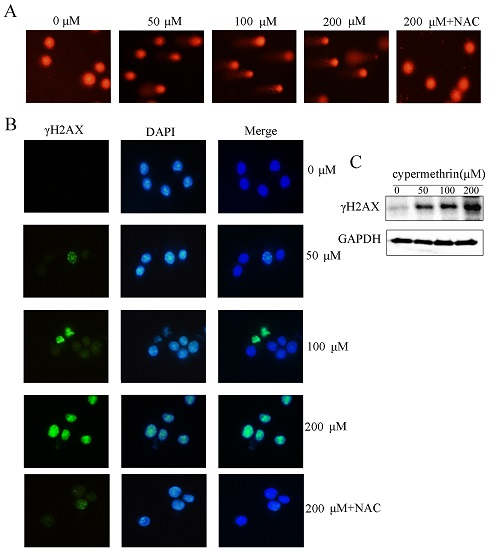
2.3. Cypermethrin-Induced ROS Generation Mediated RAW Cell Apoptosis via Causing DNA Damage
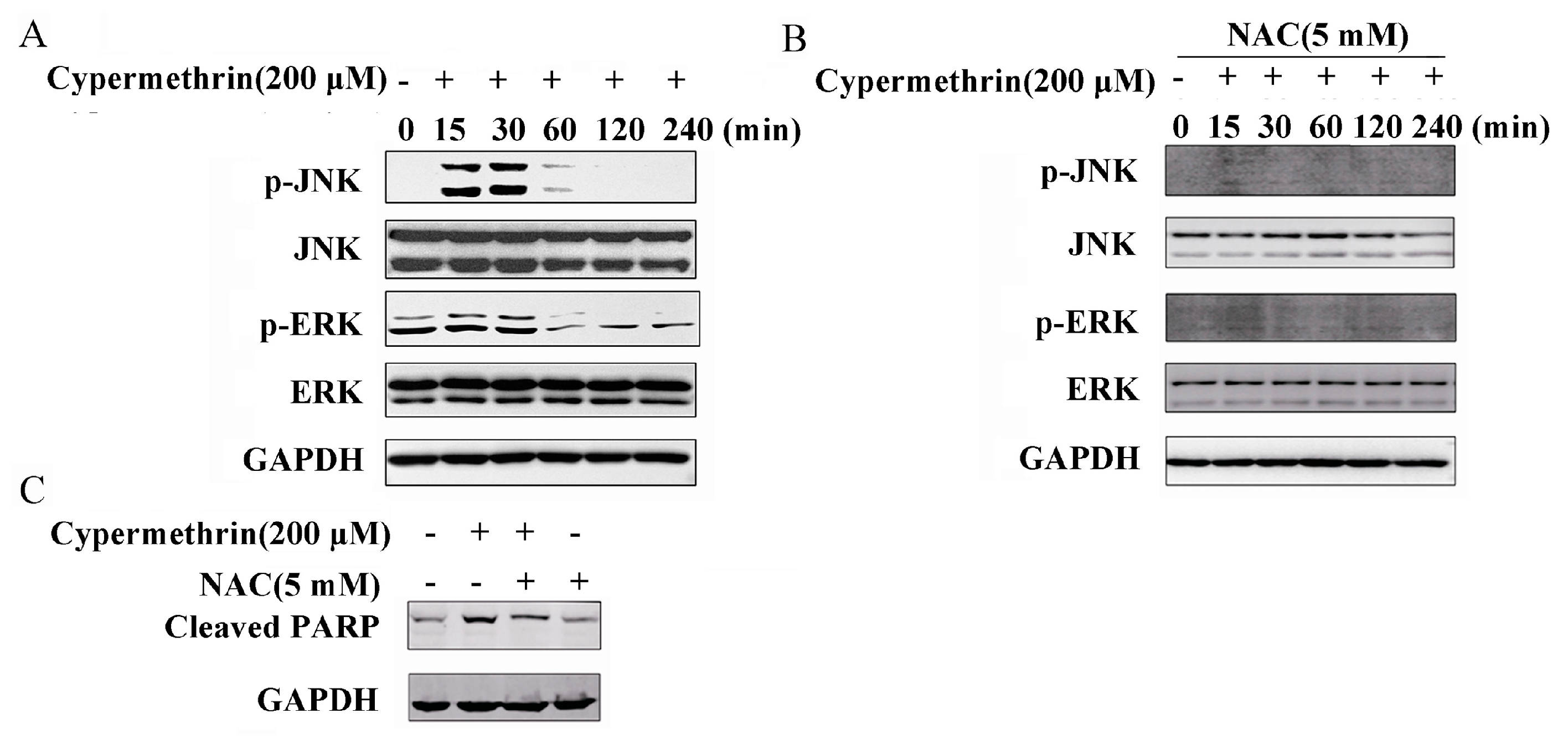
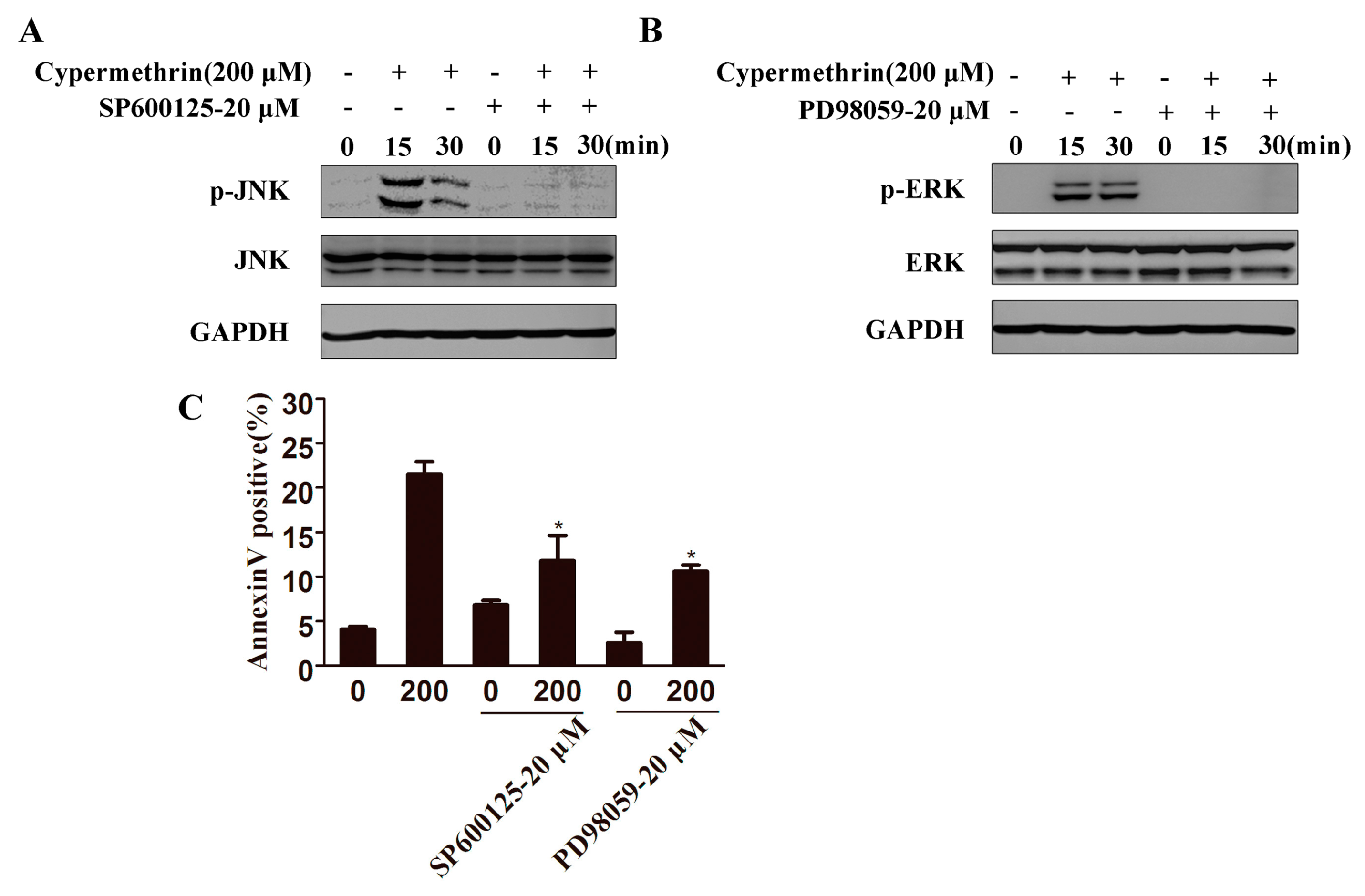
2.4. MAPK Signaling Pathway Involved in Cypermethrin-Induced RAW Cell Apoptosis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Chemicals and Antibodies
4.3. Cell Viability Assay
4.4. Cell Cycle Analysis
4.5. Analysis of Apoptosis
4.6. Comet Assay
4.7. Immunofluorescence Microscopy
4.8. Measurement of ROS Production
4.9. Immunoblotting
4.10. Hoechst 33342 Staining
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nasuti, C.; Gabbianelli, R.; Falcioni, M.L.; di Stefano, A.; Sozio, P.; Cantalamessa, F. Dopaminergic system modulation, behavioral changes, and oxidative stress after neonatal administration of pyrethroids. Toxicology 2007, 229, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Giordano, G.; Guizzetti, M.; Vitalone, A. Neurotoxicity of pesticides: A brief review. Front. Biosci. 2008, 13, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Heudorf, U.; Angerer, J.; Drexler, H. Current internal exposure to pesticides in children and adolescents in Germany: Urinary levels of metabolites of pyrethroid and organophosphorus insecticides. Int. Arch. Occup. Environ. Health 2004, 77, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Carriquiriborde, P.; Diaz, J.; Mugni, H.; Bonetto, C.; Ronco, A.E. Impact of cypermethrin on stream fish populations under field-use in biotech-soybean production. Chemosphere 2007, 68, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Jaensson, A.; Scott, A.P.; Moore, A.; Kylin, H.; Olsen, K.H. Effects of a pyrethroid pesticide on endocrine responses to female odours and reproductive behavior in male parr of brown trout (Salmo trutta L.). Aquat. Toxicol. 2007, 81, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sepici-Dincel, A.; Caglan Karasu Benli, A.; Selvi, M.; Sarikaya, R.; Sahin, D.; Ayhan Ozkul, I.; Erkoc, F. Sublethal cyfluthrin toxicity to carp (Cyprinus carpio L.) fingerlings: Biochemical, hematological, histopathological alterations. Ecotoxicol. Environ. Saf. 2009, 72, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Kan, Y.; Cengiz, E.I.; Ugurlu, P.; Yanar, M. The protective role of vitamin E on gill and liver tissue histopathology and micronucleus frequencies in peripheral erythrocytes of Oreochromis niloticus exposed to deltamethrin. Environ. Toxicol. Pharmacol. 2012, 34, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Lund, A.E.; Narahashi, T. Modification of sodium channel kinetics by the insecticide tetramethrin in crayfish giant axons. Neurotoxicology 1981, 2, 213–229. [Google Scholar] [PubMed]
- Prashanth, M.S.; David, M. Impact of cypermethrin on Na+-K+, Ca2+ and Mg2+ ATPases in Indian major carp, Cirrhinus mrigala (Hamilton). Bull. Environ. Contam. Toxicol. 2010, 84, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Agarwal, A.K.; Shukla, G.S. Effect of quinalphos and cypermethrin exposure on developing blood-brain barrier: Role of nitric oxide. Environ. Toxicol. Pharmacol. 2000, 8, 73–78. [Google Scholar] [CrossRef]
- Li, Y.F.; Pan, C.; Hu, J.X.; Li, J.; Xu, L.C. Effects of cypermethrin on male reproductive system in adult rats. Biomed. Environ. Sci. 2013, 26, 201–208. [Google Scholar] [PubMed]
- Liu, L.; Hu, J.X.; Wang, H.; Chen, B.J.; He, Z.; Xu, L.C. Effects of β-cypermethrin on male rat reproductive system. Environ. Toxicol. Pharmacol. 2010, 30, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Maurya, S.K.; Mishra, J.; Tripathi, V.K.; Sharma, R.; Siddiqui, M.H. Cypermethrin induces astrocyte damage: Role of aberrant Ca2+, ROS, JNK, P38, matrix metalloproteinase 2 and migration related reelin protein. Pestic. Biochem. Physiol. 2014, 111, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.Y.; Lee, W.Y.; Han, S.S. Effects of cypermethrin on the dopaminergic neurons in the progressive hemiparkinsonian rats. Toxicol. Mech. Methods 2005, 15, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zheng, S.; Pu, Y.; Shu, L.; Sun, L.; Liu, W.; Fu, Z. Cypermethrin has the potential to induce hepatic oxidative stress, DNA damage and apoptosis in adult zebrafish (Danio rerio). Chemosphere 2011, 82, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Tang, S.; Mayer, G.D.; Cobb, G.P.; Maul, J.D. Interactive effects of ultraviolet-B radiation and pesticide exposure on DNA photo-adduct accumulation and expression of DNA damage and repair genes in Xenopus laevis embryos. Aquat. Toxicol. 2015, 159, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Srivastava, A.K.; Singh, A.K. Cell cycle stage specific application of cypermethrin and carbendazim to assess the genotoxicity in somatic cells of Hordeum vulgare L. Bull. Environ. Contam. Toxicol. 2008, 81, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Kastan, M.B.; Canman, C.E.; Leonard, C.J. P53, cell cycle control and apoptosis: Implications for cancer. Cancer Metastasis Rev. 1995, 14, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Capetillo, O.; Lee, A.; Nussenzweig, M.; Nussenzweig, A. H2AX: The histone guardian of the genome. DNA Repair 2004, 3, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Redon, C.; Pilch, D.; Rogakou, E.; Sedelnikova, O.; Newrock, K.; Bonner, W. Histone H2A variants H2AX and H2AZ. Curr. Opin. Genet. Dev. 2002, 12, 162–169. [Google Scholar] [CrossRef]
- Cowan, K.J.; Storey, K.B. Mitogen-activated protein kinases: New signaling pathways functioning in cellular responses to environmental stress. J. Exp. Biol. 2003, 206 Pt 7, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Hussell, T.; Bell, T.J. Alveolar macrophages: Plasticity in a tissue-specific context. Nat. Rev. Immunol. 2014, 14, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Zhang, Q.; Fu, J.; Woods, C.G.; Hou, Y.; Corkey, B.E.; Collins, S.; Andersen, M.E. ROS signaling, oxidative stress and Nrf2 in pancreatic beta-cell function. Toxicol. Appl. Pharmacol. 2010, 244, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Liu, Y.; Liu, H.; Yu, H.; Wang, H.; Xia, Z. Effects of N-acetylcysteine on nicotinamide dinucleotide phosphate oxidase activation and antioxidant status in heart, lung, liver and kidney in streptozotocin-induced diabetic rats. Yonsei Med. J. 2012, 53, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Kuo, K.H.; Nagareddy, P.R.; Wang, F.; Guo, Z.; Guo, T.; Jiang, J.; McNeill, J.H. N-acetylcysteine attenuates PKCβ2 overexpression and myocardial hypertrophy in streptozotocin-induced diabetic rats. Cardiovasc. Res. 2007, 73, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Fiordaliso, F.; Bianchi, R.; Staszewsky, L.; Cuccovillo, I.; Doni, M.; Laragione, T.; Salio, M.; Savino, C.; Melucci, S.; Santangelo, F.; et al. Antioxidant treatment attenuates hyperglycemia-induced cardiomyocyte death in rats. J. Mol. Cell. Cardiol. 2004, 37, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.M.; Bae, Y.S.; Lee, S.Y. Molecular ordering of ROS production, mitochondrial changes, and caspase activation during sodium salicylate-induced apoptosis. Free Radic. Biol. Med. 2003, 34, 434–442. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R. Reactive oxygen species and death: oxidative DNA damage in atherosclerosis. Circ. Res. 2001, 88, 648–650. [Google Scholar] [CrossRef] [PubMed]
- McKelvey-Martin, V.J.; Green, M.H.; Schmezer, P.; Pool-Zobel, B.L.; de Meo, M.P.; Collins, A. The single cell gel electrophoresis assay (comet assay): A European review. Mutat. Res. 1993, 288, 47–63. [Google Scholar] [CrossRef]
- Fernandez-Capetillo, O.; Celeste, A.; Nussenzweig, A. Focusing on foci: H2AX and the recruitment of DNA-damage response factors. Cell Cycle 2003, 2, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Capetillo, O.; Chen, H.T.; Celeste, A.; Ward, I.; Romanienko, P.J.; Morales, J.C.; Naka, K.; Xia, Z.; Camerini-Otero, R.D.; Motoyama, N.; et al. DNA damage-induced G2-M checkpoint activation by histone H2AX and 53BP1. Nat. Cell Biol. 2002, 4, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, T.; Huang, P.; Zhao, H.; Zhang, R.; Ma, B.; Chen, K.; Huang, F.; Zhou, X.; Cui, C.; et al. A novel Golgi protein (GOLPH2)-regulated oncolytic adenovirus exhibits potent antitumor efficacy in hepatocellular carcinoma. Oncotarget 2015, 6, 13564–13578. [Google Scholar] [CrossRef] [PubMed]
- El-Deiry, W.S.; Tokino, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar] [CrossRef]
- Xia, M.; Knezevic, D.; Vassilev, L.T. p21 does not protect cancer cells from apoptosis induced by nongenotoxic p53 activation. Oncogene 2011, 30, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Antony, M.L.; Kim, S.H.; Singh, S.V. Critical role of p53 upregulated modulator of apoptosis in benzyl isothiocyanate-induced apoptotic cell death. PLoS ONE 2012, 7, e32267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vousden, K.H.; Lu, X. Live or let die: the cell’s response to p53. Nat. Rev. Cancer 2002, 2, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Boutros, T.; Chevet, E.; Metrakos, P. Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase regulation: Roles in cell growth, death, and cancer. Pharmacol. Rev. 2008, 60, 261–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Chen, S.J.; Su, C.C.; Yen, C.C.; Tseng, T.J.; Jinn, T.R.; Tang, F.C.; Chen, K.L.; Su, Y.C.; Lee, K.I.; et al. Chloroacetic acid induced neuronal cells death through oxidative stress-mediated p38-MAPK activation pathway regulated mitochondria-dependent apoptotic signals. Toxicology 2013, 303, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.M.; Liu, Z.G. JNK signaling pathway is a key modulator in cell death mediated by reactive oxygen and nitrogen species. Free Radic. Biol. Med. 2006, 40, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, F.; Liu, Q.; Xie, S.; Xu, J.; Huang, B.; Wu, Y.; Xia, D. Cypermethrin Induces Macrophages Death through Cell Cycle Arrest and Oxidative Stress-Mediated JNK/ERK Signaling Regulated Apoptosis. Int. J. Mol. Sci. 2016, 17, 885. https://doi.org/10.3390/ijms17060885
Huang F, Liu Q, Xie S, Xu J, Huang B, Wu Y, Xia D. Cypermethrin Induces Macrophages Death through Cell Cycle Arrest and Oxidative Stress-Mediated JNK/ERK Signaling Regulated Apoptosis. International Journal of Molecular Sciences. 2016; 17(6):885. https://doi.org/10.3390/ijms17060885
Chicago/Turabian StyleHuang, Fang, Qiaoyun Liu, Shujun Xie, Jian Xu, Bo Huang, Yihua Wu, and Dajing Xia. 2016. "Cypermethrin Induces Macrophages Death through Cell Cycle Arrest and Oxidative Stress-Mediated JNK/ERK Signaling Regulated Apoptosis" International Journal of Molecular Sciences 17, no. 6: 885. https://doi.org/10.3390/ijms17060885
APA StyleHuang, F., Liu, Q., Xie, S., Xu, J., Huang, B., Wu, Y., & Xia, D. (2016). Cypermethrin Induces Macrophages Death through Cell Cycle Arrest and Oxidative Stress-Mediated JNK/ERK Signaling Regulated Apoptosis. International Journal of Molecular Sciences, 17(6), 885. https://doi.org/10.3390/ijms17060885