
Ectopic Expression of Testis Germ Cell Proteins in Cancer and Its Potential Role in Genomic Instability


Abstract
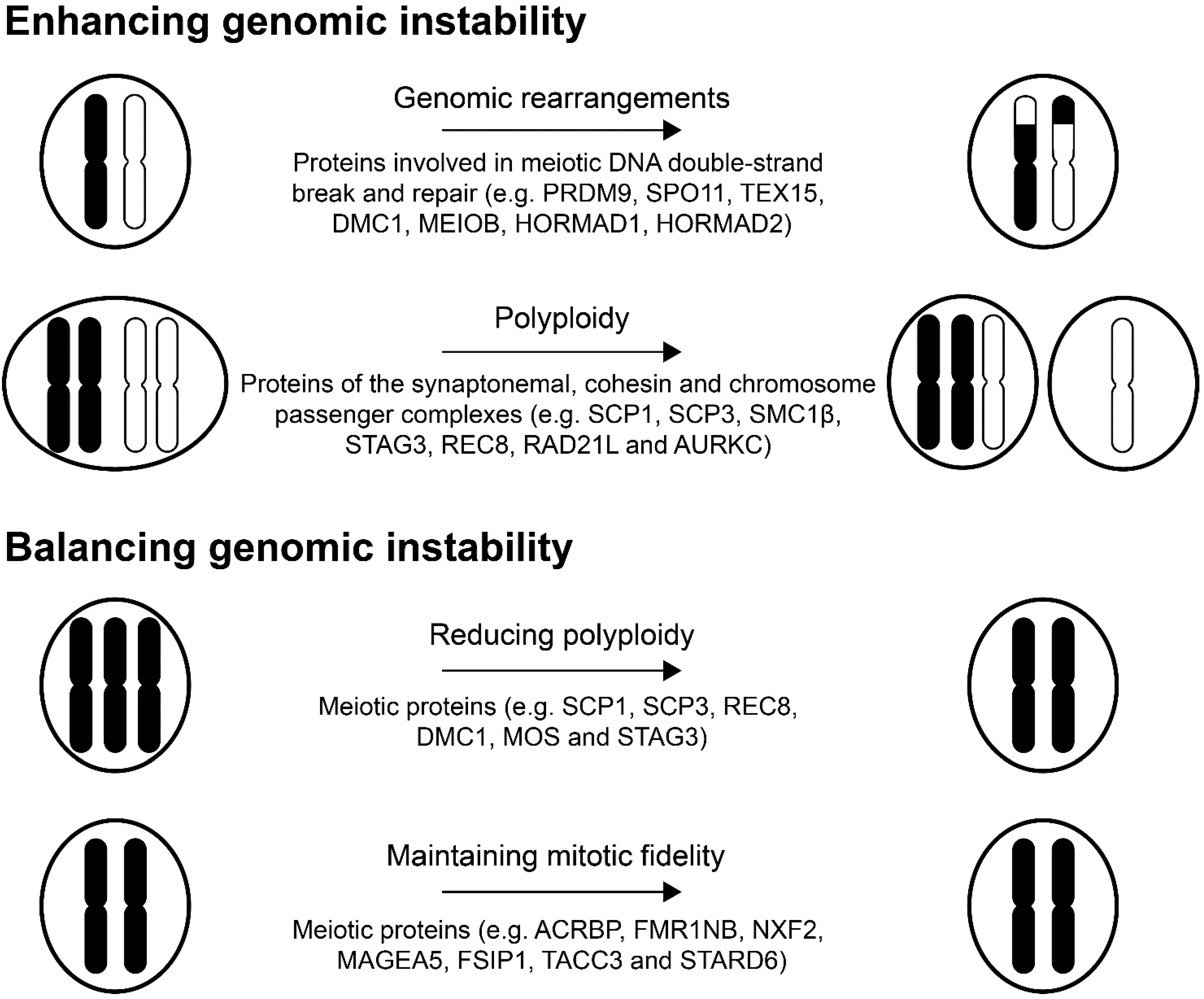
:1. Introduction
2. The Role of Meiotic Proteins in Tumorigenesis
2.1. Proteins Involved in Meiotic Recombination
2.2. Proteins Associated with the Synaptonemal Complex
2.3. Aurora Kinase C and the Chromosome Passenger Complex
3. Testis Germ Cell Proteins Regulate Polyploidy and Maintain Mitotic Fidelity
4. Oncogenic Testis Proteins May Promote Replication Stress
5. Conclusions and Future Perspectives
Author Contributions
Conflicts of Interest
References
- Shen, Z. Genomic instability and cancer: An introduction. J. Mol. Cell. Biol. 2011, 3, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.; Medema, R.H. Genetic instability: Tipping the balance. Oncogene 2013, 32, 4459–4470. [Google Scholar] [CrossRef] [PubMed]
- Hollstein, M.; Sidransky, D.; Vogelstein, B.; Harris, C.C. p53 mutations in human cancers. Science 1991, 253, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Lynch, T.J.; Bell, D.W.; Sordella, R.; Gurubhagavatula, S.; Okimoto, R.A.; Brannigan, B.W.; Harris, P.L.; Haserlat, S.M.; Supko, J.G.; Haluska, F.G.; et al. Activating mutations in the epidermal growth factor receptor underlying responsiveness of non-small-cell lung cancer to gefitinib. N. Engl. J. Med. 2004, 350, 2129–2139. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Dacic, S.; Flanagan, M.; Cieply, K.; Ramalingam, S.; Luketich, J.; Belani, C.; Yousem, S.A. Significance of EGFR protein expression and gene amplification in non-small cell lung carcinoma. Am. J. Clin. Pathol. 2006, 125, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Salesse, S.; Verfaillie, C.M. BCR/ABL: From molecular mechanisms of leukemia induction to treatment of chronic myelogenous leukemia. Oncogene 2002, 21, 8547–8559. [Google Scholar] [CrossRef] [PubMed]
- Ladanyi, M. Fusions of the SYT and SSX genes in synovial sarcoma. Oncogene 2001, 20, 5755–5762. [Google Scholar] [CrossRef] [PubMed]
- Hnisz, D.; Weintraub, A.S.; Day, D.S.; Valton, A.L.; Bak, R.O.; Li, C.H.; Goldmann, J.; Lajoie, B.R.; Fan, Z.P.; Sigova, A.A.; et al. Activation of proto-oncogenes by disruption of chromosome neighborhoods. Science 2016, 351, 1454–1458. [Google Scholar] [CrossRef] [PubMed]
- Cleaver, J.E. Cancer in xeroderma pigmentosum and related disorders of DNA repair. Nat. Rev. Cancer 2005, 5, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Bachrati, C.Z.; Hickson, I.D. RecQ helicases: Suppressors of tumorigenesis and premature aging. Biochem. J. 2003, 374, 577–606. [Google Scholar] [CrossRef] [PubMed]
- Ripperger, T.; Gadzicki, D.; Meindl, A.; Schlegelberger, B. Breast cancer susceptibility: Current knowledge and implications for genetic counselling. Eur. J. Hum. Genet. 2009, 17, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.D.; Parsons, D.W.; Jones, S.; Lin, J.; Sjoblom, T.; Leary, R.J.; Shen, D.; Boca, S.M.; Barber, T.; Ptak, J.; et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science 2007, 318, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Zhang, X.; Parsons, D.W.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Kamiyama, H.; Jimeno, A.; et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science 2008, 321, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An integrated genomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Getz, G.; Wheeler, D.A.; Mardis, E.R.; McLellan, M.D.; Cibulskis, K.; Sougnez, C.; Greulich, H.; Muzny, D.M.; Morgan, M.B.; et al. Somatic mutations affect key pathways in lung adenocarcinoma. Nature 2008, 455, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Macheret, M.; Halazonetis, T.D. DNA replication stress as a hallmark of cancer. Annu. Rev. Pathol. 2015, 10, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.J.; Caballero, O.L.; Jungbluth, A.; Chen, Y.T.; Old, L.J. Cancer/testis antigens, gametogenesis and cancer. Nat. Rev. Cancer 2005, 5, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Gjerstorff, M.F.; Andersen, M.H.; Ditzel, H.J. Oncogenic cancer/testis antigens: Prime candidates for immunotherapy. Oncotarget 2015, 6, 15772–15787. [Google Scholar] [CrossRef] [PubMed]
- Sigalotti, L.; Coral, S.; Nardi, G.; Spessotto, A.; Cortini, E.; Cattarossi, I.; Colizzi, F.; Altomonte, M.; Maio, M. Promoter methylation controls the expression of MAGE2, 3 and 4 genes in human cutaneous melanoma. J. Immunother. 2002, 25, 16–26. [Google Scholar] [CrossRef] [PubMed]
- De Smet, C.; de Backer, O.; Faraoni, I.; Lurquin, C.; Brasseur, F.; Boon, T. The activation of human gene MAGE-1 in tumor cells is correlated with genome-wide demethylation. Proc. Natl. Acad. Sci. USA 1996, 93, 7149–7153. [Google Scholar] [CrossRef] [PubMed]
- De Smet, C.; Loriot, A.; Boon, T. Promoter-dependent mechanism leading to selective hypomethylation within the 5′ region of gene MAGE-A1 in tumor cells. Mol. Cell. Biol. 2004, 24, 4781–4790. [Google Scholar] [CrossRef] [PubMed]
- Loriot, A.; de Plaen, E.; Boon, T.; de Smet, C. Transient down-regulation of DNMT1 methyltransferase leads to activation and stable hypomethylation of MAGE-A1 in melanoma cells. J. Biol. Chem. 2006, 281, 10118–10126. [Google Scholar] [CrossRef] [PubMed]
- James, S.R.; Link, P.A.; Karpf, A.R. Epigenetic regulation of X-linked cancer/germline antigen genes by DNMT1 and DNMT3b. Oncogene 2006, 25, 6975–6985. [Google Scholar] [CrossRef] [PubMed]
- De Smet, C.; Lurquin, C.; Lethe, B.; Martelange, V.; Boon, T. DNA methylation is the primary silencing mechanism for a set of germ line- and tumor-specific genes with a CpG-rich promoter. Mol. Cell. Biol. 1999, 19, 7327–7335. [Google Scholar] [CrossRef] [PubMed]
- Kimble, J. Molecular regulation of the mitosis/meiosis decision in multicellular organisms. Cold Spring Harb. Perspect. Biol. 2011, 3, a002683. [Google Scholar] [CrossRef] [PubMed]
- Roeder, G.S. Meiotic chromosomes: It takes two to tango. Genes Dev. 1997, 11, 2600–2621. [Google Scholar] [CrossRef] [PubMed]
- Baudat, F.; Buard, J.; Grey, C.; Fledel-Alon, A.; Ober, C.; Przeworski, M.; Coop, G.; de Massy, B. PRDM9 is a major determinant of meiotic recombination hotspots in humans and mice. Science 2010, 327, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Grey, C.; Barthes, P.; Chauveau-Le Friec, G.; Langa, F.; Baudat, F.; de Massy, B. Mouse PRDM9 DNA-binding specificity determines sites of histone H3 lysine 4 trimethylation for initiation of meiotic recombination. PLoS Biol. 2011, 9, e1001176. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.L.; Walker, M.; Kajita, S.; Petkov, P.M.; Paigen, K. PRDM9 binding organizes hotspot nucleosomes and limits Holliday junction migration. Genome Res. 2014, 24, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, J.; Aldeailej, I.; Anderson, R.; Almutairi, M.; Almatrafi, A.; Alsiwiehri, N.; Griffiths, K.; Stuart, N.; Wakeman, J.A.; Larcombe, L.; et al. Meta-analysis of clinical data using human meiotic genes identifies a novel cohort of highly restricted cancer-specific marker genes. Oncotarget 2012, 3, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.; Pan, J.; Cole, F.; Thelen, M.P.; Jasin, M.; Keeney, S. ATM controls meiotic double-strand-break formation. Nature 2011, 479, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef]
- Lindsey, S.F.; Byrnes, D.M.; Eller, M.S.; Rosa, A.M.; Dabas, N.; Escandon, J.; Grichnik, J.M. Potential role of meiosis proteins in melanoma chromosomal instability. J. Skin Cancer 2013, 2013, 190109. [Google Scholar] [CrossRef] [PubMed]
- Koslowski, M.; Tureci, O.; Bell, C.; Krause, P.; Lehr, H.A.; Brunner, J.; Seitz, G.; Nestle, F.O.; Huber, C.; Sahin, U. Multiple splice variants of lactate dehydrogenase C selectively expressed in human cancer. Cancer Res. 2002, 62, 6750–6755. [Google Scholar] [PubMed]
- Yang, F.; Eckardt, S.; Leu, N.A.; McLaughlin, K.J.; Wang, P.J. Mouse TEX15 is essential for DNA double-strand break repair and chromosomal synapsis during male meiosis. J. Cell Biol. 2008, 180, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Loriot, A.; Boon, T.; De Smet, C. Five new human cancer-germline genes identified among 12 genes expressed in spermatogonia. Int. J. Cancer 2003, 105, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Sehorn, M.G.; Sigurdsson, S.; Bussen, W.; Unger, V.M.; Sung, P. Human meiotic recombinase Dmc1 promotes ATP-dependent homologous DNA strand exchange. Nature 2004, 429, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Ianzini, F.; Kosmacek, E.A.; Nelson, E.S.; Napoli, E.; Erenpreisa, J.; Kalejs, M.; Mackey, M.A. Activation of meiosis-specific genes is associated with depolyploidization of human tumor cells following radiation-induced mitotic catastrophe. Cancer Res. 2009, 69, 2296–2304. [Google Scholar] [CrossRef] [PubMed]
- Shiohama, Y.; Ohtake, J.; Ohkuri, T.; Noguchi, D.; Togashi, Y.; Kitamura, H.; Nishimura, T. Identification of a meiosis-specific protein, MEIOB, as a novel cancer/testis antigen and its augmented expression in demethylated cancer cells. Immunol. Lett. 2014, 158, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Yang, F.; Leu, N.A.; Landaiche, J.; Handel, M.A.; Benavente, R.; La Salle, S.; Wang, P.J. MEIOB exhibits single-stranded DNA-binding and exonuclease activities and is essential for meiotic recombination. Nat. Commun. 2013, 4, 2788. [Google Scholar] [CrossRef] [PubMed]
- Souquet, B.; Abby, E.; Herve, R.; Finsterbusch, F.; Tourpin, S.; le Bouffant, R.; Duquenne, C.; Messiaen, S.; Martini, E.; Bernardino-Sgherri, J.; et al. MEIOB targets single-strand DNA and is necessary for meiotic recombination. PLoS Genet. 2013, 9, e1003784. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.H.; Choi, Y.; Erdin, S.U.; Yatsenko, S.A.; Kloc, M.; Yang, F.; Wang, P.J.; Meistrich, M.L.; Rajkovic, A. Hormad1 mutation disrupts synaptonemal complex formation, recombination, and chromosome segregation in mammalian meiosis. PLoS Genet. 2010, 6, e1001190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, K.; Lange, J.; Hached, K.; Fu, J.; Anastassiadis, K.; Roig, I.; Cooke, H.J.; Stewart, A.F.; Wassmann, K.; Jasin, M.; et al. Meiotic homologue alignment and its quality surveillance are controlled by mouse HORMAD1. Nat. Cell Biol. 2011, 13, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Aung, P.P.; Oue, N.; Mitani, Y.; Nakayama, H.; Yoshida, K.; Noguchi, T.; Bosserhoff, A.K.; Yasui, W. Systematic search for gastric cancer-specific genes based on SAGE data: Melanoma inhibitory activity and matrix metalloproteinase-10 are novel prognostic factors in patients with gastric cancer. Oncogene 2006, 25, 2546–2557. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.; Weekes, D.; Shah, V.; Gazinska, P.; Joshi, S.; Sidhu, B.; Gillett, C.; Pinder, S.; Vanoli, F.; Jasin, M.; et al. Genomic complexity profiling reveals that HORMAD1 overexpression contributes to homologous recombination deficiency in triple-negative breast cancers. Cancer Discov. 2015, 5, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Venditti, C.A.; Theiler, G.; Stevenson, B.J.; Iseli, C.; Gure, A.O.; Jongeneel, C.V.; Old, L.J.; Simpson, A.J. Identification of CT46/HORMAD1, an immunogenic cancer/testis antigen encoding a putative meiosis-related protein. Cancer Immun. 2005, 5, 9. [Google Scholar] [PubMed]
- Wojtasz, L.; Daniel, K.; Roig, I.; Bolcun-Filas, E.; Xu, H.; Boonsanay, V.; Eckmann, C.R.; Cooke, H.J.; Jasin, M.; Keeney, S.; et al. Mouse HORMAD1 and HORMAD2, two conserved meiotic chromosomal proteins, are depleted from synapsed chromosome axes with the help of TRIP13 AAA-ATPase. PLoS Genet. 2009, 5, e1000702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogo, H.; Tsutsumi, M.; Inagaki, H.; Ohye, T.; Kiyonari, H.; Kurahashi, H. HORMAD2 is essential for synapsis surveillance during meiotic prophase via the recruitment of ATR activity. Genes Cells 2012, 17, 897–912. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, J.; Hu, L.; Shi, X.; Zhou, Z.; Hu, Z.; Sha, J. HORMAD2/CT46.2, a novel cancer/testis gene, is ectopically expressed in lung cancer tissues. Mol. Hum. Reprod. 2012, 18, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Chung, J.Y.; Kitano, H.; Takikita, M.; Cho, H.; Noh, K.H.; Kim, T.W.; Ylaya, K.; Hanaoka, J.; Fukuoka, J.; Hewitt, S.M. Synaptonemal complex protein 3 as a novel prognostic marker in early stage non-small cell lung cancer. Hum. Pathol. 2013, 44, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.H.; Noh, K.H.; Kim, J.H.; Bae, H.C.; Lin, K.Y.; Monie, A.; Pai, S.I.; Hung, C.F.; Wu, T.C.; Kim, T.W. Ectopic expression of X-linked lymphocyte-regulated protein pM1 renders tumor cells resistant to antitumor immunity. Cancer Res. 2010, 70, 3062–3070. [Google Scholar] [CrossRef] [PubMed]
- Costa, Y.; Speed, R.; Ollinger, R.; Alsheimer, M.; Semple, C.A.; Gautier, P.; Maratou, K.; Novak, I.; Hoog, C.; Benavente, R.; et al. Two novel proteins recruited by synaptonemal complex protein 1 (SYCP1) are at the centre of meiosis. J. Cell Sci. 2005, 118, 2755–2762. [Google Scholar] [CrossRef] [PubMed]
- Ollinger, R.; Alsheimer, M.; Benavente, R. Mammalian protein SCP1 forms synaptonemal complex-like structures in the absence of meiotic chromosomes. Mol. Biol. Cell 2005, 16, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Tureci, O.; Sahin, U.; Zwick, C.; Koslowski, M.; Seitz, G.; Pfreundschuh, M. Identification of a meiosis-specific protein as a member of the class of cancer/testis antigens. Proc. Natl. Acad. Sci. USA 1998, 95, 5211–5216. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Huang, S.; Xie, X.; Stockert, E.; Chen, Y.T.; Kubuschok, B.; Pfreundschuh, M. Expression of cancer-testis genes in human hepatocellular carcinomas. Cancer Immun. 2002, 2, 11. [Google Scholar] [PubMed]
- Sahin, U.; Koslowski, M.; Tureci, O.; Eberle, T.; Zwick, C.; Romeike, B.; Moringlane, J.R.; Schwechheimer, K.; Feiden, W.; Pfreundschuh, M. Expression of cancer testis genes in human brain tumors. Clin. Cancer Res. 2000, 6, 3916–3922. [Google Scholar] [PubMed]
- Oba-Shinjo, S.M.; Caballero, O.L.; Jungbluth, A.A.; Rosemberg, S.; Old, L.J.; Simpson, A.J.; Marie, S.K. Cancer-testis (CT) antigen expression in medulloblastoma. Cancer Immun. 2008, 8, 7. [Google Scholar] [PubMed]
- Lee, M.H.; Son, E.I.; Kim, E.; Kim, I.S.; Yim, M.B.; Kim, S.P. Expression of cancer-testis genes in brain tumors. J. Korean Neurosurg. Soc. 2008, 43, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Kubuschok, B.; Xie, X.; Jesnowski, R.; Preuss, K.D.; Romeike, B.F.; Neumann, F.; Regitz, E.; Pistorius, G.; Schilling, M.; Scheunemann, P.; et al. Expression of cancer testis antigens in pancreatic carcinoma cell lines, pancreatic adenocarcinoma and chronic pancreatitis. Int. J. Cancer 2004, 109, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, H.; Maeda, Y.; Ishida, T.; Gnjatic, S.; Sato, E.; Mori, F.; Sugiyama, D.; Ito, A.; Fukumori, Y.; Utsunomiya, A.; et al. Cancer/testis antigens are novel targets of immunotherapy for adult T-cell leukemia/lymphoma. Blood 2012, 119, 3097–3104. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, K.; Watanabe, Y. Chromosome cohesion in mitosis and meiosis. J. Cell Sci. 2007, 120, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Llano, E.; Herran, Y.; Garcia-Tunon, I.; Gutierrez-Caballero, C.; de Alava, E.; Barbero, J.L.; Schimenti, J.; de Rooij, D.G.; Sanchez-Martin, M.; Pendas, A.M. Meiotic cohesin complexes are essential for the formation of the axial element in mice. J. Cell Biol. 2012, 197, 877–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Iwai, T.; Yokota, T.; Yamashita, M. Temporally and spatially selective loss of Rec8 protein from meiotic chromosomes during mammalian meiosis. J. Cell Sci. 2003, 116, 2781–2790. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hirano, T. RAD21L, a novel cohesin subunit implicated in linking homologous chromosomes in mammalian meiosis. J. Cell Biol. 2011, 192, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.M.; Dabas, N.; Byrnes, D.M.; Eller, M.S.; Grichnik, J.M. Germ cell proteins in melanoma: Prognosis, diagnosis, treatment, and theories on expression. J. Skin Cancer 2012, 2012, 621968. [Google Scholar] [CrossRef] [PubMed]
- Sasai, K.; Katayama, H.; Stenoien, D.L.; Fujii, S.; Honda, R.; Kimura, M.; Okano, Y.; Tatsuka, M.; Suzuki, F.; Nigg, E.A.; et al. Aurora-C kinase is a novel chromosomal passenger protein that can complement Aurora-B kinase function in mitotic cells. Cell Motil. Cytoskelet. 2004, 59, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Kimmins, S.; Crosio, C.; Kotaja, N.; Hirayama, J.; Monaco, L.; Hoog, C.; van Duin, M.; Gossen, J.A.; Sassone-Corsi, P. Differential functions of the Aurora-B and Aurora-C kinases in mammalian spermatogenesis. Mol. Endocrinol. 2007, 21, 726–739. [Google Scholar] [CrossRef] [PubMed]
- Ulisse, S.; Delcros, J.G.; Baldini, E.; Toller, M.; Curcio, F.; Giacomelli, L.; Prigent, C.; Ambesi-Impiombato, F.S.; D′Armiento, M.; Arlot-Bonnemains, Y. Expression of Aurora kinases in human thyroid carcinoma cell lines and tissues. Int. J. Cancer 2006, 119, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Zekri, A.; Lesan, V.; Ghaffari, S.H.; Tabrizi, M.H.; Modarressi, M.H. Gene amplification and overexpression of Aurora-C in breast and prostate cancer cell lines. Oncol. Res. 2012, 20, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Matsuda, Y.; Yoshioka, T.; Okano, Y. Cell cycle-dependent expression and centrosome localization of a third human aurora/Ipl1-related protein kinase, AIK3. J. Biol. Chem. 1999, 274, 7334–7340. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Futamura, M.; Yoshimi, N.; Sano, J.; Katada, M.; Takagi, Y.; Kimura, M.; Yoshioka, T.; Okano, Y.; Saji, S. Centrosomal kinases, HsAIRK1 and HsAIRK3, are overexpressed in primary colorectal cancers. Jpn. J. Cancer Res. 2000, 91, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Sagata, N.; Oskarsson, M.; Copeland, T.; Brumbaugh, J.; van de Woude, G.F. Function of c-mos proto-oncogene product in meiotic maturation in Xenopus oocytes. Nature 1988, 335, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Xerri, L.; Charpin, C.; Hassoun, J.; Birnbaum, D.; Delapeyriere, O. Mos oncogene expression in human ovarian tumors. Anticancer Res. 1991, 11, 1629–1634. [Google Scholar] [PubMed]
- Li, C.C.; Chen, E.; O′Connell, C.D.; Longo, D.L. Detection of c-mos proto-oncogene expression in human cells. Oncogene 1993, 8, 1685–1691. [Google Scholar] [PubMed]
- Dube, C.; Leclerc, P.; Baba, T.; Reyes-Moreno, C.; Bailey, J.L. The proacrosin binding protein, sp32, is tyrosine phosphorylated during capacitation of pig sperm. J. Androl. 2005, 26, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Niida, Y.; Michikawa, Y.; Kashiwabara, S.; Kodaira, K.; Takenaka, M.; Kohno, N.; Gerton, G.L.; Arai, Y. An acrosomal protein, sp32, in mammalian sperm is a binding protein specific for two proacrosins and an acrosin intermediate. J. Biol. Chem. 1994, 269, 10133–10140. [Google Scholar] [PubMed]
- Tammela, J.; Uenaka, A.; Ono, T.; Noguchi, Y.; Jungbluth, A.A.; Mhawech-Fauceglia, P.; Qian, F.; Schneider, S.; Sharma, S.; Driscoll, D.; et al. OY-TES-1 expression and serum immunoreactivity in epithelial ovarian cancer. Int. J. Oncol. 2006, 29, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Kurashige, T.; Harada, N.; Noguchi, Y.; Saika, T.; Niikawa, N.; Aoe, M.; Nakamura, S.; Higashi, T.; Hiraki, A.; et al. Identification of proacrosin binding protein sp32 precursor as a human cancer/testis antigen. Proc. Natl. Acad. Sci. USA. 2001, 98, 3282–3287. [Google Scholar] [CrossRef] [PubMed]
- Cappell, K.M.; Sinnott, R.; Taus, P.; Maxfield, K.; Scarbrough, M.; Whitehurst, A.W. Multiple cancer testis antigens function to support tumor cell mitotic fidelity. Mol. Cell. Biol. 2012, 32, 4131–4140. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Obata, Y.; Yoshida, M.; Stockert, E.; Williamson, B.; Jungbluth, A.A.; Chen, Y.T.; Old, L.J.; Scanlan, M.J. Immunomic analysis of human sarcoma. Proc. Natl. Acad. Sci. USA 2003, 100, 2651–2656. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Eckardt, S.; Leu, N.A.; Buffone, M.G.; Zhou, J.; Gerton, G.L.; McLaughlin, K.J.; Wang, P.J. Inactivation of Nxf2 causes defects in male meiosis and age-dependent depletion of spermatogonia. Dev. Biol. 2009, 330, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.; Bolton, L.M.; Johnson, P.; Elliott, T.; Murray, N. Breast cancer is a promising target for vaccination using cancer-testis antigens known to elicit immune responses. Breast Cancer Res. 2007, 9, R46. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Kwon, T.K.; Kim, I.H.; Sohn, S.S.; Kim, Y.S.; Kim, C.I.; Bae, O.S.; Lee, K.S.; Lee, K.D.; Lee, C.S.; et al. A new strategy for the diagnosis of MAGE-expressing cancers. J. Immunol. Methods 2002, 266, 79–86. [Google Scholar] [CrossRef]
- Tsai, J.R.; Chong, I.W.; Chen, Y.H.; Yang, M.J.; Sheu, C.C.; Chang, H.C.; Hwang, J.J.; Hung, J.Y.; Lin, S.R. Differential expression profile of MAGE family in non-small-cell lung cancer. Lung Cancer 2007, 56, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Zammatteo, N.; Lockman, L.; Brasseur, F.; de Plaen, E.; Lurquin, C.; Lobert, P.E.; Hamels, S.; Boon, T.; Remacle, J. DNA microarray to monitor the expression of MAGE-A genes. Clin. Chem. 2002, 48, 25–34. [Google Scholar] [PubMed]
- Brown, P.R.; Miki, K.; Harper, D.B.; Eddy, E.M. A-kinase anchoring protein 4 binding proteins in the fibrous sheath of the sperm flagellum. Biol. Reprod. 2003, 68, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.B.; Prendes, M.J.; Kidd, J.L.; Sternberg, H.; West, M.D.; Wagner, J. Elevated expression of cancer/testis antigen FSIP1 in ER-positive breast tumors. Biomark. Med. 2013, 7, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, M.; Jin, Z.; Wang, D.; Sun, M.; Zhao, X.; Zhao, Z.; Lei, H.; Li, M.; Liu, C. Expression and clinicopathological significance of FSIP1 in breast cancer. Oncotarget 2015, 6, 10658–10666. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.; Essmann, F.; Kletke, A.; Rio, P.; Hanenberg, H.; Wetzel, W.; Schulze-Osthoff, K.; Nurnberg, B.; Piekorz, R.P. The transforming acidic coiled coil 3 protein is essential for spindle-dependent chromosome alignment and mitotic survival. J. Biol. Chem. 2007, 282, 29273–29283. [Google Scholar] [CrossRef] [PubMed]
- Ha, G.H.; Kim, J.L.; Petersson, A.; Oh, S.; Denning, M.F.; Patel, T.; Breuer, E.K. TACC3 deregulates the DNA damage response and confers sensitivity to radiation and PARP inhibition. Oncogene 2015, 34, 1667–1678. [Google Scholar] [CrossRef] [PubMed]
- Soccio, R.E.; Adams, R.M.; Romanowski, M.J.; Sehayek, E.; Burley, S.K.; Breslow, J.L. The cholesterol-regulated StarD4 gene encodes a StAR-related lipid transfer protein with two closely related homologues, StarD5 and StarD6. Proc. Natl. Acad. Sci. USA 2002, 99, 6943–6948. [Google Scholar] [CrossRef] [PubMed]
- Lichten, M.; Goldman, A.S. Meiotic recombination hotspots. Annu. Rev. Genet. 1995, 29, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Yoshida, K.; Matsui, Y. A histone H3 methyltransferase controls epigenetic events required for meiotic prophase. Nature 2005, 438, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Shannon, M.; Richardson, L.; Christian, A.; Handel, M.A.; Thelen, M.P. Differential gene expression of mammalian SPO11/TOP6A homologs during meiosis. FEBS Lett. 1999, 462, 329–334. [Google Scholar] [CrossRef]
- Collins, C.; Rommens, J.M.; Kowbel, D.; Godfrey, T.; Tanner, M.; Hwang, S.I.; Polikoff, D.; Nonet, G.; Cochran, J.; Myambo, K.; et al. Positional cloning of ZNF217 and NABC1: Genes amplified at 20q13.2 and overexpressed in breast carcinoma. Proc. Natl. Acad. Sci. USA 1998, 95, 8703–8708. [Google Scholar] [PubMed]
- Courjal, F.; Cuny, M.; Rodriguez, C.; Louason, G.; Speiser, P.; Katsaros, D.; Tanner, M.M.; Zeillinger, R.; Theillet, C. DNA amplifications at 20q13 and MDM2 define distinct subsets of evolved breast and ovarian tumours. Br. J. Cancer 1996, 74, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.M.; Tirkkonen, M.; Kallioniemi, A.; Collins, C.; Stokke, T.; Karhu, R.; Kowbel, D.; Shadravan, F.; Hintz, M.; Kuo, W.L.; et al. Increased copy number at 20q13 in breast cancer: Defining the critical region and exclusion of candidate genes. Cancer Res. 1994, 54, 4257–4260. [Google Scholar] [PubMed]
- Hofmann, O.; Caballero, O.L.; Stevenson, B.J.; Chen, Y.T.; Cohen, T.; Chua, R.; Maher, C.A.; Panji, S.; Schaefer, U.; Kruger, A.; et al. Genome-wide analysis of cancer/testis gene expression. Proc. Natl. Acad. Sci. USA 2008, 105, 20422–20427. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.J.; McCarrey, J.R.; Yang, F.; Page, D.C. An abundance of X-linked genes expressed in spermatogonia. Nat. Genet. 2001, 27, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Neale, M.J.; Keeney, S. Clarifying the mechanics of DNA strand exchange in meiotic recombination. Nature 2006, 442, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gu, Y.; Zhang, K.; Xie, K.; Zhu, M.; Dai, N.; Jiang, Y.; Guo, X.; Liu, M.; Dai, J.; et al. Systematic identification of genes with a cancer-testis expression pattern in 19 cancer types. Nat. Commun. 2016, 7, 10499. [Google Scholar] [CrossRef] [PubMed]
- Heyting, C. Synaptonemal complexes: Structure and function. Curr. Opin. Cell Biol. 1996, 8, 389–396. [Google Scholar] [CrossRef]
- Page, S.L.; Hawley, R.S. The genetics and molecular biology of the synaptonemal complex. Annu. Rev. Cell Dev. Biol. 2004, 20, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, R.L.; Offenberg, H.H.; Dietrich, A.J.; Riesewijk, A.; van Iersel, M.; Heyting, C. A coiled-coil related protein specific for synapsed regions of meiotic prophase chromosomes. EMBO J. 1992, 11, 5091–5100. [Google Scholar] [PubMed]
- Niemeyer, P.; Tureci, O.; Eberle, T.; Graf, N.; Pfreundschuh, M.; Sahin, U. Expression of serologically identified tumor antigens in acute leukemias. Leuk. Res. 2003, 27, 655–660. [Google Scholar] [CrossRef]
- Strunnikov, A. Cohesin complexes with a potential to link mammalian meiosis to cancer. Cell Regen. (Lond.) 2013, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Ezan, F.; Cremet, J.Y.; Fautrel, A.; Gilot, D.; Lambert, M.; Benaud, C.; Troadec, M.B.; Prigent, C. Overexpression of active Aurora-C kinase results in cell transformation and tumour formation. PLoS ONE 2011, 6, e26512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobasheri, M.B.; Shirkoohi, R.; Zendehdel, K.; Jahanzad, I.; Talebi, S.; Afsharpad, M.; Modarressi, M.H. Transcriptome analysis of the cancer/testis genes, DAZ1, AURKC, and TEX101, in breast tumors and six breast cancer cell lines. Tumour Biol. 2015, 36, 8201–8206. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.W.; Wang, Y.C.; Chang-Liao, P.Y.; Lin, Y.J.; Yang, S.T.; Tsou, J.H.; Chang, K.C.; Liu, Y.W.; Tseng, J.T.; Lee, C.T.; et al. Overexpression of Aurora-C interferes with the spindle checkpoint by promoting the degradation of Aurora-B. Cell Death Dis. 2014, 5, e1106. [Google Scholar] [CrossRef] [PubMed]
- Funabiki, H.; Wynne, D.J. Making an effective switch at the kinetochore by phosphorylation and dephosphorylation. Chromosoma 2013, 122, 135–158. [Google Scholar] [CrossRef] [PubMed]
- Kops, G.J.; Weaver, B.A.; Cleveland, D.W. On the road to cancer: Aneuploidy and the mitotic checkpoint. Nat. Rev. Cancer 2005, 5, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Prieur-Carrillo, G.; Chu, K.; Lindqvist, J.; Dewey, W.C. Computerized video time-lapse (CVTL) analysis of the fate of giant cells produced by X-irradiating EJ30 human bladder carcinoma cells. Radiat. Res. 2003, 159, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Erenpreisa, J.A.; Cragg, M.S.; Fringes, B.; Sharakhov, I.; Illidge, T.M. Release of mitotic descendants by giant cells from irradiated Burkitt's lymphoma cell line. Cell Biol. Int. 2000, 24, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Kalejs, M.; Ivanov, A.; Plakhins, G.; Cragg, M.S.; Emzinsh, D.; Illidge, T.M.; Erenpreisa, J. Upregulation of meiosis-specific genes in lymphoma cell lines following genotoxic insult and induction of mitotic catastrophe. BMC Cancer 2006, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Beasley, M.D.; Warren, W.D.; van der Horst, G.T.; McKay, M.J. Absence of mouse REC8 cohesin promotes synapsis of sister chromatids in meiosis. Dev. Cell 2005, 8, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Erenpreisa, J.; Cragg, M.S.; Salmina, K.; Hausmann, M.; Scherthan, H. The role of meiotic cohesin REC8 in chromosome segregation in gamma irradiation-induced endopolyploid tumour cells. Exp. Cell Res. 2009, 315, 2593–2603. [Google Scholar] [CrossRef] [PubMed]
- Whitehurst, A.W.; Bodemann, B.O.; Cardenas, J.; Ferguson, D.; Girard, L.; Peyton, M.; Minna, J.D.; Michnoff, C.; Hao, W.; Roth, M.G.; et al. Synthetic lethal screen identification of chemosensitizer loci in cancer cells. Nature 2007, 446, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Whitehurst, A.W.; Xie, Y.; Purinton, S.C.; Cappell, K.M.; Swanik, J.T.; Larson, B.; Girard, L.; Schorge, J.O.; White, M.A. Tumor antigen acrosin binding protein normalizes mitotic spindle function to promote cancer cell proliferation. Cancer Res. 2010, 70, 7652–7661. [Google Scholar] [CrossRef] [PubMed]
- Halazonetis, T.D.; Gorgoulis, V.G.; Bartek, J. An oncogene-induced DNA damage model for cancer development. Science 2008, 319, 1352–1355. [Google Scholar] [CrossRef] [PubMed]
- Maxfield, K.E.; Taus, P.J.; Corcoran, K.; Wooten, J.; Macion, J.; Zhou, Y.; Borromeo, M.; Kollipara, R.K.; Yan, J.; Xie, Y.; et al. Comprehensive functional characterization of cancer-testis antigens defines obligate participation in multiple hallmarks of cancer. Nat. Commun. 2015, 6, 8840. [Google Scholar] [CrossRef] [PubMed]
- Song, M.H.; Kim, Y.R.; Lee, J.W.; Lee, C.H.; Lee, S.Y. Cancer/testis antigen NY-SAR-35 enhances cell proliferation, migration, and invasion. Int. J. Oncol. 2016, 48, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Greve, K.B.; Lindgreen, J.N.; Terp, M.G.; Pedersen, C.B.; Schmidt, S.; Mollenhauer, J.; Kristensen, S.B.; Andersen, R.S.; Relster, M.M.; Ditzel, H.J.; et al. Ectopic expression of cancer/testis antigen SSX2 induces DNA damage and promotes genomic instability. Mol. Oncol. 2015, 9, 437–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Por, E.; Byun, H.J.; Lee, E.J.; Lim, J.H.; Jung, S.Y.; Park, I.; Kim, Y.M.; Jeoung, D.I.; Lee, H. The cancer/testis antigen CAGE with oncogenic potential stimulates cell proliferation by up-regulating cyclins D1 and E in an AP-1- and E2F-dependent manner. J. Biol. Chem. 2010, 285, 14475–14485. [Google Scholar] [CrossRef] [PubMed]
- D′Arcy, P.; Maruwge, W.; Wolahan, B.; Ma, L.; Brodin, B. Oncogenic functions of the cancer-testis antigen SSX on the proliferation, survival, and signaling pathways of cancer cells. PLoS ONE 2014, 9, e95136. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Protein | Function in Meiosis | Species of Functional Studies | Expression in Cancer |
|---|---|---|---|
| PRDM9 | Meiotic recombination hotspot activator [29,30,31] | Mice and human protein | Embryonal carcinoma, astrocytoma, colon, prostate, breast, ovary, melanoma and leukemia cancer cell lines [32] |
| SPO11 | Meiosis-specific nuclease [29,33,34] | Mice and yeast | Melanoma and lung cancer cell lines, and melanoma and cervical cancer tissue [35,36] |
| TEX15 | Mediates loading of DSB repair proteins onto DNA (at DSB sites) [37] | Mice | Bladder carcinomas, cutaneous melanoma, esophageal carcinomas, head and neck carcinomas, lung carcinoma, neuroblastomas, prostate tumors, renal tumors and sarcomas [38] |
| DMC1 | Recombinase/DNA repair protein [39] | Human protein | Cervical cancer tissue [40] |
| MEIOB | 3′ to 5′ exonuclease [41,42,43] | Mice and human cell lines | Liver, leukemia and lung cancer cell lines [41] |
| HORMAD1 | Mediates homologous recombination, synaptonemal complex formation and recruitment of ATR kinase activity to unsynapsed chromatin [44,45] | Mice | Gastric [46], breast (including triple-negative breast cancer), non-small cell lung cancer (NSCLC), esophageal, endometrical, bladder and colon cancer tissue [47,48], and breast, ovarian, melanoma, cervical, NSCLC and small lung cancer cell lines (CT Gene Database) [48] |
| HORMAD2 | Proposed function: Similar to HORMAD1 [49,50] | Mice | Lung cancer tissue [51] |
| SCP-3/SYCP3 | Synaptonemal complex protein [52] | Mice | NSCLC [53] and cervical cancer tissue [54] |
| SYCE1 | Synaptonemal complex protein [55] | Mice | Breast cancer, melanoma and leukemia cancer cell lines (CT Gene Database) |
| SCP-1/SYCP1 | Synaptonemal complex protein [56] | Human cell line | Breast, melanoma, brain (glioma, glioblastoma, schwanoma, medulloblastoma, meningioma, astrocytoma, oligoastrocytoma and pilocytic astrocytoma), gastric, lung (including NSCLC), renal cell, stomach [57,58,59,60,61] and pancreatic cancer tissue [62], and ovarian (CT Gene Database), ATLL [63] and pancreatic [62] cancer cell lines |
| REC8 | Component of meiosis-specific cohesin complex [64,65,66,67] | Mice | Melanoma cell lines [35,68] |
| RAD21L | Component of meiosis-specific cohesin complex [64,65,66,67] | Mice | Colon, breast, ovarian, embryonal carcinoma, cervix and leukemia cancer cell lines [32] |
| SMC1β | Component of meiosis-specific cohesin complex [64,65,66,67] | Mice | Breast, leukemia and embryonal carcinoma cancer cell lines [32] |
| STAG3 | Component of meiosis-specific cohesin complex [64,65,66,67] | Mice | Various cancers (reported in Oncomine, September 2012) |
| AURKC | Component of the of the meiotic chromosomal passenger complex [69,70] | Mice and human cell lines | Breast, cervical, liver, prostate, thyroid carcinoma cancer cells lines [71,72,73], and colorectal and thyroid cancer tissue [71,74] |
| MOS | Regulates oocyte maturation [75] | Xenopus | Ovarian cancer tissue [76] and neuroblastoma and cervical carcinoma-derived cell lines [77] |
| ACRBP | Role in spermatogenesis and sperm capacitation [78,79] | Porcine | Ovarian cancer cell line and cancer tissue [80], bladder, breast, lung, liver and colon cancer tissue [81], and sarcoma, prostate, multiple myeloma, chronic myeloid leukemia, lung and ovarian cancer cell lines (CT Gene Database) |
| FMR1NB | Role in regulating microtubule nucleation and/or anchoring events in the mitotic spindle (suggested role from CT Gene Database, based on [82]) | Human cell lines | Melanoma, sarcoma, lung, breast, bladder, esophageal and ovarian cancer tissue [83], and sarcoma, multiple myeloma, chronic myeloid leukemia, choriocarcinoma, lung and breast cancer cell lines (CT Gene Database) |
| NXF2 | Nuclear RNA export factor, important for regulation of meiosis and maintenance of spermatogonial stem cells [84] | Mice | Melanoma, sarcoma, prostate, multiple myeloma, chronic myeloid leukemia, choriocarcinoma, lung, ovarian and colon cancer cell lines (CT Gene Database), and bladder, colorectal carcinoma, lung, melanoma, esophageal, head and neck, neuroblastoma, prostate, sarcoma and thyroid cancer tissue [38] |
| MAGEA5 | Breast [85], lung, head and neck [86], lung adenocarcinoma and squamous cell cancer tissue [87], and melanoma and thyroid carcinoma cell lines [88] | ||
| FSIP1 | Component of the fibrous sheath structure, unique for spermatogenic cells [89] | Mice and yeast | Breast cancer tissue [90,91] |
| TACC3 | Mitosis: Plays a role in spindle stability and kinetochore-microtubule interactions [92] | Human cell lines | Breast, lung, colon and liver cancer tissue [93] |
| STARD6 | Involved in transport of lipids [94] | Mice |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, A.Y.; Gjerstorff, M.F. Ectopic Expression of Testis Germ Cell Proteins in Cancer and Its Potential Role in Genomic Instability. Int. J. Mol. Sci. 2016, 17, 890. https://doi.org/10.3390/ijms17060890
Nielsen AY, Gjerstorff MF. Ectopic Expression of Testis Germ Cell Proteins in Cancer and Its Potential Role in Genomic Instability. International Journal of Molecular Sciences. 2016; 17(6):890. https://doi.org/10.3390/ijms17060890
Chicago/Turabian StyleNielsen, Aaraby Yoheswaran, and Morten Frier Gjerstorff. 2016. "Ectopic Expression of Testis Germ Cell Proteins in Cancer and Its Potential Role in Genomic Instability" International Journal of Molecular Sciences 17, no. 6: 890. https://doi.org/10.3390/ijms17060890
APA StyleNielsen, A. Y., & Gjerstorff, M. F. (2016). Ectopic Expression of Testis Germ Cell Proteins in Cancer and Its Potential Role in Genomic Instability. International Journal of Molecular Sciences, 17(6), 890. https://doi.org/10.3390/ijms17060890