
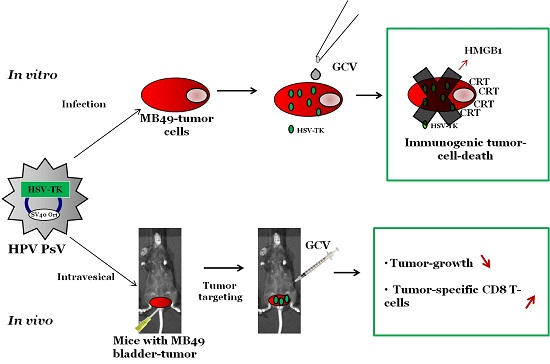
Immunogenic Human Papillomavirus Pseudovirus-Mediated Suicide-Gene Therapy for Bladder Cancer

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
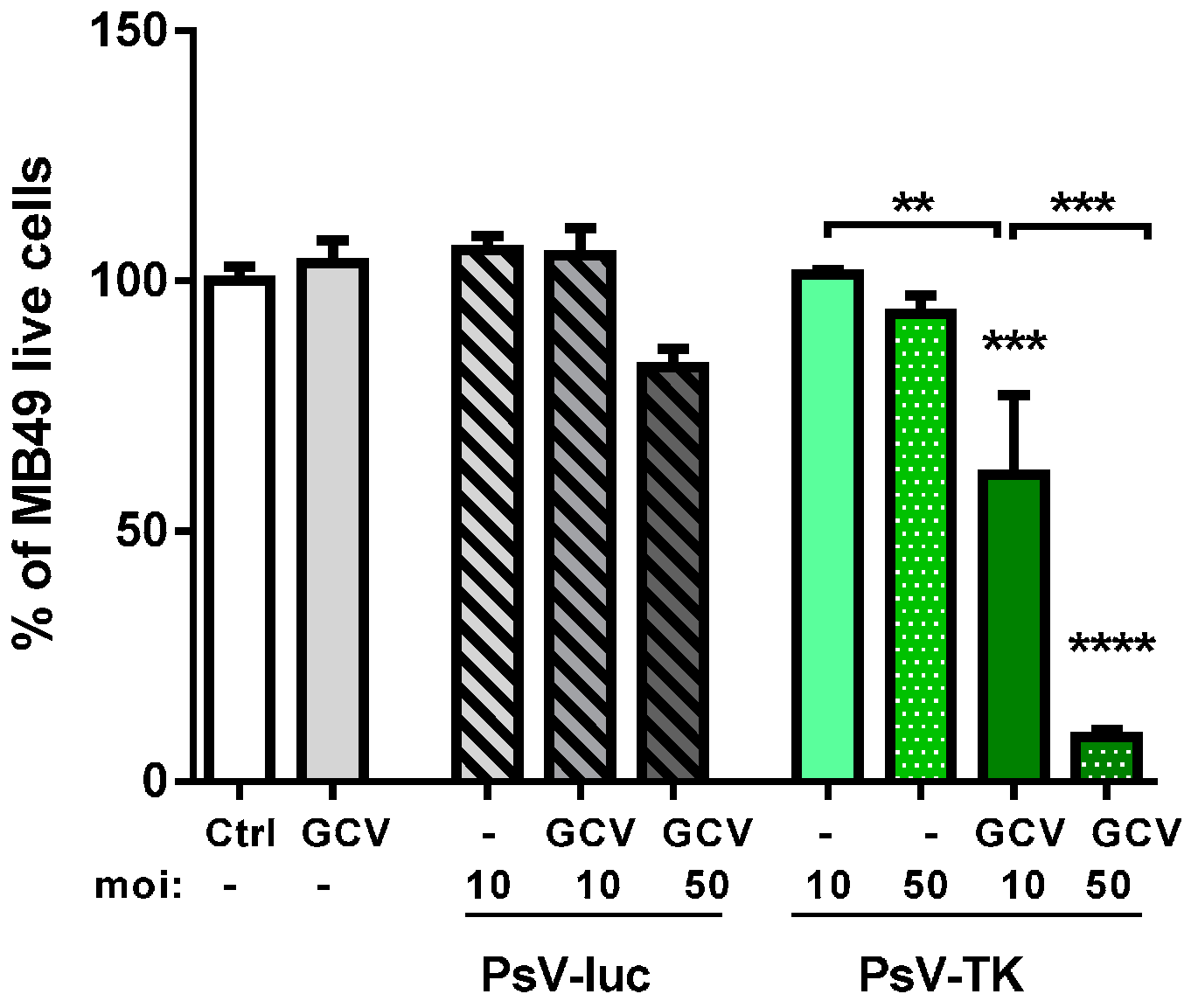
2.1. In Vitro Induction of Immunogenic Tumor Cell-Death (TCD) by PsV Encoding for Thymidine Kinase (PsV-TK)/Ganciclovir (GCV)
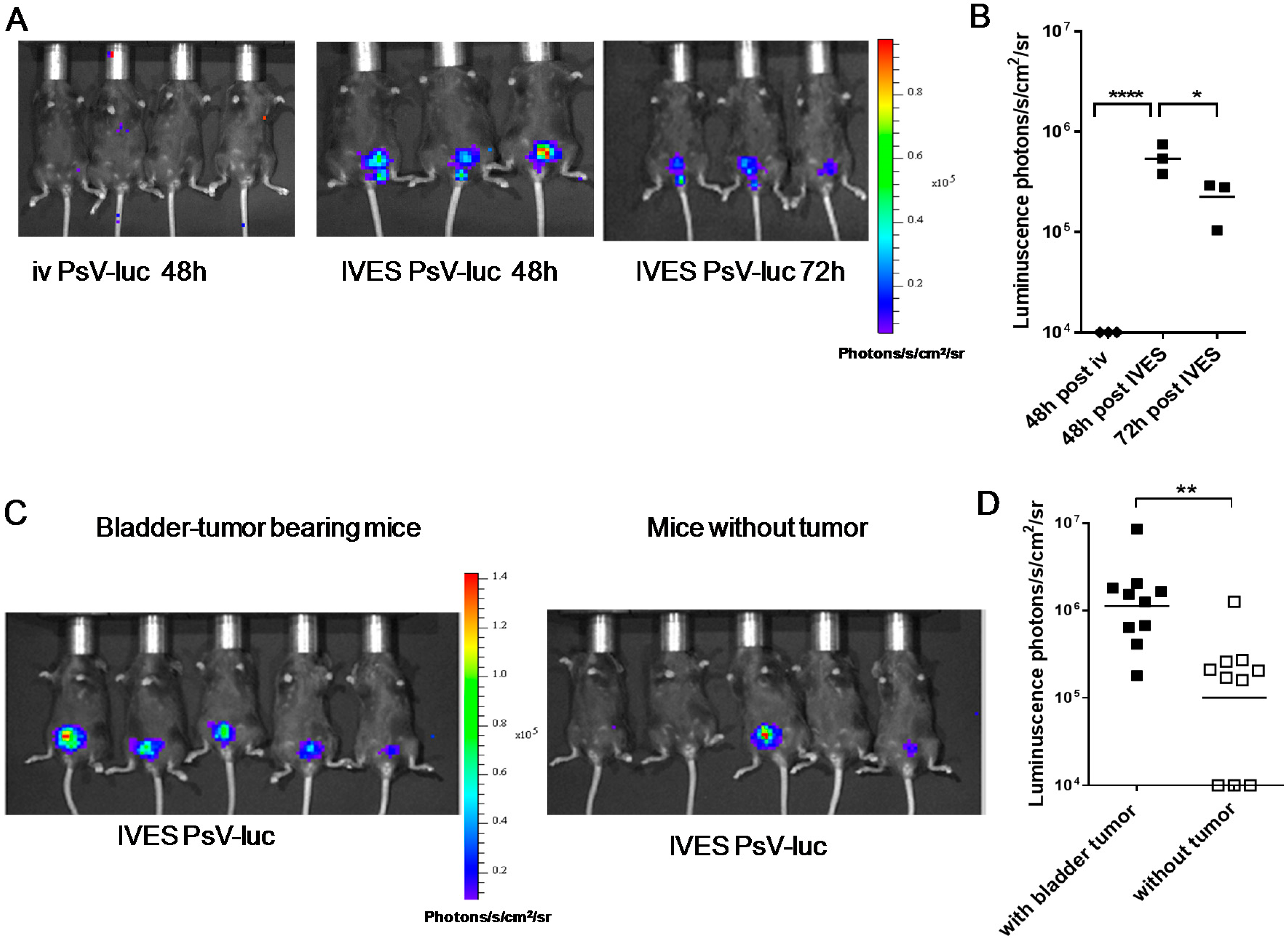
2.2. Preferential Targeting of MB49 Bladder-Tumors by PsV
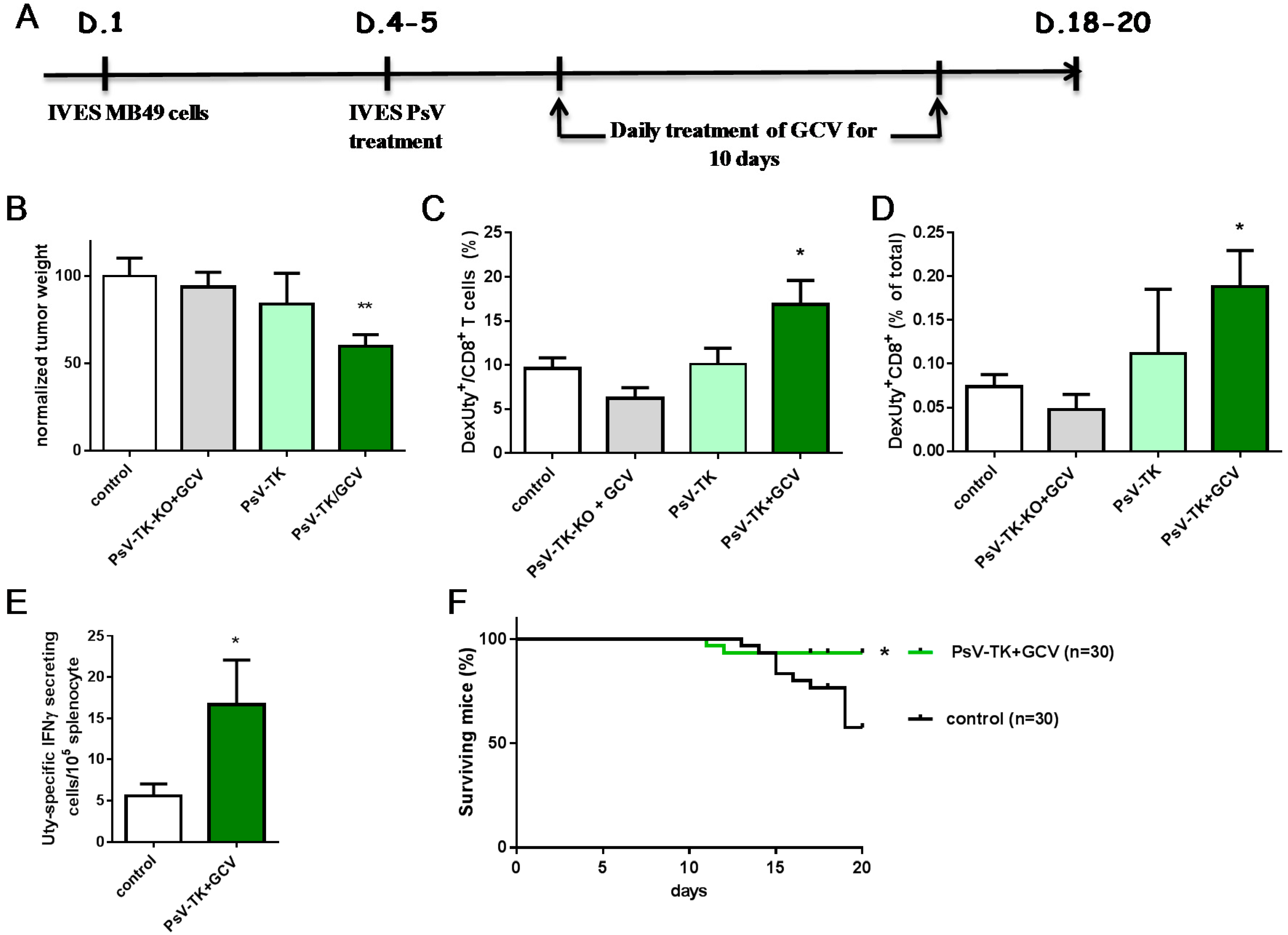
2.3. PsV-TK/GCV Therapy Reduced Bladder-Tumor Growth and Increased Tumor-Specific CD8 T Cells and Mice Survival
3. Discussion
4. Materials and Methods
4.1. MB49 Cells and the Orthotopic Bladder Tumor Model
4.2. PsV Production and PsV/GCV Suicide Gene Therapy in Vitro and in Vivo
4.3. Preparation of Murine Cells, IFN-γ ELISPOT and T-Cell Labeling
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BCG | Bacillus-Calmette Guérin |
| CRT | Calreticuline |
| GCV | Ganciclovir |
| HSPG | Heparan sulfate proteoglycans |
| HSV | Herpes simplex virus |
| HMGB-1 | High-mobility group box-1 |
| HPV | Human papillomavirus |
| IVES | intravesical |
| NMIBC | Non muscle-invasive bladder cancer |
| PsV | Pseudovirion |
| TCD | Tumor cell-death |
| TK | Thymidine kinase |
| TK-KO | Non functional TK |
| TRLU | transducing relative light units |
References
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2012. CA Cancer J. Clin. 2012, 62, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Steliarova-Foucher, E.; Lortet-Tieulent, J.; Rosso, S.; Coebergh, J.W.; Comber, H.; Forman, D.; Bray, F. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries in 2012. Eur. J. Cancer 2013, 49, 1374–1403. [Google Scholar] [CrossRef] [PubMed]
- Babjuk, M.; Burger, M.; Zigeuner, R.; Shariat, S.F.; van Rhijn, B.W.; Comperat, E.; Sylvester, R.J.; Kaasinen, E.; Bohle, A.; Palou-Redorta, J.; et al. EAU guidelines on non-muscle-invasive urothelial carcinoma of the bladder: Update 2013. Eur. Urol. 2013, 64, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Babjuk, M.; Oosterlinck, W.; Sylvester, R.; Kaasinen, E.; Bohle, A.; Palou-Redorta, J.; Roupret, M. EAU guidelines on non-muscle-invasive urothelial carcinoma of the bladder, the 2011 update. Eur. Urol. 2011, 59, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.R.; Roupret, M. Contemporary management of patients with high-risk non-muscle-invasive bladder cancer who fail intravesical BCG therapy. World J. Urol. 2011, 29, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Kepp, O.; Zitvogel, L. Immunogenic cell death in cancer therapy. Annu. Rev. Immunol. 2013, 31, 51–72. [Google Scholar] [CrossRef] [PubMed]
- Portsmouth, D.; Hiavaty, J.; Renner, M. Suicide genes for cancer therapy. Mol. Asp. Med. 2007, 1, 4–41. [Google Scholar] [CrossRef] [PubMed]
- Fillat, C.; Carrio, M.; Cascante, A.; Sangro, B. Suicide gene therapy mediated by the Herpes Simplex virus thymidine kinase gene/Ganciclovir system: Fifteen years of application. Curr. Gene Ther. 2003, 3, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Tani, K. Multimodal immunogenic cancer cell death as a consequence of anticancer cytotoxic treatments. Cell Death Differ. 2014, 21, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.T.; Day, P.M.; Kines, R.C. Current understanding of the mechanism of HPV infection. Gynecol. Oncol. 2010, 118, S12–S17. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.; Pastrana, D.V.; Lowy, D.R.; Schiller, J. Efficient intracellular assembly of papillomaviral vectors. J. Virol. 2004, 78, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Monie, A.; Kang, T.H.; Hung, C.-F.; Roden, R.; Wu, T.C. Efficient delivery of DNA vaccines using human papillomavirs pseudovirions. Gene Ther. 2010, 17, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.S.; Kines, R.C.; Corbett, K.S.; Nicewonger, J.; Johnson, T.R.; Chen, M.; La Vigne, D.; Roberts, J.N.; Cuburu, N.; Schiller, J.T.; et al. Mucosal delivery of human papillomarivus pseurovius-encapsidated plasmids impoves the potency of DNA vaccination. Mucosal Immunol. 2010, 3, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Cuburu, N.; Graham, B.S.; Buck, C.B.; Kines, R.C.; Pang, Y.Y.; Day, P.M.; Lowy, D.R.; Schiller, J.T. Intravaginal immunization with HPV vectors induces tissue-resident CD8+ T cell responses. J. Clin. Investig. 2012, 122, 4606–4620. [Google Scholar] [CrossRef] [PubMed]
- Selinka, H.C.; Florin, L.; Patel, H.D.; Freitag, K.; Schmidtke, M.; Makarov, V.A.; Sapp, M. Inhibition of transfer to secondary receptors by heparan sulfate-binding drug or antibody induces noninfectious uptake of human papillomavirus. J. Virol. 2007, 81, 10970–10980. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, R.D.; Yang, Y.; Kelly, T.; MacLeod, V.; Dai, Y.; Theus, A. Enzymatic remodeling of heparan sulfate proteoglycans within the tumor microenvironment: Growth regulation and the prospect of new cancer therapies. J. Cell. Biochem. 2005, 96, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Kines, R.C.; Cerio, R.J.; Roberts, J.N.; Thompson, C.D.; de Los Pinos, E.; Lowy, D.R.; Schiller, J.T. Human papillomavirus capsids preferentially bind and infect tumor cells. Int. J. Cancer 2016, 138, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Summerhayes, I.C.; Franks, L.M. Effects of donor age on neoplastic transformation of adult mouse bladder epithelium in vitro. J. Natl. Cancer Inst. 1979, 62, 1017–1023. [Google Scholar] [PubMed]
- Obeid, M.; Tesniere, A.; Ghiringhelli, F.; Fimia, G.M.; Apetoh, L.; Perfettini, J.L.; Castedo, M.; Mignot, G.; Panaretakis, T.; Casares, N.; et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat. Med. 2007, 13, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Longet, S.; Schiller, J.T.; Bobst, M.; Jichlinski, P.; Nardelli-Haefliger, D. A murine genital-challenge model is a sensitive measure of protective antibodies against human papillomavirus infection. J. Virol. 2011, 85, 13253–13259. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.B.; Thompson, C.D. Production of papillomavirus-based gene transfer vectors. Curr. Protoc. Cell Biol. 2007. [Google Scholar] [CrossRef]
- Liu, Q.Y.; Summers, W.C. Site-directed mutagenesis of a nucleotide-binding domain in HSV-1 thymidine kinase: Effects on catalytic activity. Virology 1988, 163, 638–642. [Google Scholar] [CrossRef]
- Yang, A.S.; Monken, C.E.; Lattime, E.C. Intratumoral vaccination with vaccinia-expressed tumor antigen and granulocytes macrophage colony-stimulating factor overcomes immunological ingorance to tumor antigen. Cancer Res. 2003, 63, 6956–6961. [Google Scholar] [PubMed]
- Millrain, M.; Chandler, P.; Dazzi, F.; Scott, D.; Simpson, E.; Dyson, P.J. Examination of HY response: T cell expansion, immunodominance, and cross-priming revealed by HY tetramer analysis. J. Immunol. 2001, 167, 3756–3764. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.F.; Chiang, A.J.; Tsai, H.H.; Pomper, M.G.; Kang, T.H.; Roden, R.R.; Wu, T.C. Ovarian cancer gene therapy using HPV-16 pseudovirion carrying the HSV-tk gene. PLoS ONE 2012, 7, e40983. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.; Patel, A.; Heston, W.; Larchian, W. Mouse orthotopic models for bladder cancer research. BJU Int. 2009, 104, 1286–1291. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.N.; Buck, C.B.; Thompson, C.D.; Kines, R.; Bernardo, M.; Choyke, P.L.; Lowy, D.; Schiller, J. Genital transmissionof HPV in a mouse model is potentiated by nonoxynol-9 and inhibited by carrageenan. Nat. Med. 2007, 13, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Lopez-Beltran, A.; Bostwick, D. Bladder Pathology; Wiley-Blackwell: Hoboken, NJ, USA, 2012. [Google Scholar]
- Jichlinski, P.; Lovisa, B. High magnification cystoscopy in the primary diagnosis of bladder tumors. Curr. Opin. Urol. 2011, 21, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Kamat, A.M.; Flaig, T.W.; Grossman, H.B.; Konety, B.; Lamm, D.; O’Donnell, M.A.; Uchio, E.; Efstathiou, J.A.; Taylor, J.A., III. Expert consensus document: Consensus statement on best practice management regarding the use of intravesical immunotherapy with BCG for bladder cancer. Nat. Rev. Urol. 2015, 12, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Duarte, S.; Carle, G.; Faneca, H.; de Lima, M.C.; Pierrefite-Carle, V. Suicide gene therapy in cancer: Where do we stand now? Cancer Lett. 2012, 324, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.M.; Ramesh, R.; Marrogi, A.J. Immune system in suicide-gene therapy. Lancet 1997, 9044, 2–3. [Google Scholar] [CrossRef]
- Bochner, B.H. Gene therapy in bladder cancer. Curr. Opin. Urol. 2008, 18, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Kosai, K.; Xu, B.; Pham-Nguyen, K.; Contant, C.; Finegold, M.J.; Woo, S.L. Combination suicide and cytokine gene therapy for hepatic metastases of colon carcinoma: Sustained antitumor immunity prolongs animal survival. Cancer Res. 1996, 16, 3758–3762. [Google Scholar]
- Chen, S.H.; Chen, X.H.; Wang, Y.; Kosai, K.; Finegold, M.J.; Rich, S.S.; Woo, S.L. Combination gene therapy for liver metastasis of colon carcinoma in vivo. Proc. Natl. Acad. Sci. USA 1995, 7, 2577–2581. [Google Scholar] [CrossRef]
- Ramesh, R.; Munshi, A.; Marrogi, A.J.; Freeman, S.M. Enhancement of tumor killing using a combination of tumor immunization and HSV-tk suicide gene therapy. Int. J. Cancer 1999, 80, 380–386. [Google Scholar] [CrossRef]
- Muhammad, G.; Candolfi, M.; King, G.D.; Yagiz, K.; Foulad, D.; Mineharu, Y.; Kroeger, K.M.; Treuer, K.A.; Nichols, W.S.; Sanderson, N.S.; et al. Antiglioma immunological memory in response to conditional cytotoxic/immune-stimulatory gene therapy: Humoral and cellular immunita lead to tumor regression. Clin. Cancer Res. 2009, 19, 6113–6127. [Google Scholar] [CrossRef] [PubMed]
- Lab of Cellular Oncology Technical Files. NCI, NIH. Available online: http://home.ccr.cancer.gov/LCO/plasmids.asp (accessed on 12 July 2016).
- Revaz, V.; Debonneville, A.; Bobst, M.; Nardelli-Haefliger, D. Monitoring of vaccine-specific γ interferon inductionin in genital mucosa of mice by real-time reverse transcription-PCR. Clin. Vaccine Immunol. 2008, 5, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Domingos-Pereira, S.; Decrausaz, L.; Derre, L.; Bobst, M.; Romero, P.; Schiller, J.T.; Jichlinski, P.; Nardelli-Haefliger, D. Intravaginal TLR agonists increase local vaccine-specific CD8 T cells and human papillomavirus-associated genital-tumor regression in mice. Mucosal Immunol. 2013, 6, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Decrausaz, L.; Domingos-Pereira, S.; Duc, M.; Bobst, M.; Romero, P.; Schiller, J.T.; Jichlinski, P.; Nardelli-Haefliger, D. Parenteral but not mucosal immunization is able to induce regression of human papillomavirus associated genital tumors. Int. J. Cancer 2011, 129, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Shen, T.; Wientjes, M.G.; O’Donnell, M.A.; Au, J.L. Intravesical treatments of bladder cancer: Review. Pharm. Res. 2008, 25, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Botteman, M.F.; Pashos, C.L.; Redaelli, A.; Laskin, B.; Hauser, R. The health economics of bladder cancer: A comprehensive review of the published literature. Pharmacoeoconomics 2003, 21, 1315–1330. [Google Scholar] [CrossRef]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hojeij, R.; Domingos-Pereira, S.; Nkosi, M.; Gharbi, D.; Derré, L.; Schiller, J.T.; Jichlinski, P.; Nardelli-Haefliger, D. Immunogenic Human Papillomavirus Pseudovirus-Mediated Suicide-Gene Therapy for Bladder Cancer. Int. J. Mol. Sci. 2016, 17, 1125. https://doi.org/10.3390/ijms17071125
Hojeij R, Domingos-Pereira S, Nkosi M, Gharbi D, Derré L, Schiller JT, Jichlinski P, Nardelli-Haefliger D. Immunogenic Human Papillomavirus Pseudovirus-Mediated Suicide-Gene Therapy for Bladder Cancer. International Journal of Molecular Sciences. 2016; 17(7):1125. https://doi.org/10.3390/ijms17071125
Chicago/Turabian StyleHojeij, Rim, Sonia Domingos-Pereira, Marianne Nkosi, Dalila Gharbi, Laurent Derré, John T. Schiller, Patrice Jichlinski, and Denise Nardelli-Haefliger. 2016. "Immunogenic Human Papillomavirus Pseudovirus-Mediated Suicide-Gene Therapy for Bladder Cancer" International Journal of Molecular Sciences 17, no. 7: 1125. https://doi.org/10.3390/ijms17071125
APA StyleHojeij, R., Domingos-Pereira, S., Nkosi, M., Gharbi, D., Derré, L., Schiller, J. T., Jichlinski, P., & Nardelli-Haefliger, D. (2016). Immunogenic Human Papillomavirus Pseudovirus-Mediated Suicide-Gene Therapy for Bladder Cancer. International Journal of Molecular Sciences, 17(7), 1125. https://doi.org/10.3390/ijms17071125