
Cytoprotection against Hypoxic and/or MPP+ Injury: Effect of δ–Opioid Receptor Activation on Caspase 3


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
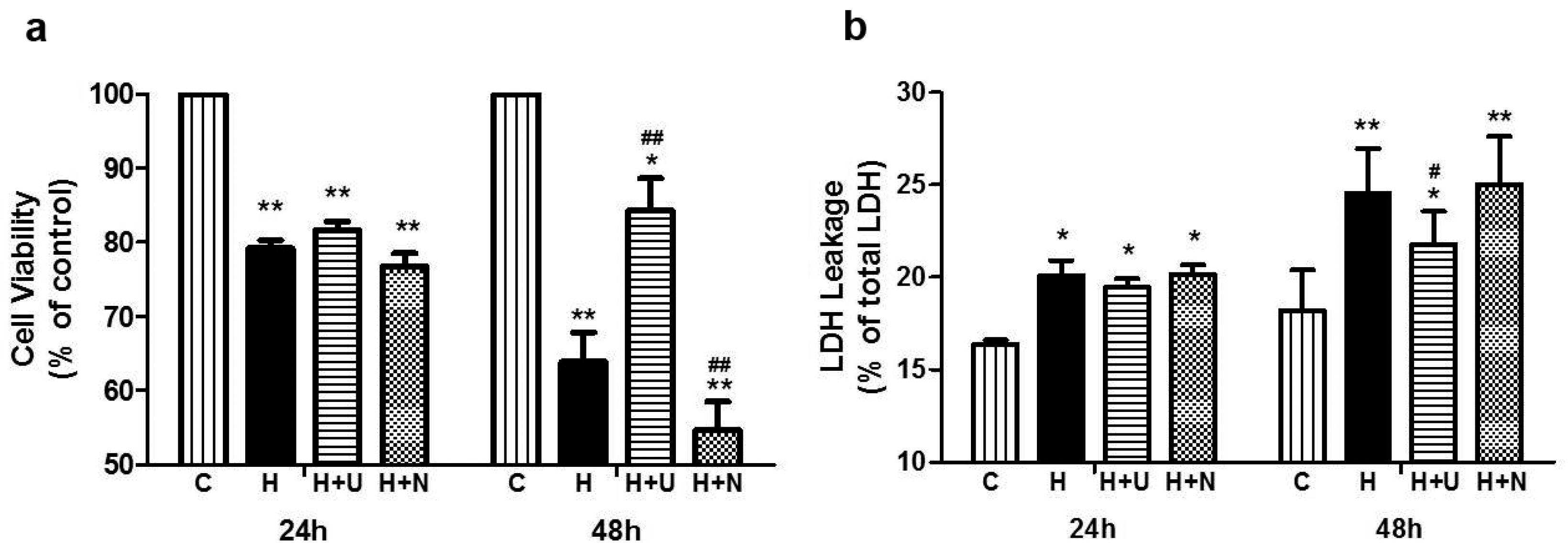
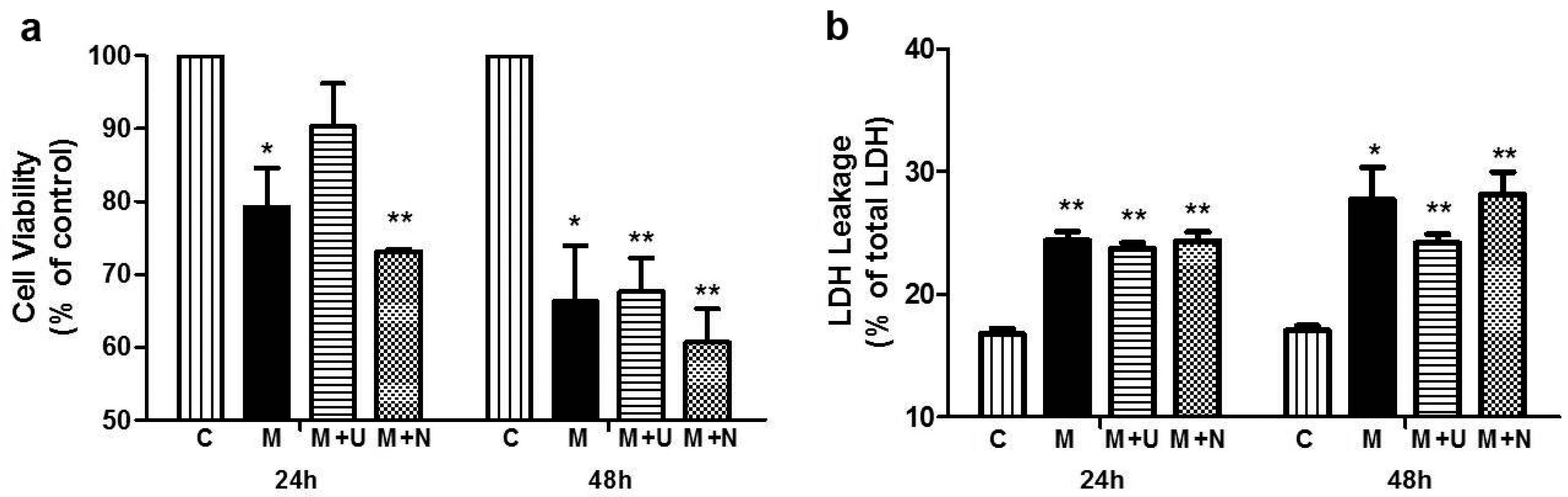
2.1. Prolonged Hypoxia and MPP+ Stress Caused Severe PC12 Cell Injury
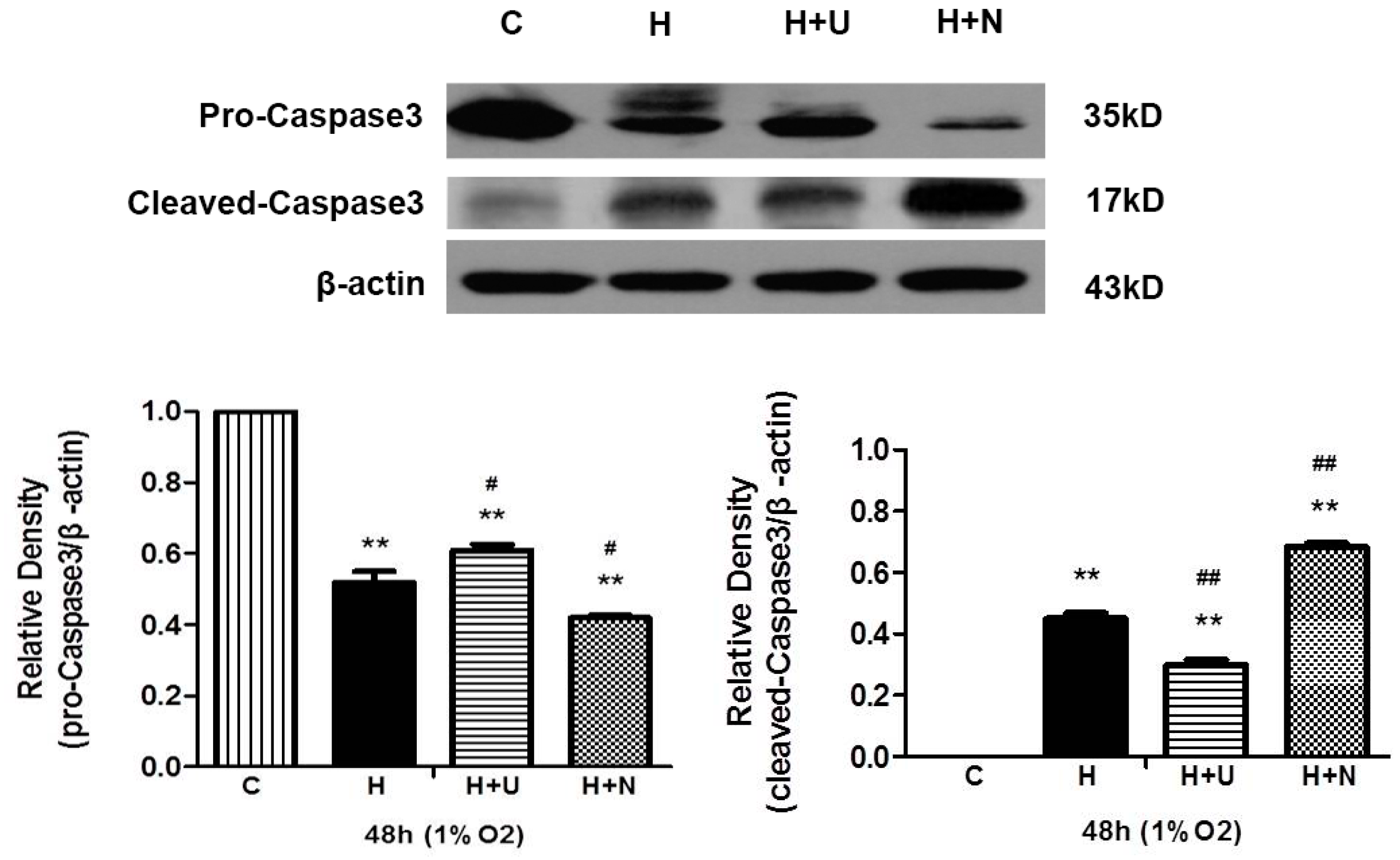
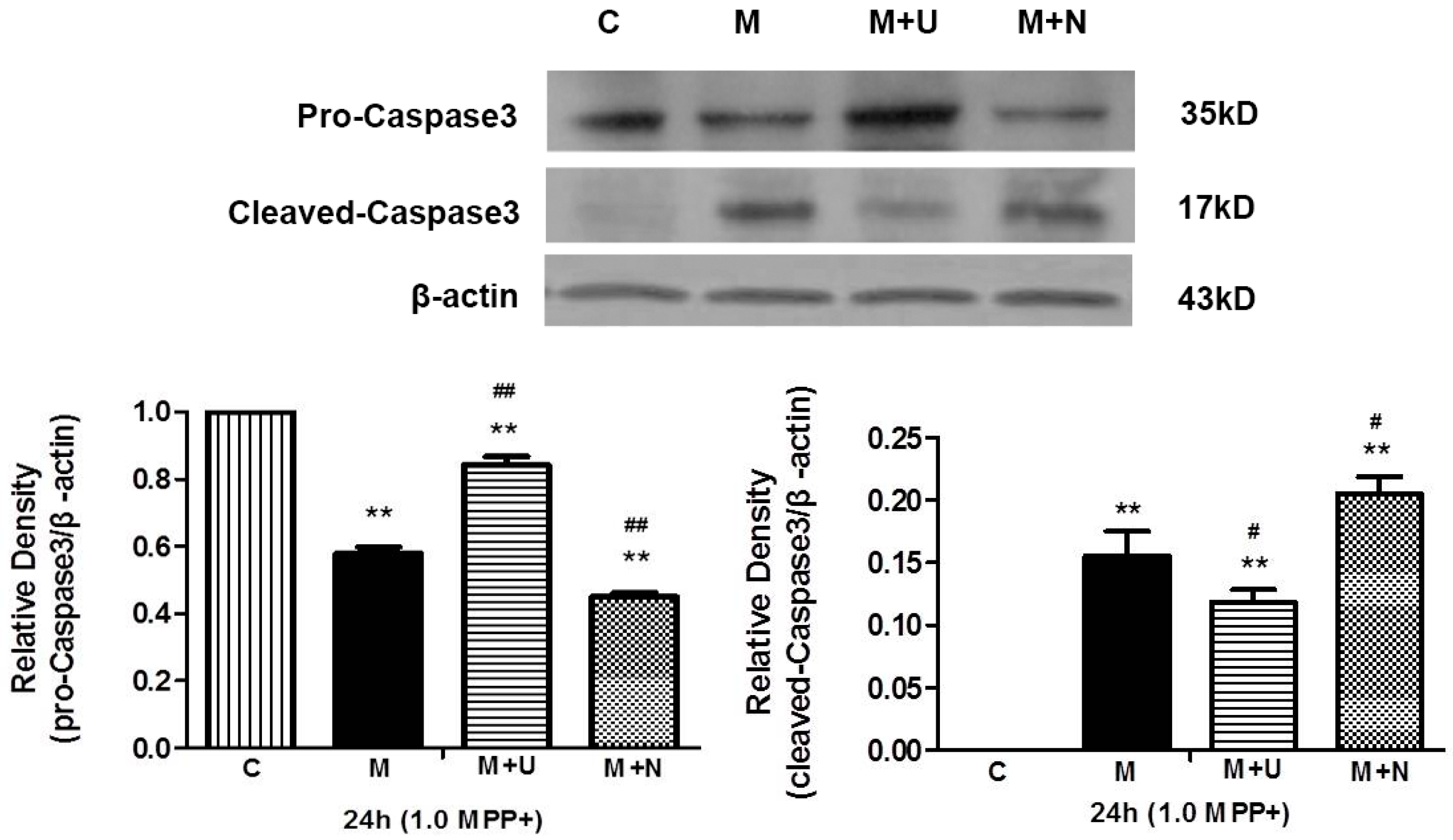
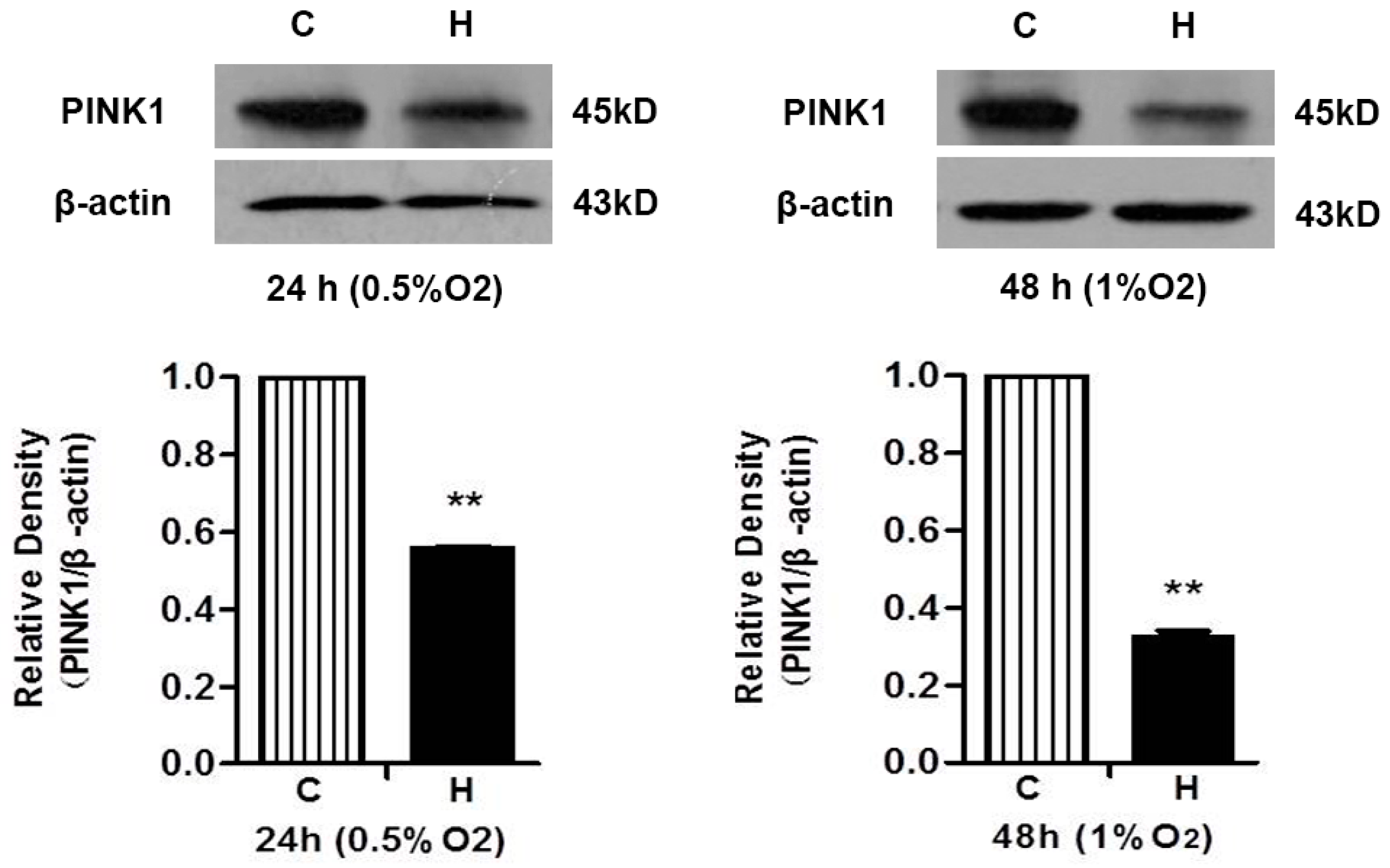
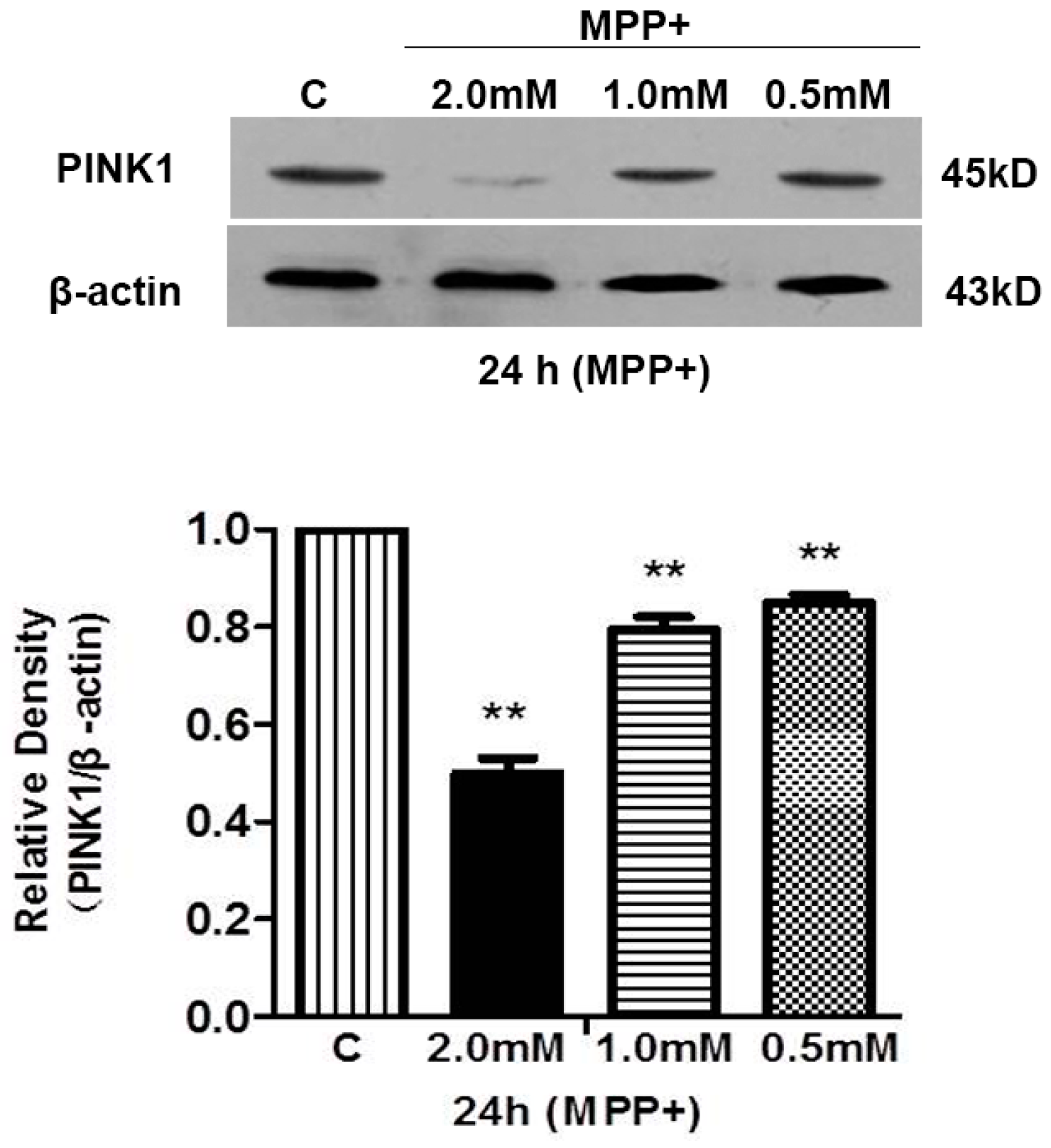
2.2. Both Prolonged Hypoxia and MPP+ Insults Increased the Level of Cleaved Caspase 3 with a Reduction of PINK1 Protein
2.3. DOR Activation Attenuated PC12 Cell Injury Induced by Hypoxic and MPP+ Stress
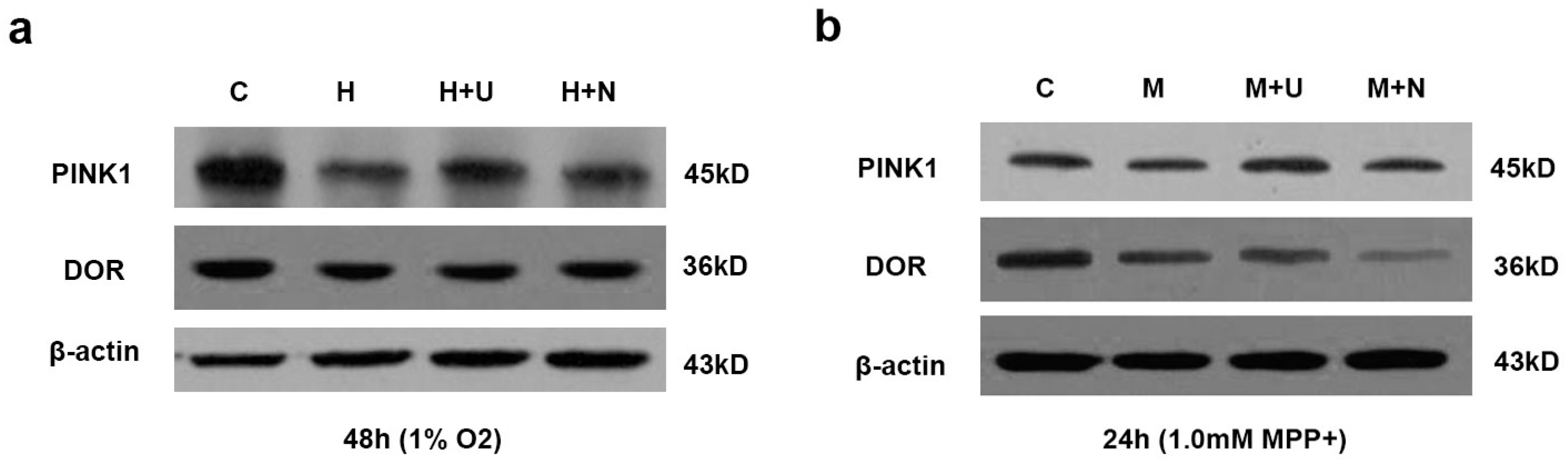
2.4. DOR Activation Upregulated PINK1 Expression in Hypoxia or MPP+ Stress
2.5. DOR Activation Inhibited Hypoxia/MPP+-Induced Production of Cleaved Caspase 3
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Cultures and Experimental Groups
- C: normoxic control.
- H: hypoxia.
- H + U: DOR activation with UFP-512 (5 μM) in hypoxic condition.
- H + N: DOR inhibition with naltrindole (1 μM) in hypoxic condition.
- M: MPP+.
- M + U: DOR activation with UFP-512 (5 μM) under MPP+ insult.
- M + N: DOR inhibition with naltrindole (1 μM) under MPP+ insult.
4.3. Cell Viability Assay
Lactate Dehydrogenase Assessment
4.4. Western Blotting
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchi, S.; Frosini, D.; Ripoli, A.; Nicoletti, V.; Linsalata, G.; Bonuccelli, U.; Ceravolo, R. Serotonergic antidepressant drugs and L-dopa-induced dyskinesias in Parkinson’s disease. Acta Neurol. Scand. 2015, 131, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Politis, M.; Lindvall, O. Clinical application of stem cell therapy in Parkinson’s disease. BMC Med. 2012, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.R.; Maguire-Zeiss, K.A. Gene therapy in Parkinson’s disease: Rationale and current status. CNS Drugs 2010, 24, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhu, W.; Tian, L.; Zhang, J.; Li, Z. MCT4-mediated expression of EAAT1 is involved in the resistance to hypoxia injury in astrocyte-neuron co-cultures. Neurochem. Res. 2015, 40, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Karunasinghe, R.N.; Lipski, J. Oxygen and glucose deprivation (OGD)-induced spreading depression in the Substantia Nigra. Brain Res. 2013, 1527, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Grande, B.; Blackabey, V.; Gittens, B.; Pinteaux, E.; Denes, A. Loss of substance P and inflammation precede delayed neurodegeneration in the substantia nigra after cerebral ischemia. Brain Behav. Immun. 2013, 29, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Siuda, J.; Jasinska-Myga, B.; Boczarska-Jedynak, M.; Opala, G.; Fiesel, F.C.; Moussaud-Lamodiere, E.L.; Scarffe, L.A.; Dawson, V.L.; Ross, O.A.; Springer, W.; et al. Early-onset Parkinson’s disease due to PINK1 p.Q456X mutation—Clinical and functional study. Parkinsonism Relat. Disord. 2014, 20, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Yunfu, W.; Guangjian, L.; Ping, Z.; Yanpeng, S.; Xiaoxia, F.; Wei, H.; Jiang, Y.; Jingquan, H.; Songlin, W.; Hongyan, Z.; et al. PINK1 and its familial Parkinson’s disease-associated mutation regulate brain vascular endothelial inflammation. J. Mol. Neurosci. 2014, 53, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Minami, A.; Nakanishi, A.; Matsuda, S.; Kitagishi, Y.; Ogura, Y. Function of alpha-synuclein and PINK1 in Lewy body dementia (Review). Int. J. Mol. Med. 2015, 35, 3–9. [Google Scholar] [PubMed]
- Kazlauskaite, A.; Muqit, M.M. PINK1 and Parkin—Mitochondrial interplay between phosphorylation and ubiquitylation in Parkinson’s disease. FEBS J. 2015, 282, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, C.; van Dyk, H.C.; Engelbrecht, L.; van der Westhuizen, F.H.; Kinnear, C.; Loos, B.; Bardien, S. Curcumin Rescues a PINK1 Knock Down SH-SY5Y Cellular Model of Parkinson’s Disease from Mitochondrial Dysfunction and Cell Death. Mol. Neurobiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Burbulla, L.F.; Fitzgerald, J.C.; Stegen, K.; Westermeier, J.; Thost, A.K.; Kato, H.; Mokranjac, D.; Sauerwald, J.; Martins, L.M.; Woitalla, D.; et al. Mitochondrial proteolytic stress induced by loss of mortalin function is rescued by Parkin and PINK1. Cell Death Dis. 2014, 5, e1180. [Google Scholar] [CrossRef] [PubMed]
- Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Analysis of mitochondrial dysfunction during cell death. Methods Mol. Biol. 2015, 1264, 385–393. [Google Scholar] [PubMed]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Calcium and mitochondria in the regulation of cell death. Biochem. Biophys. Res. Commun. 2015, 460, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, G.S.; Al-Harbi, S.; Almasan, A. Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis. Methods Mol. Biol. 2015, 1219, 1–9. [Google Scholar]
- Sakahira, H.; Enari, M.; Nagata, S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 1998, 391, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zou, H.; Slaughter, C.; Wang, X. DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell 1997, 89, 175–184. [Google Scholar] [CrossRef]
- Liu, X.; Li, P.; Widlak, P.; Zou, H.; Luo, X.; Garrard, W.T.; Wang, X. The 40-kDa subunit of DNA fragmentation factor induces DNA fragmentation and chromatin condensation during apoptosis. Proc. Natl. Acad. Sci. USA 1998, 95, 8461–8466. [Google Scholar] [CrossRef] [PubMed]
- Rathnasamy, G.; Sivakumar, V.; Rangarajan, P.; Foulds, W.S.; Ling, E.A.; Kaur, C. NF-kappaB-mediated nitric oxide production and activation of caspase-3 cause retinal ganglion cell death in the hypoxic neonatal retina. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5878–5889. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gibney, G.T.; Zhao, P.; Xia, Y. Neuroprotective role of delta-opioid receptors in cortical neurons. Am. J. Physiol. Cell Physiol. 2002, 282, C1225–C1234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qian, H.; Zhao, P.; Hong, S.S.; Xia, Y. Rapid hypoxia preconditioning protects cortical neurons from glutamate toxicity through delta-opioid receptor. Stroke J. Cereb. Circ. 2006, 37, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.C.; Qian, H.; Ghassemi, F.; Zhao, P.; Xia, Y. Oxygen-sensitive-opioid receptor-regulated survival and death signals: Novel insights into neuronal preconditioning and protection. J. Biol. Chem. 2005, 280, 16208–16218. [Google Scholar] [CrossRef] [PubMed]
- Monti, C.; Bondi, H.; Urbani, A.; Fasano, M.; Alberio, T. Systems biology analysis of the proteomic alterations induced by MPP(+), a Parkinson’s disease-related mitochondrial toxin. Front. Cell. Neurosci. 2015, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, R.; Li, C.; Wu, C.; Xu, S. MPTP/MPP+ suppresses activation of protein C in Parkinson’s disease. J. Alzheimer Dis. JAD 2015, 43, 133–142. [Google Scholar]
- Ding, A.X.; Sun, G.; Argaw, Y.G.; Wong, J.O.; Easwaran, S.; Montell, D.J. CasExpress reveals widespread and diverse patterns of cell survival of caspase-3 activation during development. eLife 2016, 5, e10936. [Google Scholar] [CrossRef] [PubMed]
- Arena, G.; Gelmetti, V.; Torosantucci, L.; Vignone, D.; Lamorte, G.; De Rosa, P.; Cilia, E.; Jonas, E.A.; Valente, E.M. PINK1 protects against cell death induced by mitochondrial depolarization, by phosphorylating Bcl-xL and impairing its pro-apoptotic cleavage. Cell Death Differ. 2013, 20, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.A.; Corti, O.; Brice, A. Mitochondrial dysfunctions in Parkinson’s disease. Rev. Neurol. 2014, 170, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.R.; Chesselet, M.F. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease. Prog. Neurobiol. 2013, 106–107, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.R.; Li, Y.Q.; Hua, C.; Li, S.J.; Zhao, G.; Song, H.M.; Yu, M.X.; Huang, Q. Oxidative stress inhibits growth and induces apoptotic cell death in human U251 glioma cells via the caspase-3-dependent pathway. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4068–4075. [Google Scholar] [PubMed]
- Meng, X.W.; Fraser, M.J.; Feller, J.M.; Ziegler, J.B. Caspase-3-dependent and caspase-3-independent pathways leading to chromatin DNA fragmentation in HL-60 cells. Apoptosis Int. J. Program. Cell Death 2000, 5, 61–67. [Google Scholar] [CrossRef]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar] [CrossRef]
- Lax, A.; Soler, F.; Fernandez-Belda, F. Mitochondrial damage as death inducer in heart-derived H9c2 cells: More than one way for an early demise. J. Bioenerg. Biomembr. 2009, 41, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Rakovic, A.; Shurkewitsch, K.; Seibler, P.; Grunewald, A.; Zanon, A.; Hagenah, J.; Krainc, D.; Klein, C. Phosphatase and tensin homolog (PTEN)-induced putative kinase 1 (PINK1)-dependent ubiquitination of endogenous Parkin attenuates mitophagy: Study in human primary fibroblasts and induced pluripotent stem cell-derived neurons. J. Biol. Chem. 2013, 288, 2223–2237. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hu, G.K. PINK1 protects cortical neurons from thapsigargin-induced oxidative stress and neuronal apoptosis. Biosci. Rep. 2015, 35, e00174. [Google Scholar] [CrossRef] [PubMed]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Zhi, F.; Shao, N.; Wang, R.; Yang, Y.; Xia, Y. Cytoprotection against Hypoxic and/or MPP+ Injury: Effect of δ–Opioid Receptor Activation on Caspase 3. Int. J. Mol. Sci. 2016, 17, 1179. https://doi.org/10.3390/ijms17081179
Xu Y, Zhi F, Shao N, Wang R, Yang Y, Xia Y. Cytoprotection against Hypoxic and/or MPP+ Injury: Effect of δ–Opioid Receptor Activation on Caspase 3. International Journal of Molecular Sciences. 2016; 17(8):1179. https://doi.org/10.3390/ijms17081179
Chicago/Turabian StyleXu, Yuan, Feng Zhi, Naiyuan Shao, Rong Wang, Yilin Yang, and Ying Xia. 2016. "Cytoprotection against Hypoxic and/or MPP+ Injury: Effect of δ–Opioid Receptor Activation on Caspase 3" International Journal of Molecular Sciences 17, no. 8: 1179. https://doi.org/10.3390/ijms17081179
APA StyleXu, Y., Zhi, F., Shao, N., Wang, R., Yang, Y., & Xia, Y. (2016). Cytoprotection against Hypoxic and/or MPP+ Injury: Effect of δ–Opioid Receptor Activation on Caspase 3. International Journal of Molecular Sciences, 17(8), 1179. https://doi.org/10.3390/ijms17081179