
MiR-132-3p Regulates the Osteogenic Differentiation of Thoracic Ligamentum Flavum Cells by Inhibiting Multiple Osteogenesis-Related Genes

Abstract
:
1. Introduction
2. Results
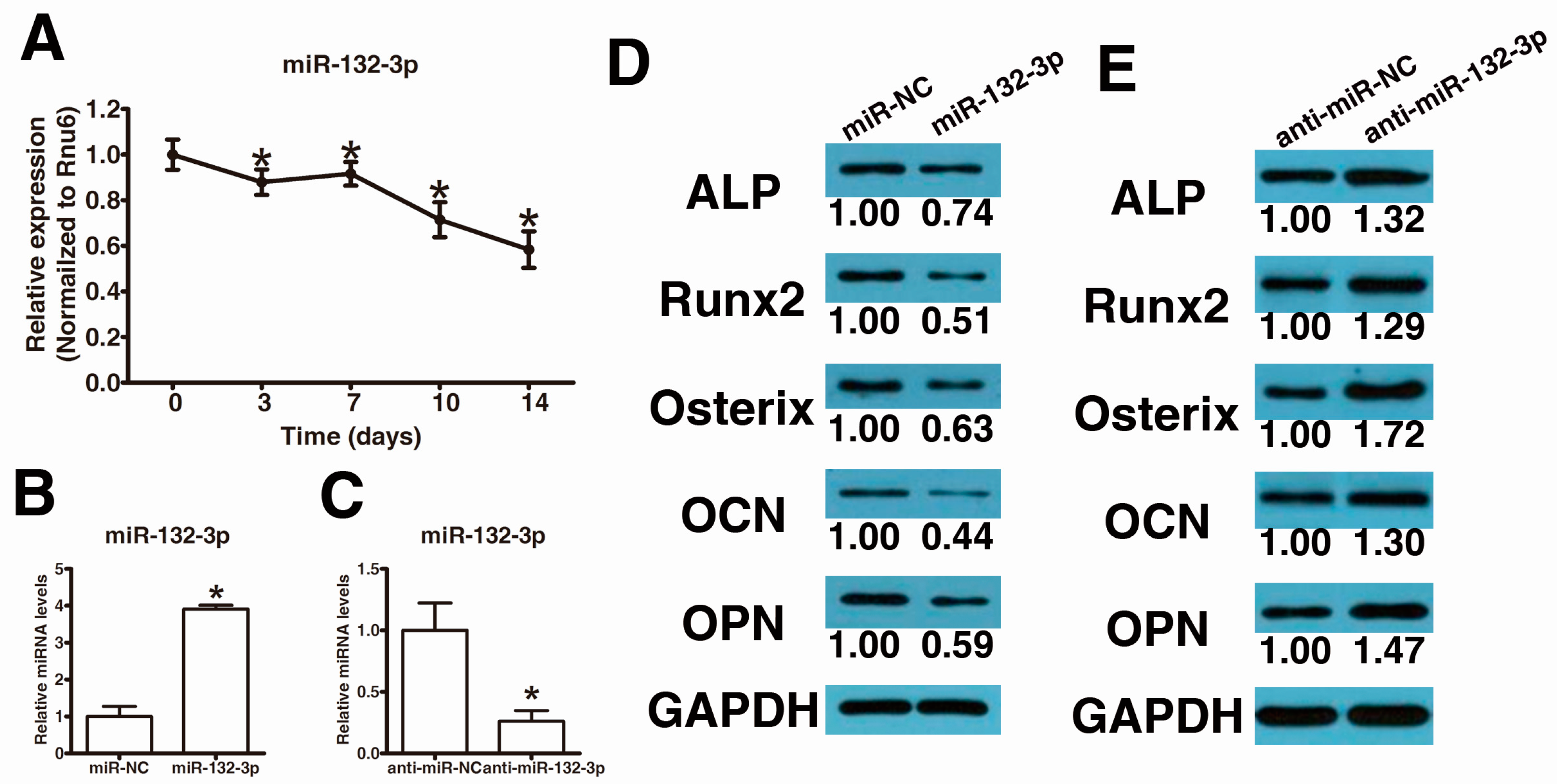
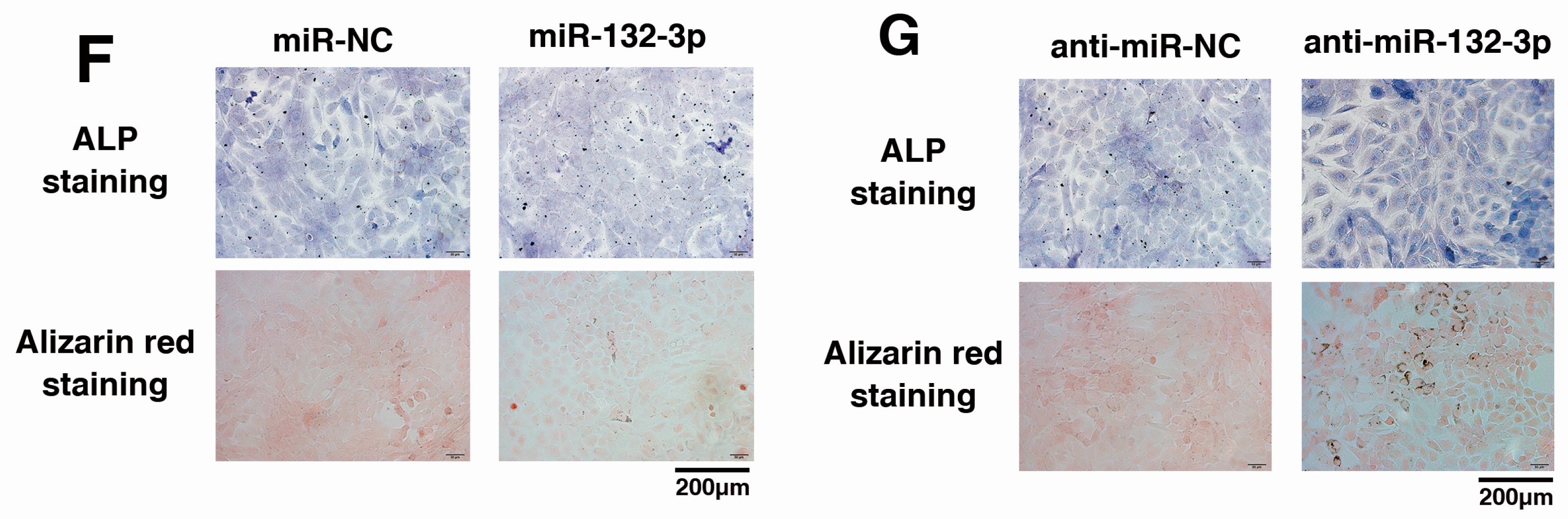
2.1. MiR-132-3p Inhibits the Osteogenic Differentiation of Ligamentum Flavum (LF) Cells
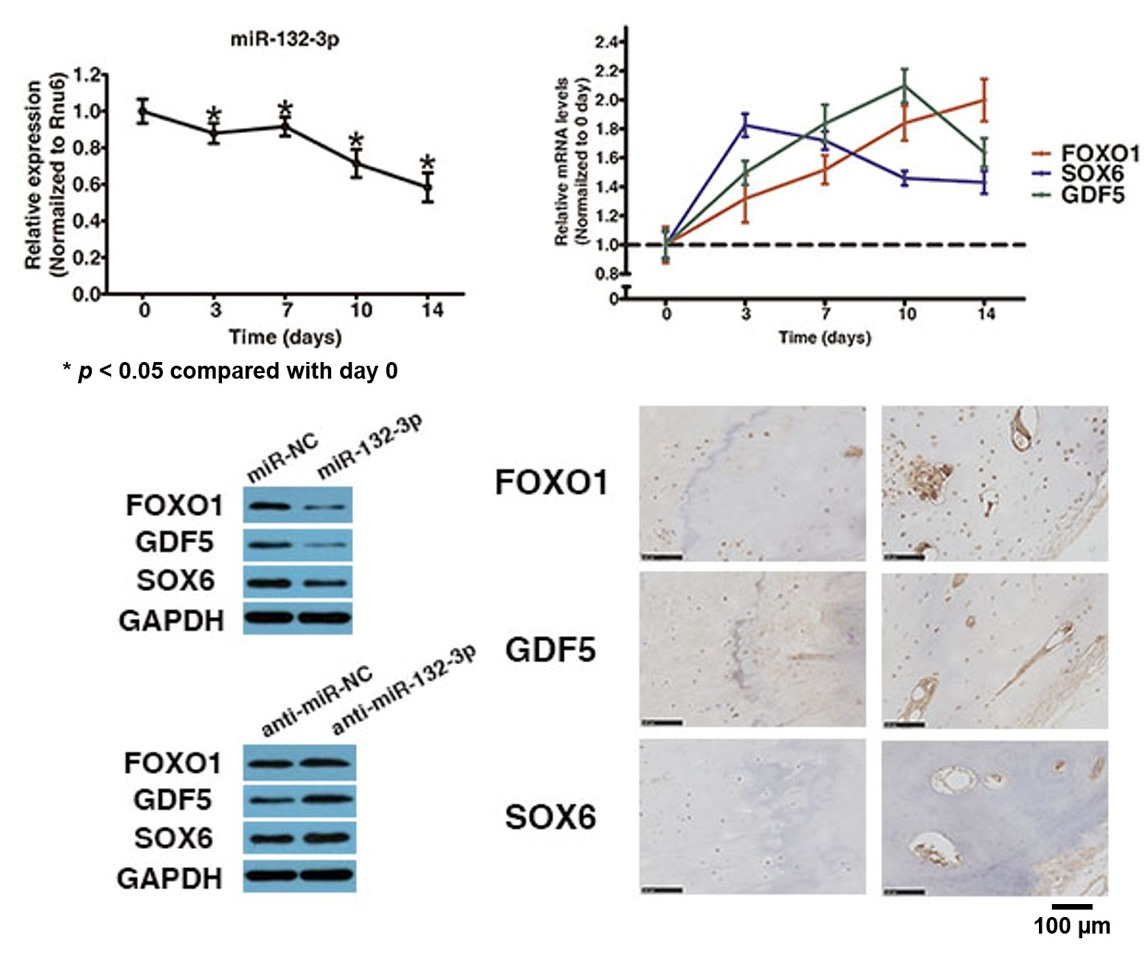
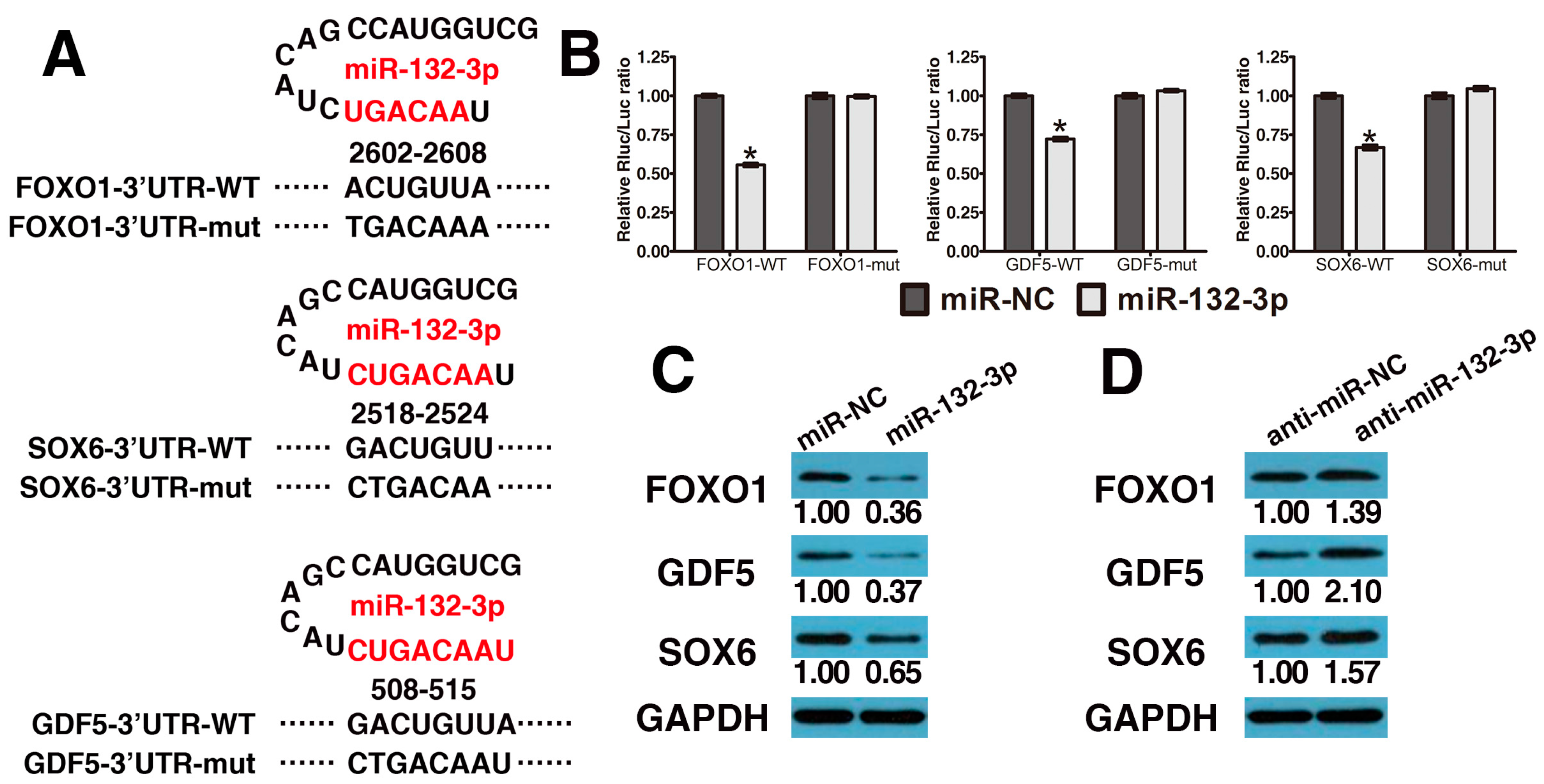
2.2. MiR-132-3p Directly Targets FOXO1, GDF5 and SOX6
2.3. FOXO1, GDF5 and SOX6 Knockdown Inhibits Osteogenic Differentiation in LF Cells
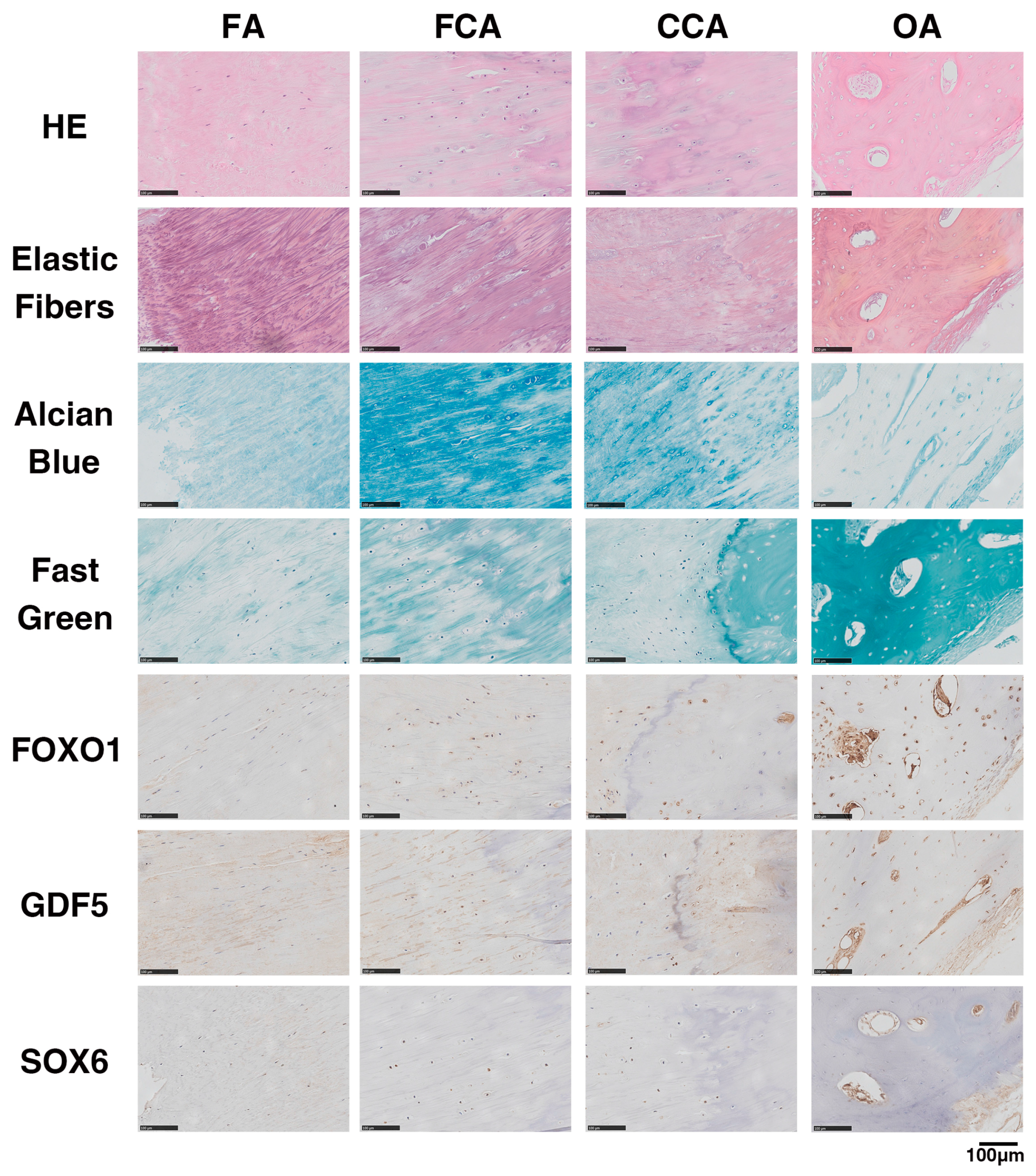
2.4. FOXO1, GDF5 and SOX6 Protein Expression in OLF Samples
3. Discussion
4. Materials and Methods
4.1. Patient Specimens
4.2. Cell Cultures and Osteogenic Differentiation
4.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
4.4. Western Blot Analysis
4.5. Alkaline Phosphatase (ALP) Activity Assay and Alizarin Red Staining
4.6. miRNA/siRNA Transfection
4.7. Luciferase Constructs and Reporter Assay
4.8. Hematoxylin-Eosin (HE) Staining, Tissue-Specific Staining and Immunohistochemical (IHC) Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hou, X.; Sun, C.; Liu, X.; Liu, Z.; Qi, Q.; Guo, Z.; Li, W.; Zeng, Y.; Chen, Z. Clinical features of thoracic spinal stenosis-associated myelopathy: A retrospective analysis of 427 cases. Clin. Spine Surg. 2016, 29, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.B.; Sun, C.G.; Chen, Z.Q. Progress on clinical characteristics and identification of location of thoracic ossification of the ligamentum flavum. Orthop. Surg. 2015, 7, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Chen, Z.; Wang, D.; Guo, Z.; Qiang, Q.; Shang, Y. Osterix is a key target for mechanical signals in human thoracic ligament flavum cells. J. Cell. Physiol. 2007, 211, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, X.; Wang, C.; Yuan, W. Rotational stress: role in development of ossification of posterior longitudinal ligament and ligamentum flavum. Med. Hypotheses 2011, 76, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, C.; He, H.; Xu, G.; Ye, X. Spinal balance failure: A potential cause of spinal ligament ossification. Med. Hypotheses 2011, 76, 908–910. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.X.; Yayama, T.; Uchida, K.; Nakajima, H.; Sugita, D.; Guerrero, A.R.; Yoshida, A.; Baba, H. Cyclic tensile strain facilitates the ossification of ligamentum flavum through β-catenin signaling pathway: In vitro analysis. Spine 2012, 37, E639–E646. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Chen, Z.; Chen, Y.; Shang, Y. Mechanistic roles of leptin in osteogenic stimulation in thoracic ligament flavum cells. J. Biol. Chem. 2007, 282, 29958–29966. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.; Yoon, J.W.; Chung, C.K. Increased bone mineral density in patients with ossification of the ligamentum flavum: A case-control study. J. Clin. Densitom. 2014, 17, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.; Yuan, H.S.; Wang, H.L.; Liao, J.; Li, M.; Guo, F.X.; Shi, S.; Chen, Z.Q. Epidemiological survey of ossification of the ligamentum flavum in thoracic spine: CT imaging observation of 993 cases. Eur. Spine J. 2013, 22, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Ma, X.; Li, F.; Guo, Z.; Qi, Q.; Li, W.; Yuan, H.; Wang, Z.; Chen, Z. COL6A1 polymorphisms associated with ossification of the ligamentum flavum and ossification of the posterior longitudinal ligament. Spine 2007, 32, 2834–2838. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, Y.; Chen, Y.; Shi, G.; Yuan, W. RUNX2 polymorphisms associated with OPLL and OLF in the Han population. Clin. Orthop. Relat. Res. 2010, 468, 3333–3341. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Holmstrom, K.; Qiu, W.; Ditzel, N.; Shi, K.; Hokland, L.; Kassem, M. MicroRNA-34a inhibits osteoblast differentiation and in vivo bone formation of human stromal stem cells. Stem Cells 2014, 32, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Gao, D.; Gao, C.; Wei, P.; Niu, M.; Shuai, C. MicroRNAs regulate signaling pathways in osteogenic differentiation of mesenchymal stem cells. Mol. Med. Rep. 2016, 14, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Deng, Y.; Gu, P.; Fan, X. MicroRNAs regulate bone development and regeneration. Int. J. Mol. Sci. 2015, 16, 8227–8253. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Song, W.; Gao, H.; Li, T.; Cao, X.; Zhong, S.; Wang, Y. miR-29b-loaded gold nanoparticles targeting to the endoplasmic reticulum for synergistic promotion of osteogenic differentiation. ACS Appl. Mater. Interfaces 2016, 8, 19217–19227. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Zhong, L.; Chen, C.; Tang, Z.; Liu, H.; Zhou, Y.; Tang, M.; Zhou, L.; Zuo, G.; Luo, J.; et al. miR-21 synergizes with BMP9 in osteogenic differentiation by activating the BMP9/smad signaling pathway in murine multilineage cells. Int. J. Mol. Med. 2015, 36, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, L.; Zhou, Y.; Ji, X.; Liu, J.; Liu, D.; Yin, P.; Peng, Y.; Hao, M.; Zhang, L.; et al. Synergistic effects of BMP9 and miR-548d-5p on promoting osteogenic differentiation of mesenchymal stem cells. BioMed Res. Int. 2015, 2015, 309747. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wang, S.; Bian, C.; Yang, Z.; Zhou, H.; Zeng, Y.; Li, H.; Han, Q.; Zhao, R.C. Upregulation of miR-22 promotes osteogenic differentiation and inhibits adipogenic differentiation of human adipose tissue-derived mesenchymal stem cells by repressing HDAC6 protein expression. Stem Cells Dev. 2012, 21, 2531–2540. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Ito, H.; Yoshitomi, H.; Yamamoto, K.; Fukuda, A.; Yoshikawa, J.; Furu, M.; Ishikawa, M.; Shibuya, H.; Matsuda, S. Inhibition of miR-92a enhances fracture healing via promoting angiogenesis in a model of stabilized fracture in young mice. J. Bone Miner. Res. 2014, 29, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Guo, B.; Li, Q.; Peng, J.; Yang, Z.; Wang, A.; Li, D.; Hou, Z.; Lv, K.; Kan, G.; et al. miR-214 targets ATF4 to inhibit bone formation. Nat. Med. 2013, 19, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhou, H.; Hong, Y.; Li, J.; Jiang, X.; Huang, H. miR-30 family members negatively regulate osteoblast differentiation. J. Biol. Chem. 2012, 287, 7503–7511. [Google Scholar] [CrossRef] [PubMed]
- Zuo, B.; Zhu, J.; Li, J.; Wang, C.; Zhao, X.; Cai, G.; Li, Z.; Peng, J.; Wang, P.; Shen, C.; et al. microRNA-103a functions as a mechanosensitive microRNA to inhibit bone formation through targeting Runx2. J. Bone Miner. Res. 2015, 30, 330–345. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Jia, L.; Zheng, Y.; Jin, C.; Liu, Y.; Liu, H.; Zhou, Y. miR-34a promotes osteogenic differentiation of human adipose-derived stem cells via the RBP2/notch1/cyclin D1 coregulatory network. Stem Cell Rep. 2016, 7, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Geng, J.; Jiang, S. MicroRNAs in regulation of osteogenic differentiation of mesenchymal stem cells. Cell Tissue Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bicker, S.; Lackinger, M.; Weiss, K.; Schratt, G. MicroRNA-132, -134, and -138: A microRNA troika rules in neuronal dendrites. Cell. Mol. Life Sci. 2014, 71, 3987–4005. [Google Scholar] [CrossRef] [PubMed]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. miR-212/132 expression and functions: Within and beyond the neuronal compartment. Nucleic Acids Res. 2012, 40, 4742–4753. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, Y.; Sun, Z.; Wang, H.; Zhou, H.; Zhang, L.; Zhang, S.; Cao, X. miRNA-132-3p inhibits osteoblast differentiation by targeting Ep300 in simulated microgravity. Sci. Rep. 2015, 5, 18655. [Google Scholar] [CrossRef] [PubMed]
- Gong, K.; Qu, B.; Liao, D.; Liu, D.; Wang, C.; Zhou, J.; Pan, X. miR-132 regulates osteogenic differentiation via downregulating Sirtuin1 in a peroxisome proliferator-activated receptor β/δ-dependent manner. Biochem. Biophys. Res. Commun. 2016, 478, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Cui, X.; Liu, Y.; Zhang, W.; Liu, M.; Sun, Y.E.; Ma, Z.; Gu, X. Positive feedback regulation between microRNA-132 and CREB in spinal cord contributes to bone cancer pain in mice. Eur. J. Pain 2016, 20, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Liu, J.; Wu, Y.; Zhu, Q. MicroRNA-132 inhibits cell growth and metastasis in osteosarcoma cell lines possibly by targeting Sox4. Int. J. Oncol. 2015, 47, 1672–1684. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, G.; Shen, F.; Kang, Y. miR-132 targeting cyclin E1 suppresses cell proliferation in osteosarcoma cells. Tumour Biol. 2014, 35, 4859–4865. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gao, T.; Tang, J.; Cai, H.; Lin, L.; Fu, S. Loss of microRNA-132 predicts poor prognosis in patients with primary osteosarcoma. Mol. Cell. Biochem. 2013, 381, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Rapp, J.; Rainone, S.; Hebert, S.S. MicroRNAs underlying memory deficits in neurodegenerative disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016. [Google Scholar] [CrossRef] [PubMed]
- Tognini, P.; Pizzorusso, T. MicroRNA212/132 family: Molecular transducer of neuronal function and plasticity. Int. J. Biochem. Cell Biol. 2012, 44, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Liu, P.; Yang, S.; Ye, S.; Xu, W.; Liu, X. A three-plasma miRNA signature serves as novel biomarkers for osteosarcoma. Med. Oncol. 2013, 30, 340. [Google Scholar] [CrossRef] [PubMed]
- Gougelet, A.; Pissaloux, D.; Besse, A.; Perez, J.; Duc, A.; Dutour, A.; Blay, J.Y.; Alberti, L. Micro-RNA profiles in osteosarcoma as a predictive tool for ifosfamide response. Int. J. Cancer 2011, 129, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Chen, Y.; Zhang, H.; Chen, Y.; Shen, X.; Shi, C.; Liu, Y.; Yuan, W. Integrated microRNA-mRNA analyses reveal OPLL specific microRNA regulatory network using high-throughput sequencing. Sci. Rep. 2016, 6, 21580. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, J.; Chen, T.; Yin, P.; Yang, J.; Cao, Y. miR-132 upregulation promotes gastric cancer cell growth through suppression of FoxO1 translation. Tumour Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Yayama, T.; Cai, H.X.; Nakajima, H.; Sugita, D.; Guerrero, A.R.; Kobayashi, S.; Yoshida, A.; Chen, K.B.; Baba, H. Ossification process involving the human thoracic ligamentum flavum: Role of transcription factors. Arthritis Res. Ther. 2011, 13, R144. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.M.; Chen, J.T.; Zhang, Y.; Zha, D.S.; Lin, Z.S.; Zhao, C.Y.; Xu, J.C.; Li, T.; Xu, Z.X. Growth/differentiation factor-5 induces osteogenic differentiation of human ligamentum flavum cells through activation of ERK1/2 and p38 MAPK. Cell. Physiol. Biochem. 2010, 26, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Park, J.O.; Lee, B.H.; Kang, Y.M.; Kim, T.H.; Yoon, J.Y.; Kim, H.; Kwon, U.H.; Lee, K.I.; Lee, H.M.; Moon, S.H. Inflammatory cytokines induce fibrosis and ossification of human ligamentum flavum cells. J. Spinal Disord. Tech. 2013, 26, E6–E12. [Google Scholar] [CrossRef] [PubMed]
- Rached, M.T.; Kode, A.; Xu, L.; Yoshikawa, Y.; Paik, J.H.; Depinho, R.A.; Kousteni, S. FoxO1 is a positive regulator of bone formation by favoring protein synthesis and resistance to oxidative stress in osteoblasts. Cell Metab. 2010, 11, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, M.F.; Flowers, S.; Bhattacharya, R.; Faibish, D.; Behl, Y.; Kotton, D.N.; Gerstenfeld, L.; Moran, E.; Graves, D.T. FOXO1 modulates osteoblast differentiation. Bone 2011, 48, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.C.; Liu, Y.; Thant, L.M.; Pang, J.; Palmer, G.; Alikhani, M. Foxo1, a novel regulator of osteoblast differentiation and skeletogenesis. J. Biol. Chem. 2010, 285, 31055–31065. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, X. Growth differentiation factor 5 regulation in bone regeneration. Curr. Pharm. Des. 2013, 19, 3364–3373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, T.L.; Li, X.; Guo, Y. Functional analyses reveal the essential role of SOX6 and RUNX2 in the communication of chondrocyte and osteoblast. Osteoporos. Int. 2015, 26, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Q.; Sun, C.G. Clinical guideline for treatment of symptomatic thoracic spinal stenosis. Orthop. Surg. 2015, 7, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, C.; Liu, X.; Liu, Z.; Qi, Q.; Guo, Z.; Leng, H.; Chen, Z. The frequency and treatment of dural tears and cerebrospinal fluid leakage in 266 patients with thoracic myelopathy caused by ossification of the ligamentum flavum. Spine 2012, 37, E702–E707. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Chen, Z.; Fan, D.; Sun, C.; Zeng, Y.; Hou, X.; Ning, S. Notch signaling pathways in human thoracic ossification of the ligamentum flavum. J. Orthop. Res. 2016. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer (5′–3′) |
|---|---|
| FOXO1 | Fw: AAGCTCCCAAGTGACTTGGATG Rv: CTGCTCACTAACCCTCAGCCTGA |
| GDF5 | Fw: AAAAGGACAGCTTCCCGGAG Rv: GCCTCCCTTTCTGTCAGCAT |
| SOX6 | Fw: TCAACATGTGGCCTCCCATC Rv: GATGACAGAACGCTGTCCCA |
| GAPDH | Fw: TCAAGGCTGAGAACGGGAAG Rv: TGGACTCCACGACGTACTCA |
| miR-132-3p | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCGACCATG Fw: GCGCGCGTAACAGTCTACAGC Rv: GTCGTATCCAGTGCAGGGTCC |
| U6 | RT: AAAATATGGAACGCTTCACGAATTTG Fw: CTCGCTTCGGCAGCACATATACT Rv: CGCTTCACGAATTTGCGTGT |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, X.; Chen, Z.; Fan, D.; Sun, C.; Zeng, Y. MiR-132-3p Regulates the Osteogenic Differentiation of Thoracic Ligamentum Flavum Cells by Inhibiting Multiple Osteogenesis-Related Genes. Int. J. Mol. Sci. 2016, 17, 1370. https://doi.org/10.3390/ijms17081370
Qu X, Chen Z, Fan D, Sun C, Zeng Y. MiR-132-3p Regulates the Osteogenic Differentiation of Thoracic Ligamentum Flavum Cells by Inhibiting Multiple Osteogenesis-Related Genes. International Journal of Molecular Sciences. 2016; 17(8):1370. https://doi.org/10.3390/ijms17081370
Chicago/Turabian StyleQu, Xiaochen, Zhongqiang Chen, Dongwei Fan, Chuiguo Sun, and Yan Zeng. 2016. "MiR-132-3p Regulates the Osteogenic Differentiation of Thoracic Ligamentum Flavum Cells by Inhibiting Multiple Osteogenesis-Related Genes" International Journal of Molecular Sciences 17, no. 8: 1370. https://doi.org/10.3390/ijms17081370
APA StyleQu, X., Chen, Z., Fan, D., Sun, C., & Zeng, Y. (2016). MiR-132-3p Regulates the Osteogenic Differentiation of Thoracic Ligamentum Flavum Cells by Inhibiting Multiple Osteogenesis-Related Genes. International Journal of Molecular Sciences, 17(8), 1370. https://doi.org/10.3390/ijms17081370