1. Introduction
Red palm weevil (RPW) is an exotic highly destructive pest of palms, particularly
Phoenix dactylifera, in different geographical areas worldwide. The legless, creamy white larvae (grubs) of
Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) are the most destructive stage of the weevil. These insects feed on tender soft palm tissues and move toward the centre of the infested host palm. Under severe attack, such a feeding pattern causes the crown to collapse [
1,
2].
The control of the RPW primarily relies on the frequent use of synthetic pesticides [
1]. These practices are not sustainable, endanger biological diversity, and deteriorate environmental quality. These setbacks have led to the search for alternative methods of RPW control, particularly bio-control agents. Previous studies have reported the use of bacteria [
3], fungi [
4,
5,
6], nematodes [
7], and parasitoids [
8] against RPW. However, the use of entomopathogenic fungi, is a promising alternative for reversing the hazardous dependence of agriculture on synthetic insecticides. The initial step toward the development of mycoinsecticides should involve laboratory evaluation to grade the tested isolates in terms of virulence [
9].
The virulence of entomopathogenic fungal isolates primarily depends on a series of complex factors, including conidial hydrophobicity, germination, polarity, and fungal hydrolytic enzyme activities. The conidial hydrophobicity of entomopathogenic fungal isolates varies with the developmental stage of the fungus. Previous studies have characterized the cysteine-rich hydrophobin genes from
Metarhizium anisopliae and
Beauveria bassiana. Furthermore, these findings provided evidence of the involvement of these genes in the adhesion and virulence of entomopathogenic fungal isolates [
10].
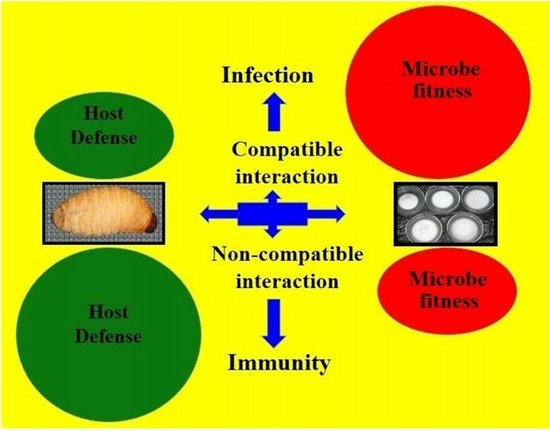
Host immune defence mechanisms are triggered immediately after entomopathogenic fungal spores invade and compromise the integrity of the host cuticle. Subsequently, a struggle between the host and pathogen is initiated, potentially leading to infection (compatible interaction) or disease resistance (non-compatible interaction). This host immune defence primarily involves different innate immune reactions, such as cellular and humoral immune responses [
11]. Thus, the host cuticle covering the entire body provides the first line of immune defence against invading pathogens. The pathogens breach the cuticle through the production of proteases [
12,
13]. In addition, insects display cellular immune responses, such as self-defence. However, the invading pathogens release toxins (e.g., destruxins) to reduce the impact of host immune responses. Once the pathogen has overcome the physical barriers, the host membranes produce molecular structures that secrete proteins involved in host defence to cope with the pathogen attack [
14]. The fat body is a major immune responsive organ involved in the synthesis of unique antimicrobial peptides (AMPs) targeting specific microorganisms. There have been a few scattered reports on the immune responses of red palm weevils induced through nematodes [
15] and bacteria [
16,
17,
18]. Currently, there are no studies concerning the molecular mechanisms of the RPW immune response induced through entomopathogenic fungi. Thus, the present study was designed to evaluate the potential of entomopathogenic fungal isolates, determine the effect of the tested fungal isolates on the growth and development of
R. ferrugineus larvae, and gain insights into the molecular mechanisms of the red palm weevil immune response against invading entomopathogenic fungal microbes to improve
R. ferrugineus control strategies.
3. Discussion
The pathogenic and growth inhibition activities of naturally-occurring entomopathogenic fungi against insects of different orders have previously been reported [
19]. Previous studies have given little consideration to the virulence-determining traits of entomopathogenic fungi and their impact on the growth and immune defence mechanism of an economically important pest of the date palm,
R. ferrugineus. The present study showed that microbe virulence factors, including the germination percentage and relative conidial hydrophobicity, significantly affect the virulence of the studied isolates. Entomopathogenic fungal isolates with higher virulence, elicit higher expression of immune related genes, and slowing the larval growth, ultimately leading to higher mortality among infected larvae.
The current findings clearly indicated that conidial germination and hydrophobicity defined the virulence of the isolates. We found variations in germination and conidial hydrophobicity among the studied isolates. Conidial hydrophobicity is specifically provided through a group of small cysteine-rich proteins (hydrophobins), involved in conidial adhesion, virulence, and dispersion. Recently, an empirical evidence of the involvement of three hydrophobin genes
Hyd1,
Hyd2 and
Hyd3 in virulence, hydrophobicity, and conidiation was provided [
20]. In another study, the hydrophobin genes,
Hyd1 and
Hyd2, were implicated in conidial hydrophobicity and, ultimately, the regulation of the level of adhesion and virulence of the strain [
21,
22]. Based on these findings, we suggested that pathogen virulence is important to invade the host. Conidia with high levels of hydrophobicity resulted in the high mortality of red palm weevil larvae. During fungal pathogen selection, virulence-determining traits, such as conidial hydrophobicity and conidial germination, reported in the current study should be thoroughly considered.
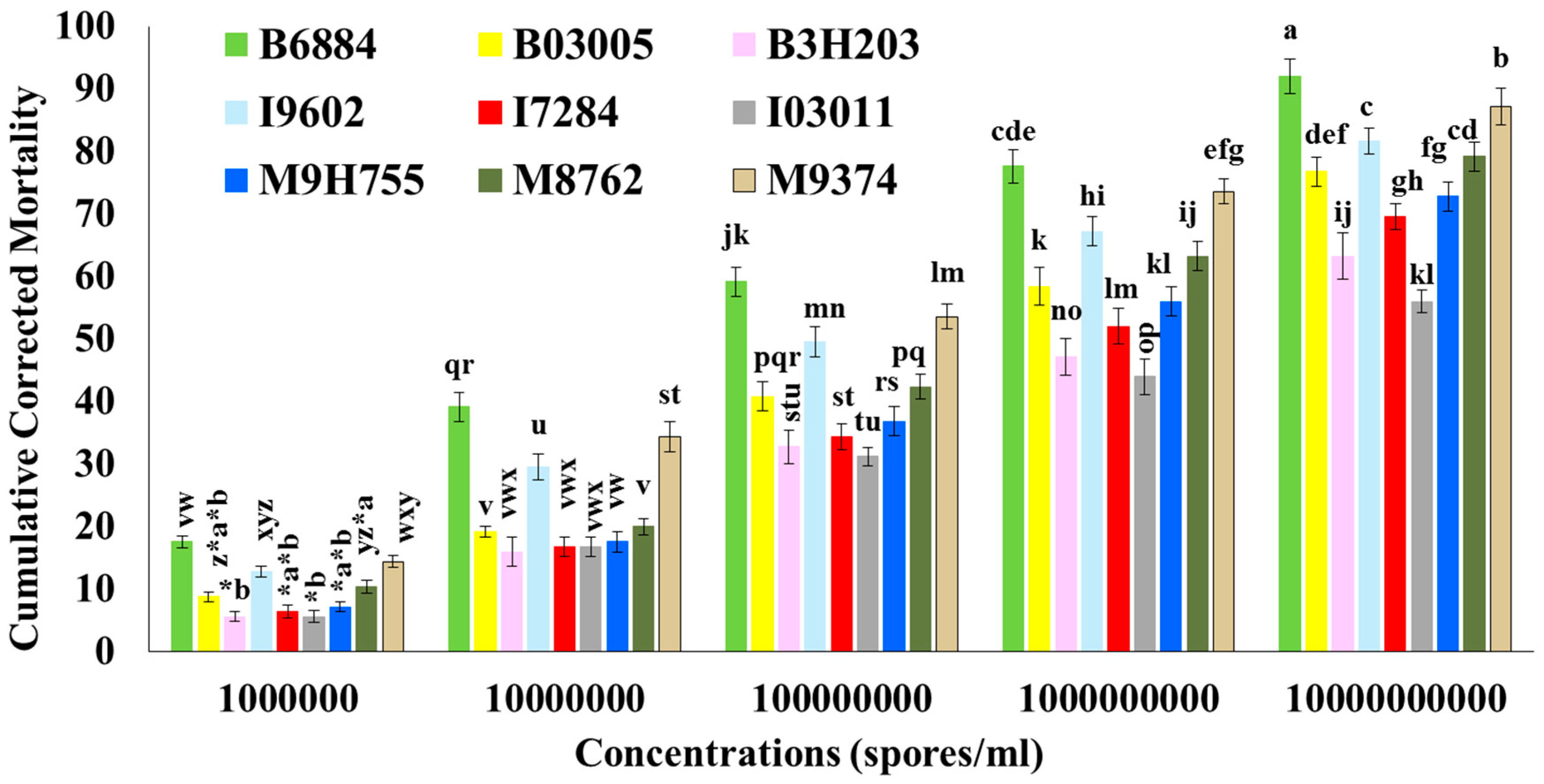
R. ferrugineus larvae infected with isolates B6884 and M9374 gained less weight compared with uninfected (control) larvae. Isolate B6884 remained the most virulent isolate, followed by isolates M9374 > I9602 > M8762 > B03005 > M9H755 > I7284 > B3H203 > I03011. Nutritional indices, particularly the ECI and ECD values of
R. ferrugineus larvae, were significantly different, suggesting that virulent isolates tremendously reduced the growth of red palm weevil larvae. The ECI values representing the overall measurement of ingested food used by the larvae for growth were greatly reduced by the infection of isolate B6884 (46.41%), M9374 (36.59%), I9602 (29.25%), M8762 (20.00%), B03005 (14.88%), M9H755 (8.74%), I7284 (4.22%), B3H203 (2.67%), and I03011 (0.29%). The decrease in ECI values compared with the control revealed that most of the ingested food is used for energy to combat the invading pathogen, and less food is used for larval growth. These findings are consistent with a previous investigation in which
M. anisopliae (strain 406) significantly reduced the ECI values against all of the studied larval instars of
Ocinara varians Walker [
23].
ECD, an index to calculate the precise insect biomass from digested food, was also decreased upon infection with the conidia of virulent isolates. This reduction might reflect the fact that digested food is metabolized for energy production, thereby reducing the ECD compared with the control [
5]. In another study, similar growth inhibition pattern of ECI and ECD was reported in
O. varians Walker larvae in studies of the virulence of entomopathogenic fungal strains [
9].
The fungal infection of virulent isolates tremendously increased the AD values of the red palm weevil larvae. The enhanced AD values might reflect the fact that the fungus-inoculated nutrient-deficient larvae require energy for immune defence. This energy could only be obtained through the use of intrinsic abilities to enhance the approximate digestibilities of the limited foodstuff. Similar results were obtained when the pesticidal activity of labramin was evaluated against the Mediterranean flour moth [
24].
It is clear from the mortality and nutritional indices results that isolates B6884 and M9374 are the most virulent. This virulence is mainly regulated by the conidial hydrophobicity that enhances the infection by facilitating conidial adhesion with the host cuticle. In case of compatible interaction (infection), the host immune defence mechanism is greatly up-regulated to combat infection and less energy remained for host growth. The host, under such circumstances (low energy), becomes weaker and more vulnerable to the sporulating conidia that starts their growth in vivo.
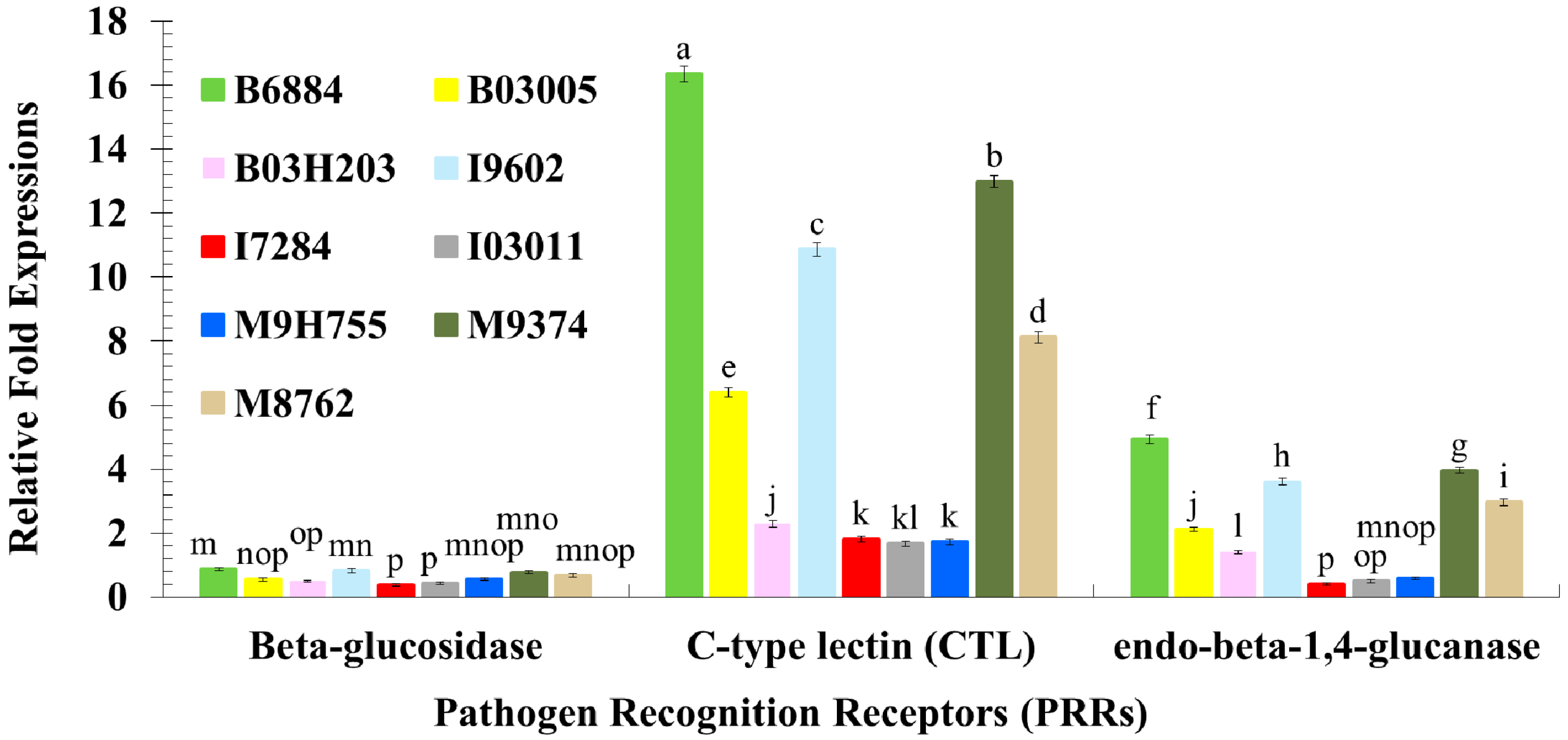
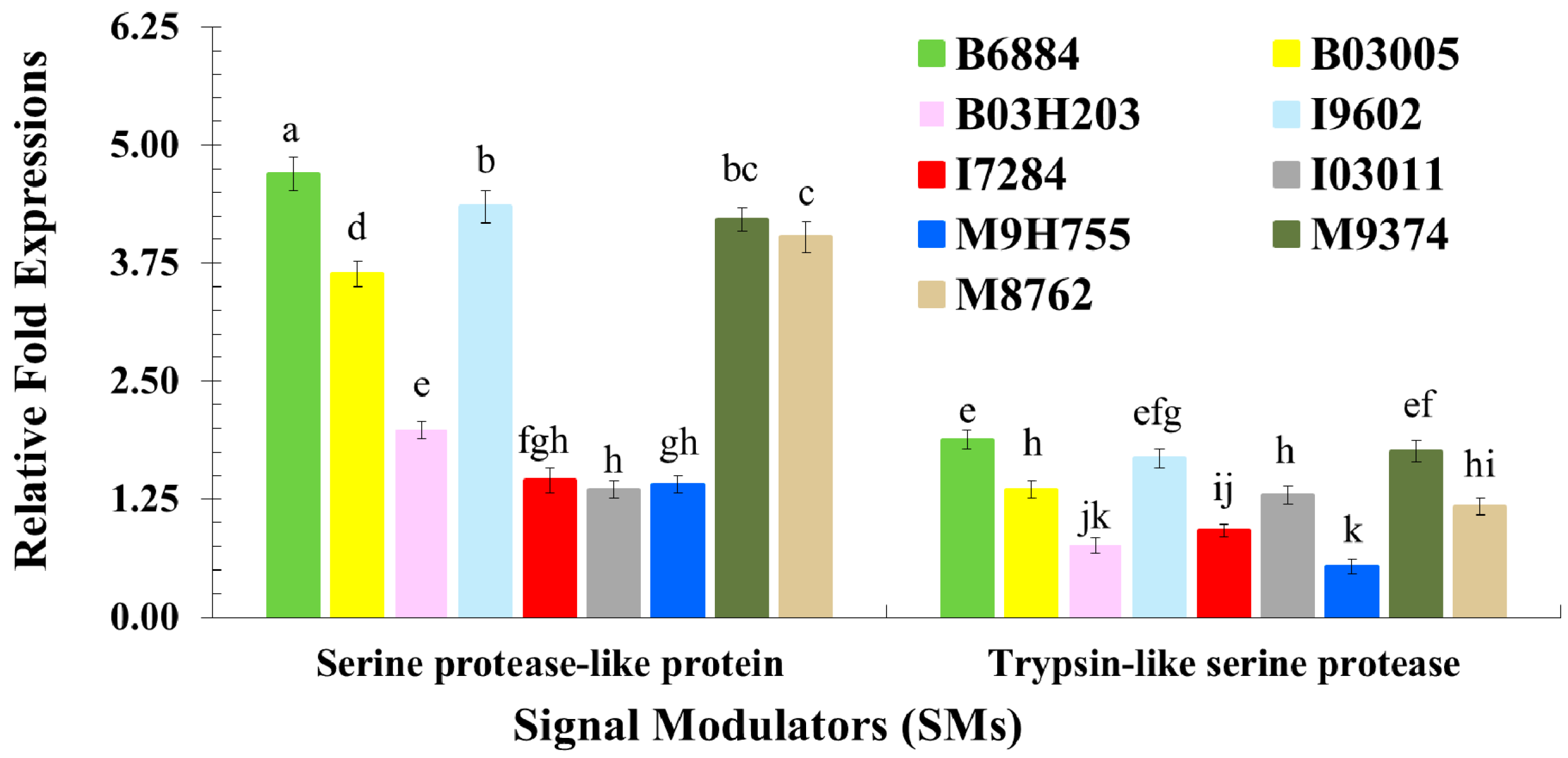
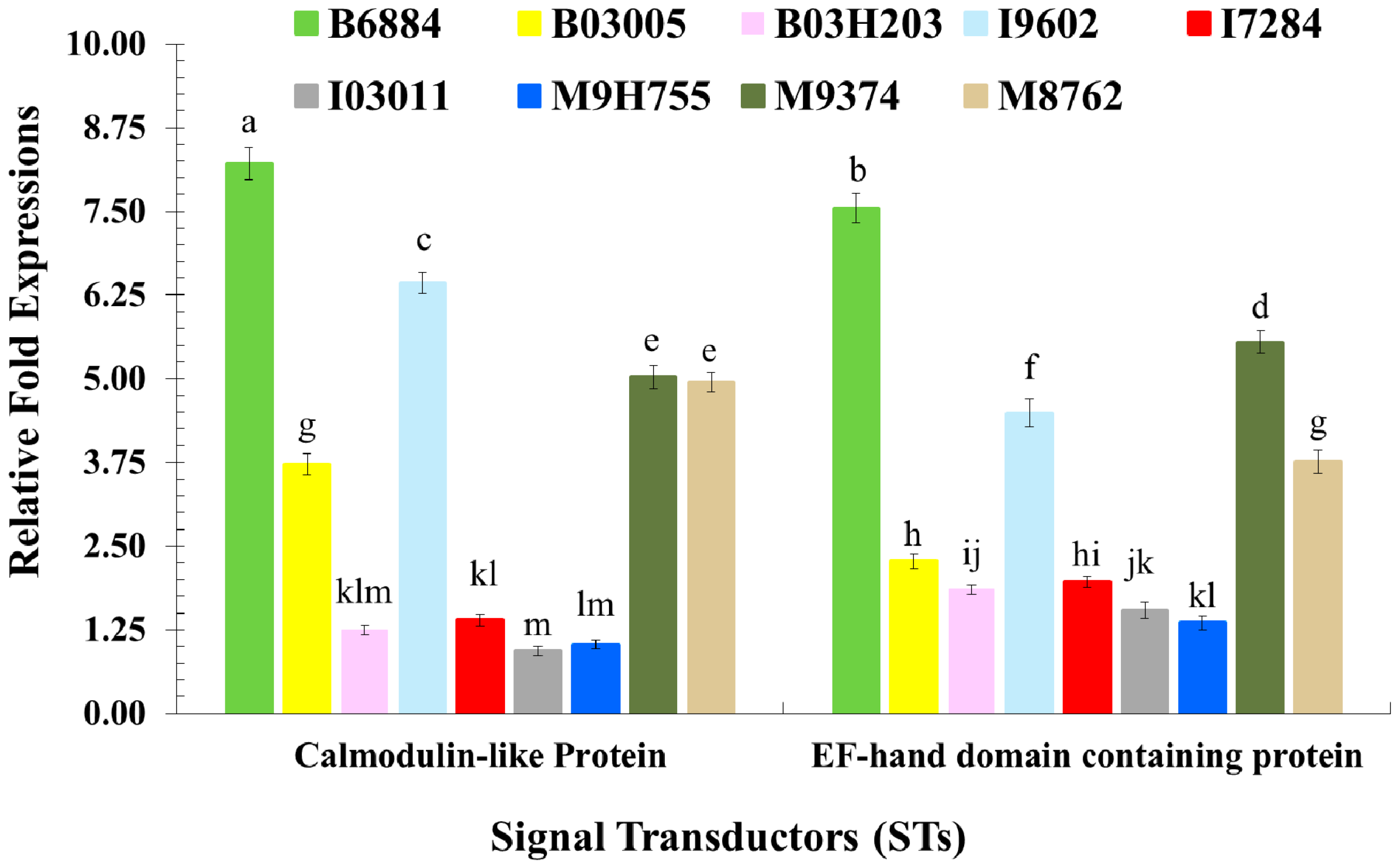
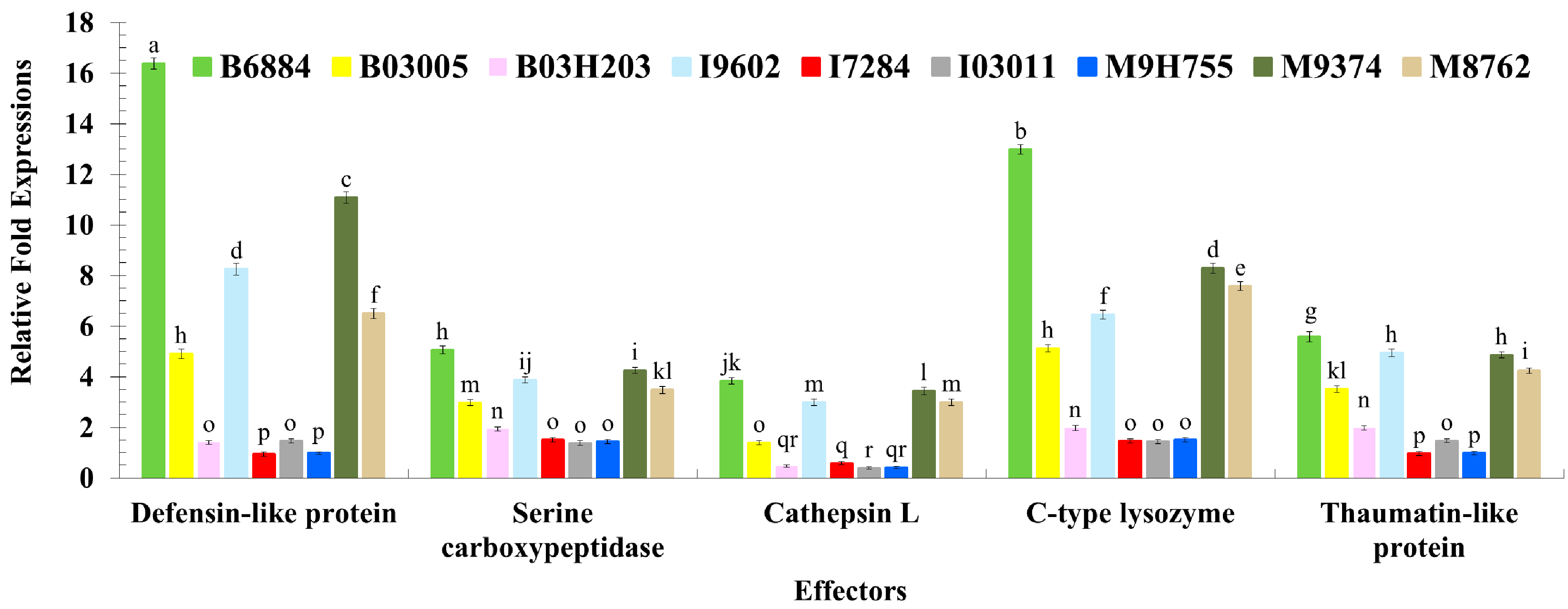
These findings also showed that isolates B6884 and M9374 enhanced the response of the pathogen recognition receptors, signal modulators, signal transductors and effectors responsible for host immune defence mechanisms. The enhanced activities of PRRs (particularly C-type lectin and endo-beta-1,4-glucanse); signal modulators (particularly Serine protease-like protein); signal transductors (Calmodulin-like protein and EF-hand domain containing protein); and effectors (C-type lysozyme, Cathepsin L., Defensin-like protein, Serine carboxypeptidase, and Thaumatin-like protein) reduced ECI and ECD values, revealing that the host spends most of its energy combating the invading pathogen attack; hence, little nutrition remains for growth and development. The strains with less conidial hydrophobicity and low germination remained less pathogenic and should not be incorporated into the integrated red palm weevil management programme.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}