
Expression of Transglutaminase in Foreskin of Children with Balanitis Xerotica Obliterans

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
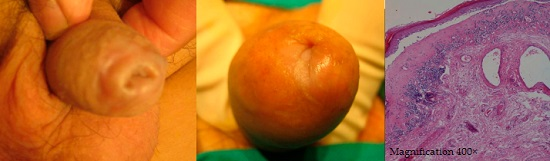
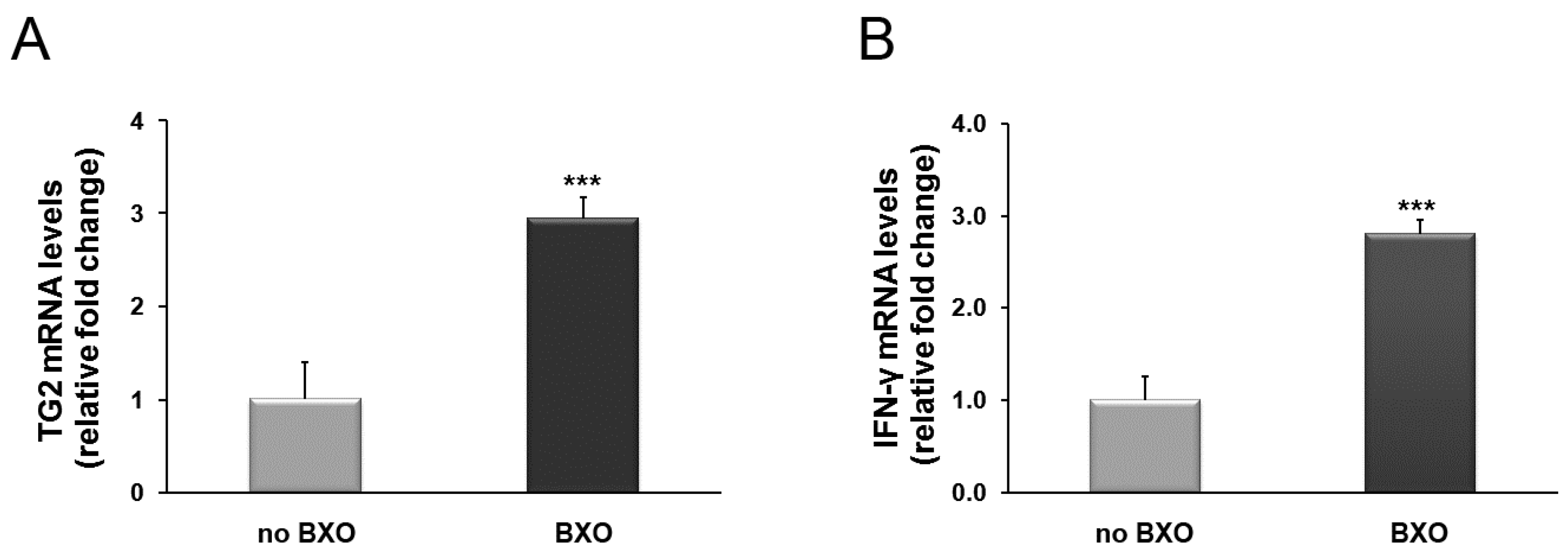
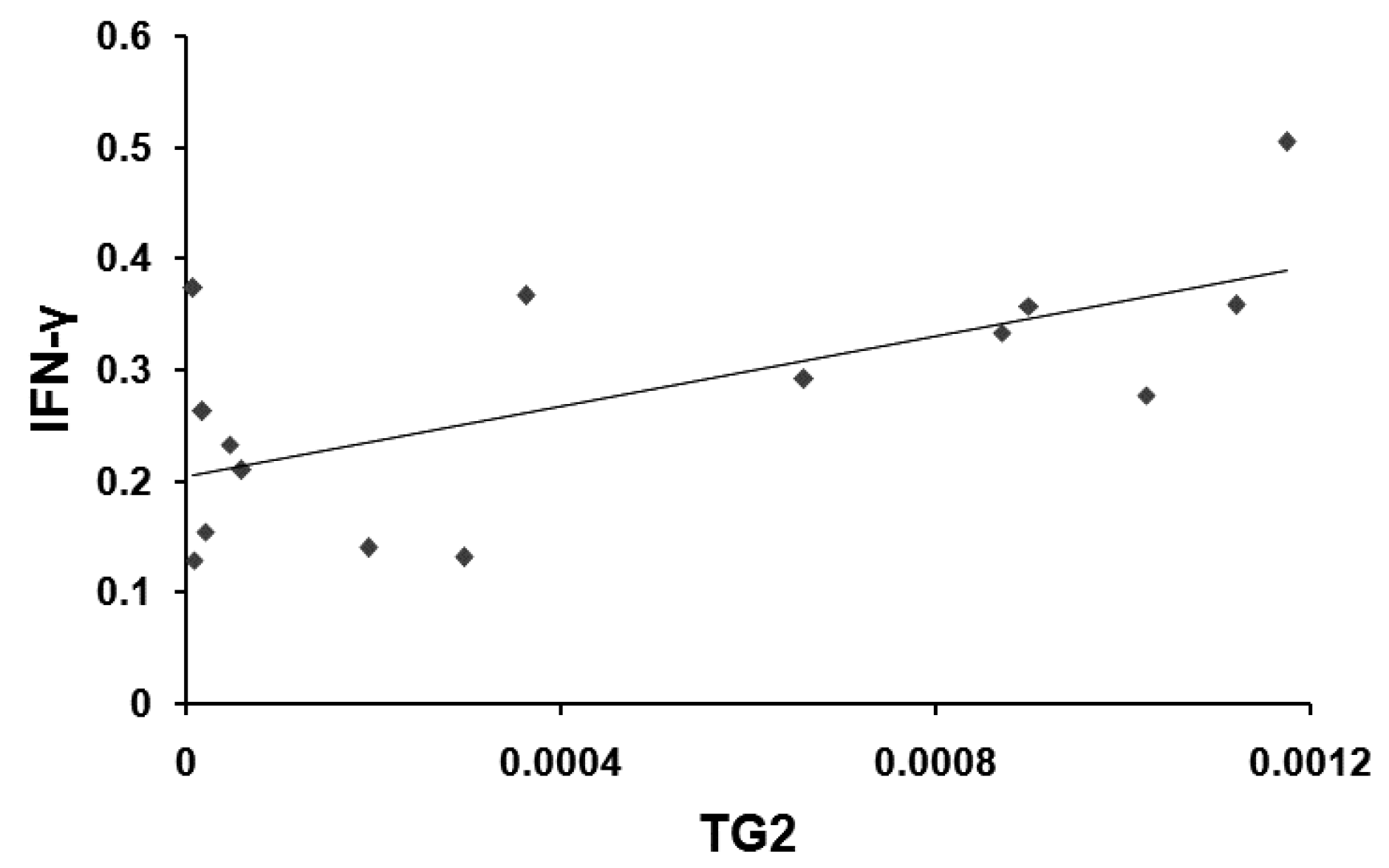
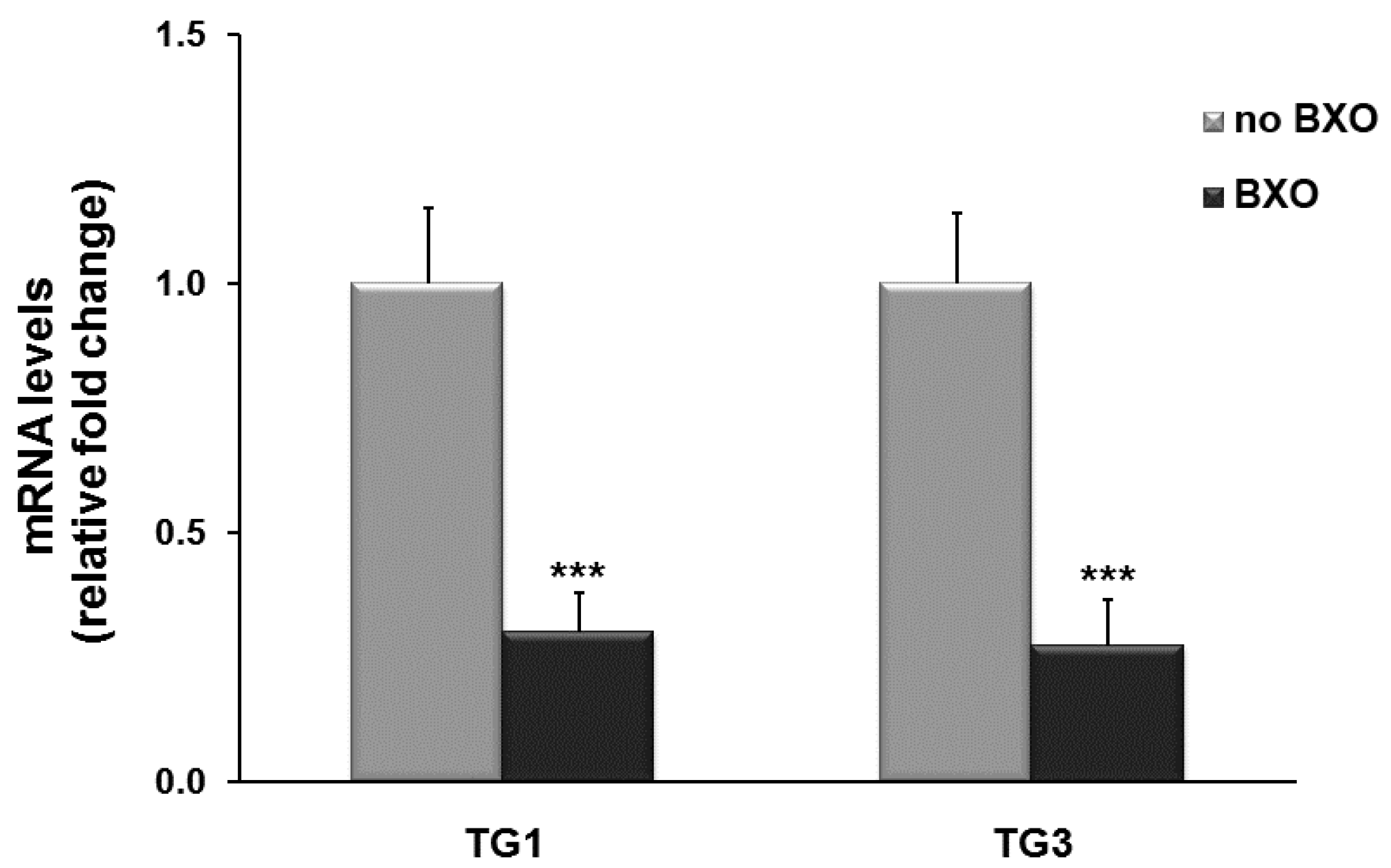
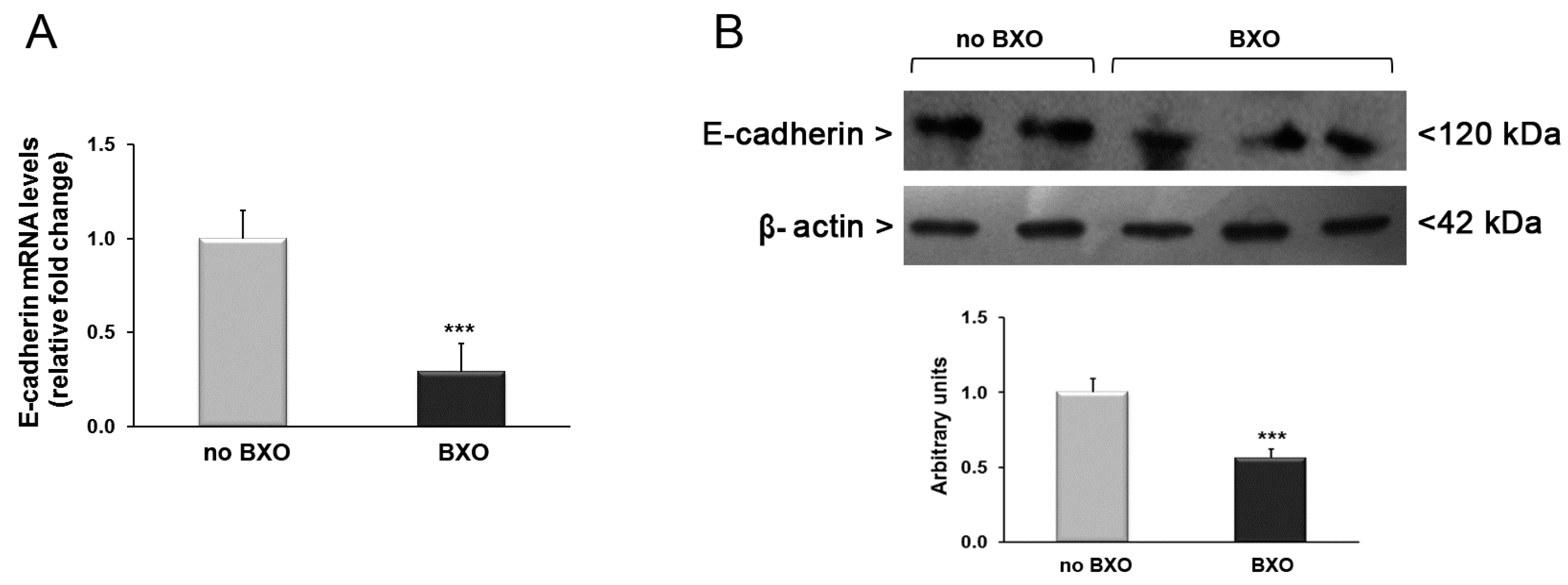
2. Results
3. Discussion
4. Material and Methods
4.1. Patient Recruitment
4.2. Gene Expression Analysis
4.3. Western Blotting
4.4. Statistical Analysis
Author Contributions
Conflicts of Interest
References
- Kizer, W.S.; Prarie, T.; Morey, A.F. Balanitis xerotica obliterans: Epidemiologic distribution in an equal access health care system. South Med. J. 2003, 96, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Bochove-Overgaauw, D.M.; Gelders, W.; de Vylder, A.M. Routine biopsies in pediatric circumcision: (Non) sense? J. Pediatr. Urol. 2009, 5, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.C.; Kirtschig, G.; Baldo, M.; Brackenbury, F.; Lewis, F.; Wojnarowska, F. Topical interventions for genital lichen sclerosus. Cochrane Database Syst. Rev. 2011, 12, CD008240. [Google Scholar] [PubMed]
- Becker, K.; Meissner, V.; Farwick, W.; Bauer, R.; Gaiser, M.R. Lichen sclerosus and atopy in boys: Coincidence or correlation? Br. J. Dermatol. 2013, 168, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, S.; Antao, B.; Bevington, O.; Furness, P.; Ninan, G.K. Balanitis Xerotica Obliterans in children and its incidence under the age of 5 years. J. Pediatr. Urol. 2012, 8, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.M.; Dean, D.; Millard, P.R.; Charnock, F.M.; Wojnarowska, F. Cytokine alterations in lichen sclerosus: An immunohistochemical study. Br. J. Dermatol. 2006, 155, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Nakaoka, H.; Perez, D.M.; Baek, K.J.; Das, T.; Husain, A.; Misono, K.; Im, M.J.; Graham, R.M. G(h): A GTP-binding protein with transglutaminase activity and receptor signaling function. Science 1994, 264, 1593–1596. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, G.; Suwa, M.; Ichikawa, Y.; Ohtsuka, T.M.; Kumagai, S.; Kikuchi, M.; Sato, Y.; Saito, Y.G. A novel function of tissue-type transglutaminase: Protein disulphide isomerise. Biochem. J. 2003, 373, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Murphy, L.J. Tissue transglutaminase has intrinsic kinase activity. Identification of transglutaminase 2 as an insulin-like growth factor-binding protein-3 kinase. J. Biol. Chem. 2004, 279, 23863–23868. [Google Scholar] [CrossRef] [PubMed]
- Belkin, A.M. Extracellular TG2: Emerging functions and regulation. FEBS J. 2011, 278, 4704–4716. [Google Scholar] [CrossRef] [PubMed]
- Nadalutti, C.; Viiri, K.M.; Kaukinen, K.; Mäki, M.; Lindfors, K. Extracellular transglutaminase 2 has a role in cell adhesion, whereas intracellular transglutaminase 2 is involved in regulation of endothelial cell proliferation and apoptosis. Cell Prolif. 2011, 44, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Zemskov, E.A.; Janiak, A.; Hang, J.; Waghray, A.; Belkin, A.M. The role of tissue transglutaminase in cell-matrix interactions. Front. Biosci. 2006, 11, 1057–1076. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.F.; Tatsukawa, H.; Kojima, S. New insights into the functions and localization of nuclear transglutaminase 2. FEBS J. 2011, 278, 4756–4767. [Google Scholar] [CrossRef] [PubMed]
- Verderio, E.A.; Johnson, T.; Griffin, M. Tissue transglutaminase in normal and abnormal wound healing: Review article. Amino Acids 2004, 26, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Ientile, R.; Caccamo, D.; Griffin, M. Tissue transglutaminase and the stress response. Amino Acids 2007, 33, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Iismaa, S.E.; Mearns, B.M.; Lorand, L.; Graham, R.M. Transglutaminases and disease: Lessons from genetically engineered mouse modelsand inherited disorders. Physiol. Rev. 2009, 89, 991–1023. [Google Scholar] [CrossRef] [PubMed]
- Kalinin, A.; Marekov, L.N.; Steinert, P.M. Assembly of the epidermal cornified cell envelope. J. Cell Sci. 2001, 114, 3069–3070. [Google Scholar] [PubMed]
- Zhang, Z.; Xing, J.; Ma, L.; Gong, R.; Chin, Y.E.; Zhuang, S. Transglutaminase-1 regulates renal epithelial cell proliferation through activation of Stat-3. J. Biol. Chem. 2009, 284, 3345–3353. [Google Scholar] [CrossRef] [PubMed]
- Steinert, P.M.; Marekov, L.N. Direct evidence that involucrin is a major early isopeptide cross-linked component of the keratinocyte cornifed cell envelope. J. Biol. Chem. 1997, 272, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Egberts, F.; Heinrich, M.; Jensen, J.M.; Winoto-Morbach, S.; Pfeiffer, S.; Wickel, M.; Schunck, M.; Steude, J.; Saftig, P.; Proksch, E.; et al. Cathepsin D is involved in the regulation of transglutaminase 1 and epidermal differentiation. J. Cell Sci. 2004, 117, 2295–2307. [Google Scholar] [CrossRef] [PubMed]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, K.; Horio, Y.; Ikura, K.; Yamanishi, K.; Maki, M. Analysis of epidermal-type transglutaminase (TGase 3) expression in mouse tissues and cell lines. Int. J. Biochem. Cell Biol. 2001, 33, 491–498. [Google Scholar] [CrossRef]
- Sardy, M.; Karpati, S.; Merkl, B.; Paulsson, M.; Smyth, N. Epidermal transglutaminase (TGase 3) is the autoantigen of dermatitis herpetiformis. J. Exp. Med. 2002, 195, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Celis, S.; Reed, F.; Murphy, F.; Adams, S.; Gillick, J.; Abdelhafeez, A.H.; Lopez, P.J. Balanitis xerotica obliterans in children and adolescents: A literature review and clinical series. J. Pediatr. Urol. 2014, 10, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Bale, P.; Lochhead, A.; Martin, H.C.; Gollow, I. BXO in children. Pediatr. Pathol. 1987, 7, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.; Király, L.; Kutasy, B.; Merksz, M. High incidence of Balanitis Xerotica Obliterans in boys with phimosis: Prospective 10-year study. Pediatr. Dermatol. 2005, 22, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Gambichler, T.; Skrygan, M.; Czempiel, V.; Tigges, C.; Kobus, S.; Meier, J.J.; Köhler, C.U.; Scola, N.; Stücker, M.; Altmeyer, P.; et al. Differential expression of connective tissue growth factor and extracellular matrix proteins in lichen sclerosus. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Terlou, A.; Santegoets, L.A.; van der Meijden, W.I.; Heijmans-Antonissen, C.; Swagemakers, S.M.; van der Spek, P.J.; Ewing, P.C.; van Beurden, M.; Helmerhorst, T.J.; Blok, L.J. An autoimmune phenotype in vulvar lichen sclerosus and lichen planus: A Th1 response and high levels of microRNA-155. J. Investig. Dermatol. 2012, 132, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, E.; Barton, G.; Buisson, S.; Francis, N.; Gotch, F.; Game, L.; Haddad, M.; Dinneen, M.; Bunker, C. Gene expression profiling in male genital lichen sclerosus. Int. J. Exp. Pathol. 2011, 92, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Pilatz, A.; Altinkilic, B.; Schormann, E.; Maegel, L.; Izykowski, N.; Becker, J.; Weidner, W.; Kreipe, H.; Jonigk, D. Congenital phimosis in patients with and without lichen sclerosus: Distinct expression patterns of tissue remodeling associated genes. J. Urol. 2013, 189, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Carli, P.; Cattane, A.; Pimpinelli, N.; Cozza, A.; Bracco, G.; Giannotti, B. Immunohistochemical evidence of skin immune system involvement in vulvar lichen sclerosus et atrophicus. Dermatologica 1991, 182, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Fayyazi, A.; Schweyer, S.; Soruri, A.; Duong, L.Q.; Radzun, H.J.; Peters, J.; Parwaresch, R.; Berger, H. T lymphocytes and altered keratinocytes express interferon-γ and interleukin 6 in lichen planus. Arch. Dermatol. Res. 1999, 291, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Mehta, K.; Kumar, A.; Kim, H.I. Transglutaminase 2: A multi-tasking protein in the complex circuitry of inflammation and cancer. Biochem. Pharmacol. 2010, 80, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Diraimondo, T.R.; Klöck, C.; Khosla, C. Interferon-γ Activates Transglutaminase 2 via a phosphatidylinositol-3-kinase-dependent pathway: Implications for celiac sprue therapy. J. Pharmacol. Exp. Ther. 2012, 341, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Condello, S.; Caccamo, D.; Currò, M.; Ferlazzo, N.; Parisi, G.; Ientile, R. Transglutaminase 2 and NF-κB interplay during NGF-induced differentiation of neuroblastoma cells. Brain Res. 2008, 1207, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mehta, K. Tissue transglutaminase constitutively activates HIF-1α promoter and nuclearfactor-κB via a non-canonical pathway. PLoS ONE 2012, 7, e49321. [Google Scholar] [CrossRef] [PubMed]
- Yakubov, B.; Chelladurai, B.; Schmitt, J.; Emerson, R.; Turchi, J.J.; Matei, D. Extracellular tissue transglutaminase activates noncanonical NF-κB signaling and promotes metastasis in ovarian cancer. Neoplasia 2013, 15, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Matarese, G.; Currò, M.; Isola, G.; Caccamo, D.; Vecchio, M.; Giunta, M.L.; Ramaglia, L.; Cordasco, G.; Williams, R.C.; Ientile, R. Transglutaminase 2 up-regulation is associated with RANKL/OPG pathway in cultured HPDL cells and THP-1-differentiatedmacrophages. Amino Acids 2015, 47, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Telci, D.; Griffin, M. Tissue transglutaminase (TG2)—A wound response enzyme. Front. Biosci. 2006, 11, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Smith, L.L.; Wynne, J. Changes in transglutaminase activity in an experimental model of pulmonary fibrosis induced by paraquat. Br. J. Exp. Pathol. 1979, 60, 653–661. [Google Scholar] [PubMed]
- Mirza, A.; Liu, S.L.; Frizell, E.; Zhu, J.; Maddukuri, S.; Martinez, J.; Davies, P.; Schwarting, R.; Norton, P.; Zern, M.A. A role for tissue transglutaminase in hepatic injury and fibrogenesis, and its regulation by NF-kappaB. Am. J. Physiol. 1997, 272, G281–G288. [Google Scholar] [PubMed]
- Small, K.; Feng, J.F.; Lorenz, J.; Donnelly, E.T.; Yu, A.; Im, M.J.; Dorn, G.W., 2nd; Liggett, S.B. Cardiac specific overexpression of transglutaminase II G(h) results in a unique hypertrophy phenotype independent of phospholipase C activation. J. Biol. Chem. 1999, 274, 21291–21296. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.S.; Fisher, M.; Haylor, J.L.; Hau, Z.; Skill, N.J.; Jones, R.; Saint, R.; Coutts, I.; Vickers, M.E.; El Nahas, A.M.; et al. Transglutaminase inhibition reduces fibrosis and preserves function in experimental chronic kidney disease. J. Am. Soc. Nephrol. 2007, 18, 3078–3088. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.S.; Greenlee, B.M.; Wills-Karp, M.; Moller, D.R. Attenuation of lung inflammation and fibrosis in interferon-γ-deficient mice after intratracheal bleomycin. Am. J. Respir. Cell Mol. Biol. 2001, 24, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Marko, L.; Kvakan, H.; Park, J.K.; Qadri, F.; Spallek, B.; Binger, K.J.; Bowman, E.P.; Kleinewietfeld, M.; Fokuhl, V.; Dechend, R.; et al. Interferon-γ signaling inhibition ameliorates angiotensin II-induced cardiac damage. Hypertension 2012, 60, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.L.; Li, Y.L.; Jia, L.X.; Cheng, J.Z.; Qi, Y.F.; Zhang, H.J.; Du, J. Reciprocal interaction between macrophages and T cells stimulates IFN-gamma and MCP-1 production in Ang II-induced cardiac inflammation and fibrosis. PLoS ONE 2012, 7, e35506. [Google Scholar]
- Eickelberg, O.; Pansky, A.; Koehler, E.; Bihl, M.; Tamm, M.; Hildebrand, P.; Perruchoud, A.P.; Kashgarian, M.; Roth, M. Molecular mechanisms of TGF-β antagonism by interferon γ and cyclosporine a in lung fibroblasts. FASEB J. 2001, 15, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, D.; Frisancho-Kiss, S.; Yusung, S.A.; Barrett, M.A.; Davis, S.E.; Gatewood, S.J.; Njoku, D.B.; Rose, N.R. Interferon-γ protects against chronic viral myocarditis by reducing mast cell degranulation, fibrosis, and the profibrotic cytokines transforming growth factor-β1, interleukin-1β, and interleukin-4 in the heart. Am. J. Pathol. 2004, 165, 1883–1894. [Google Scholar] [CrossRef]
- Shao, D.D.; Suresh, R.; Vakil, V.; Gomer, R.H.; Pilling, D. Pivotal advance: Th-1 cytokines inhibit, and Th-2 cytokines promote fibrocyte differentiation. J. Leukoc. Biol. 2008, 83, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Eckert, R.L.; Sturniolo, M.T.; Broome, A.M.; Ruse, M.; Rorke, E.A. Transglutaminase function in epidermis. J. Investig. Dermatol. 2005, 124, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, K. Transglutaminase in skin epidermis. Eur. J. Dermatol. 2005, 15, 313–319. [Google Scholar] [PubMed]
- Zeeuwen, P.L. Epidermal differentiation: The role of proteases and their inhibitors. Eur. J. Cell Biol. 2004, 83, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Inada, R.; Matsuki, M.; Yamada, K.; Morishima, Y.; Shen, S.C.; Kuramoto, N.; Yasuno, H.; Takahashi, K.; Miyachi, Y.; Yamanishi, K. Facilitated wound healing by activation of the Transglutaminase 1 gene. Am. J. Pathol. 2000, 157, 1875–1882. [Google Scholar] [CrossRef]
- Bognar, P.; Nemeth, I.; Mayer, B.; Haluszka, D.; Wikonkal, N.; Ostorhazi, E.; John, S.; Paulsson, M.; Smyth, N.; Pasztoi, M.; et al. Reduced inflammatory threshold indicates skin barrier defect in transglutaminase 3 knockout mice. J. Investig. Dermatol. 2014, 134, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currò, M.; Matarese, G.; Isola, G.; Caccamo, D.; Ventura, V.P.; Cornelius, C.; Lentini, M.; Cordasco, G.; Ientile, R. Differential expression of transglutaminase genes in patients with chronicperiodontitis. Oral Dis. 2014, 20, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.A.; Berika, M.Y.; Alghadir, A.H. Apoptosis and clinical severity in patients with psoriasis and HCV infection. Indian J. Dermatol. 2014, 59, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Rodolfo, C.; Mormone, E.; Matarrese, P.; Ciccosanti, F.; Farrace, M.G.; Garofano, E.; Piredda, L.; Fimia, G.M.; Malorni, W.; Piacentini, M. Tissue transglutaminase is a multifunctional BH3-only protein. J. Biol. Chem. 2004, 279, 54783–54792. [Google Scholar] [CrossRef] [PubMed]
- Fesus, L.; Szondy, Z. Transglutaminase 2 in the balance of cell death and survival. FEBS Lett. 2005, 579, 3297–3302. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, L.; Farrace, M.G.; Nini, G.; Piacentini, M. Abnormal Bcl-2 and “Tissue” Transglutaminase expression in psoriatic skin. J. Investig. Dermatol. 1994, 103, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Presland, R.B.; Jurevic, R.J. Making sense of the epithelial barrier: What molecular biology and genetics tell us about the functions of oral mucosal and epidermal tissues. J. Dent. Educ. 2002, 66, 564–574. [Google Scholar] [PubMed]
- Gooding, J.M.; Yap, K.L.; Ikura, M. The cadherin-catenin complex as a focal point of cell adhesion and signalling: New insights from three-dimensional structures. Bioessays 2004, 26, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Neppelberg, E.; Loro, L.L.; Oijordsbakken, G.; Johannessen, A.C. Altered CD40 and E-cadherin expression—putative role in oral lichen planus. J. Oral Pathol. Med. 2007, 36, 153–160. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, T.; Currò, M.; Barbera, A.; Caccamo, D.; Antonuccio, P.; Arena, S.; Montalto, A.S.; Parisi, S.; Marseglia, L.; Gitto, E.; et al. Expression of Transglutaminase in Foreskin of Children with Balanitis Xerotica Obliterans. Int. J. Mol. Sci. 2016, 17, 1551. https://doi.org/10.3390/ijms17091551
Russo T, Currò M, Barbera A, Caccamo D, Antonuccio P, Arena S, Montalto AS, Parisi S, Marseglia L, Gitto E, et al. Expression of Transglutaminase in Foreskin of Children with Balanitis Xerotica Obliterans. International Journal of Molecular Sciences. 2016; 17(9):1551. https://doi.org/10.3390/ijms17091551
Chicago/Turabian StyleRusso, Tiziana, Monica Currò, Anna Barbera, Daniela Caccamo, Pietro Antonuccio, Salvatore Arena, Angela Simona Montalto, Saveria Parisi, Lucia Marseglia, Eloisa Gitto, and et al. 2016. "Expression of Transglutaminase in Foreskin of Children with Balanitis Xerotica Obliterans" International Journal of Molecular Sciences 17, no. 9: 1551. https://doi.org/10.3390/ijms17091551
APA StyleRusso, T., Currò, M., Barbera, A., Caccamo, D., Antonuccio, P., Arena, S., Montalto, A. S., Parisi, S., Marseglia, L., Gitto, E., Ientile, R., Impellizzeri, P., & Romeo, C. (2016). Expression of Transglutaminase in Foreskin of Children with Balanitis Xerotica Obliterans. International Journal of Molecular Sciences, 17(9), 1551. https://doi.org/10.3390/ijms17091551