The NAC Transcription Factor Gene OsY37 (ONAC011) Promotes Leaf Senescence and Accelerates Heading Time in Rice

Abstract
:1. Introduction
2. Results
2.1. Isolation of the OsY37I Gene
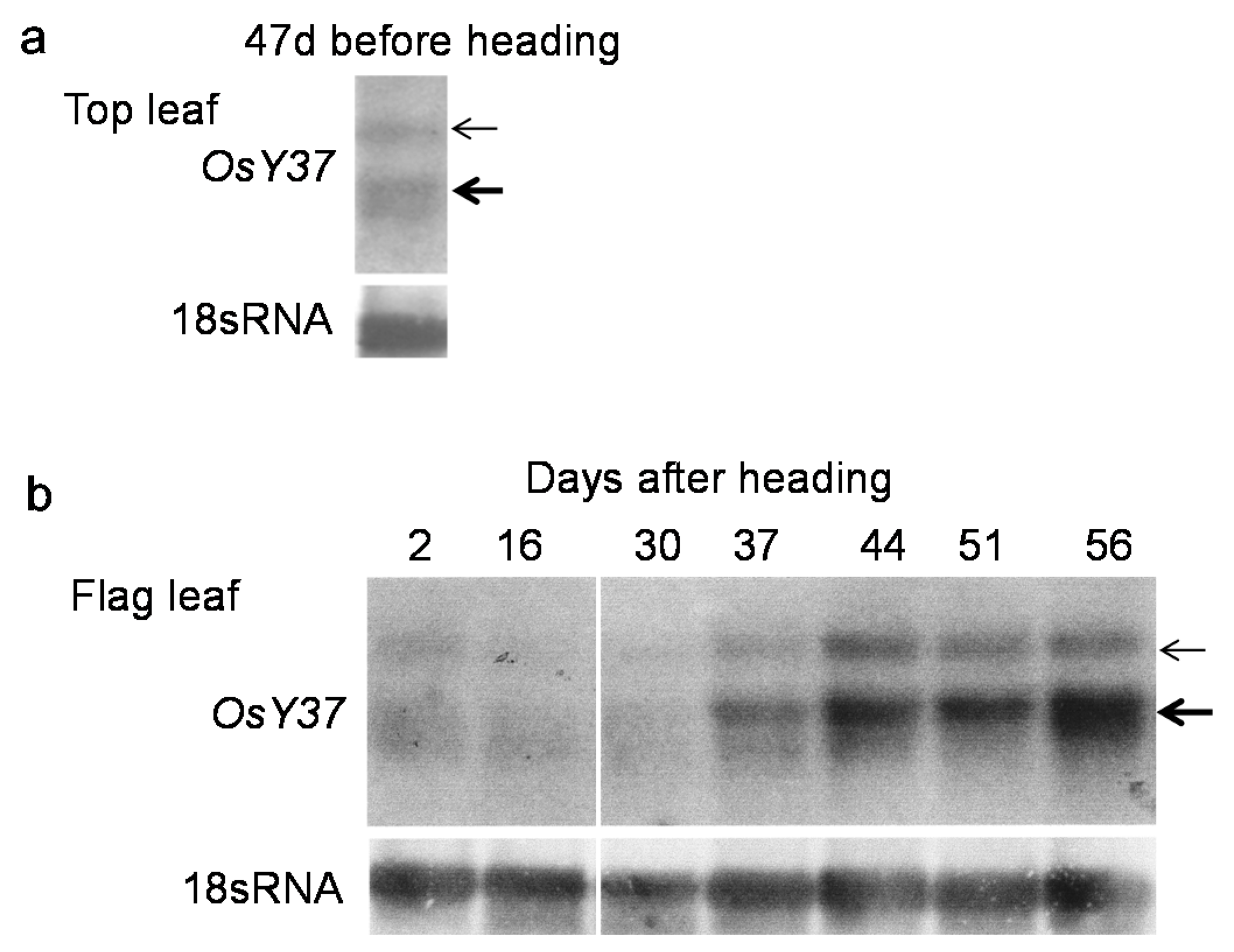
2.2. Increased Accumulation of OsY37 Transcripts during Natural Senescence
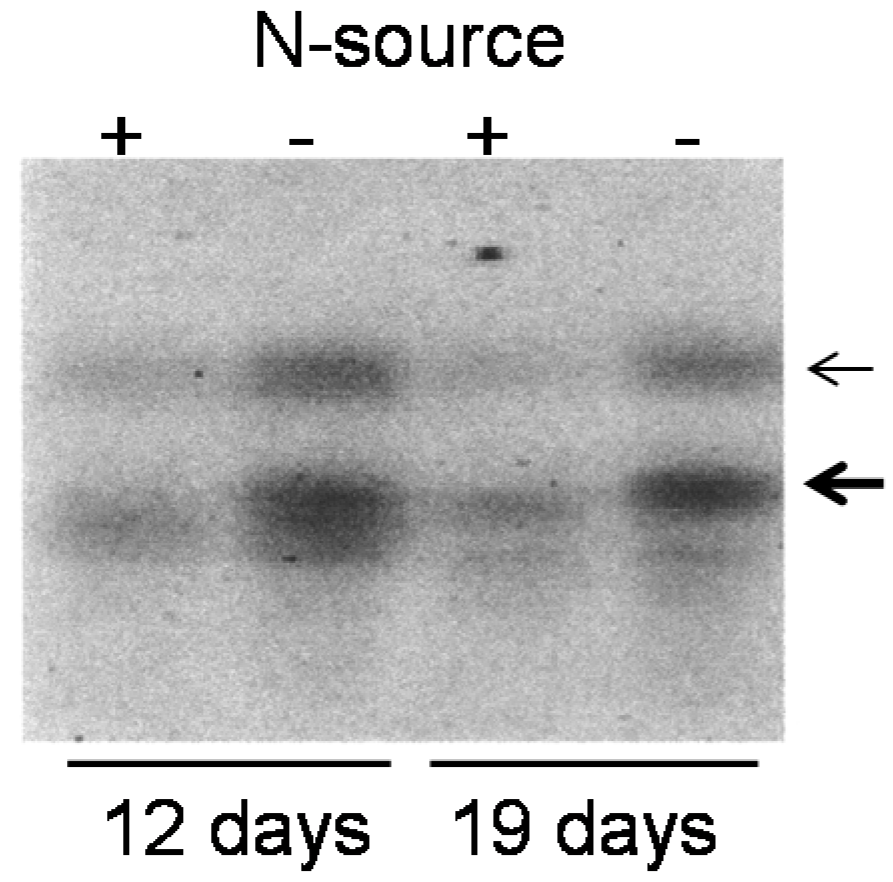
2.3. Induction of OsY37 Expression Under Nitrogen Starvation Conditions
2.4. Nuclear Localization of GFP/OsY37 Fusion Polypeptide
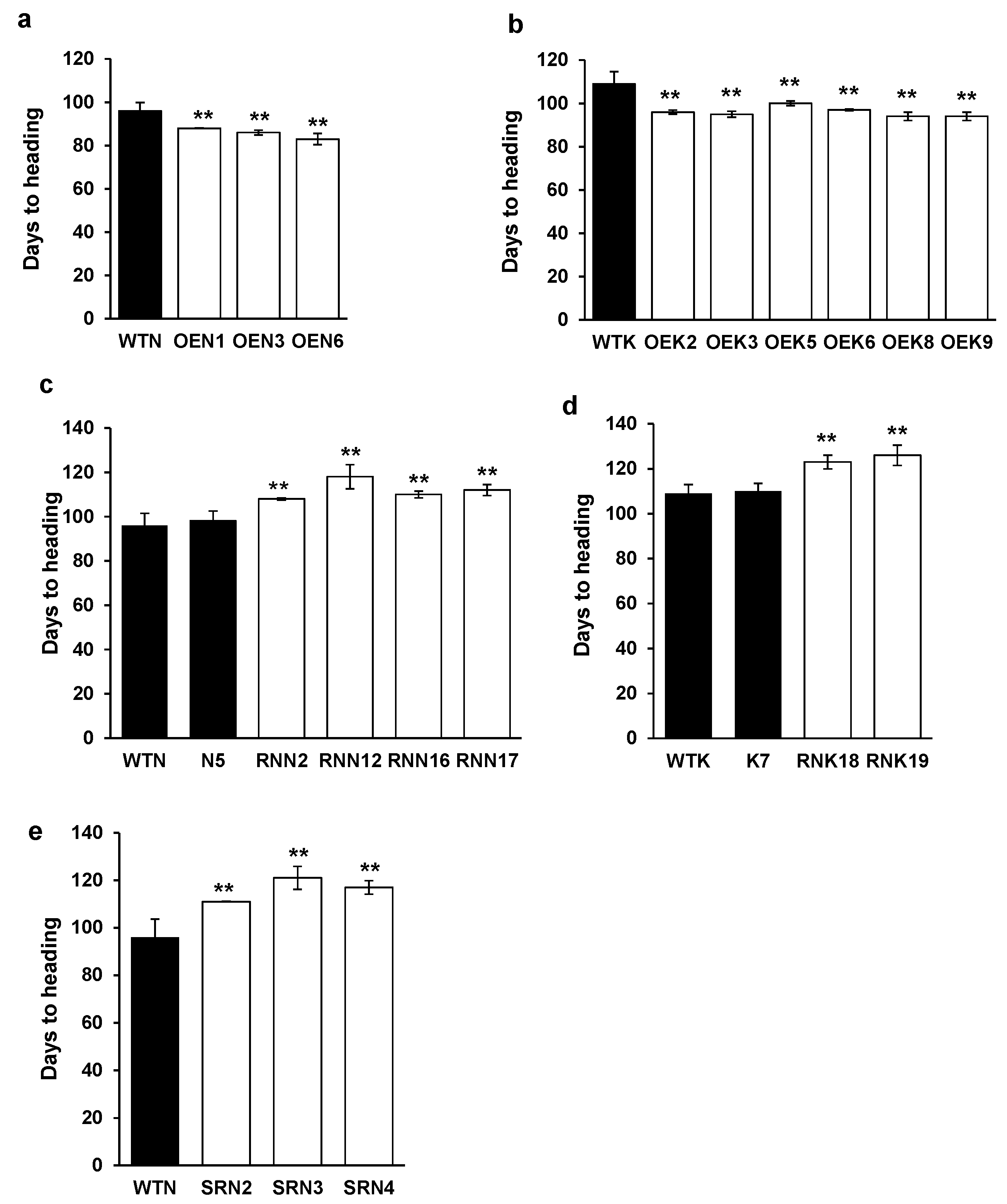
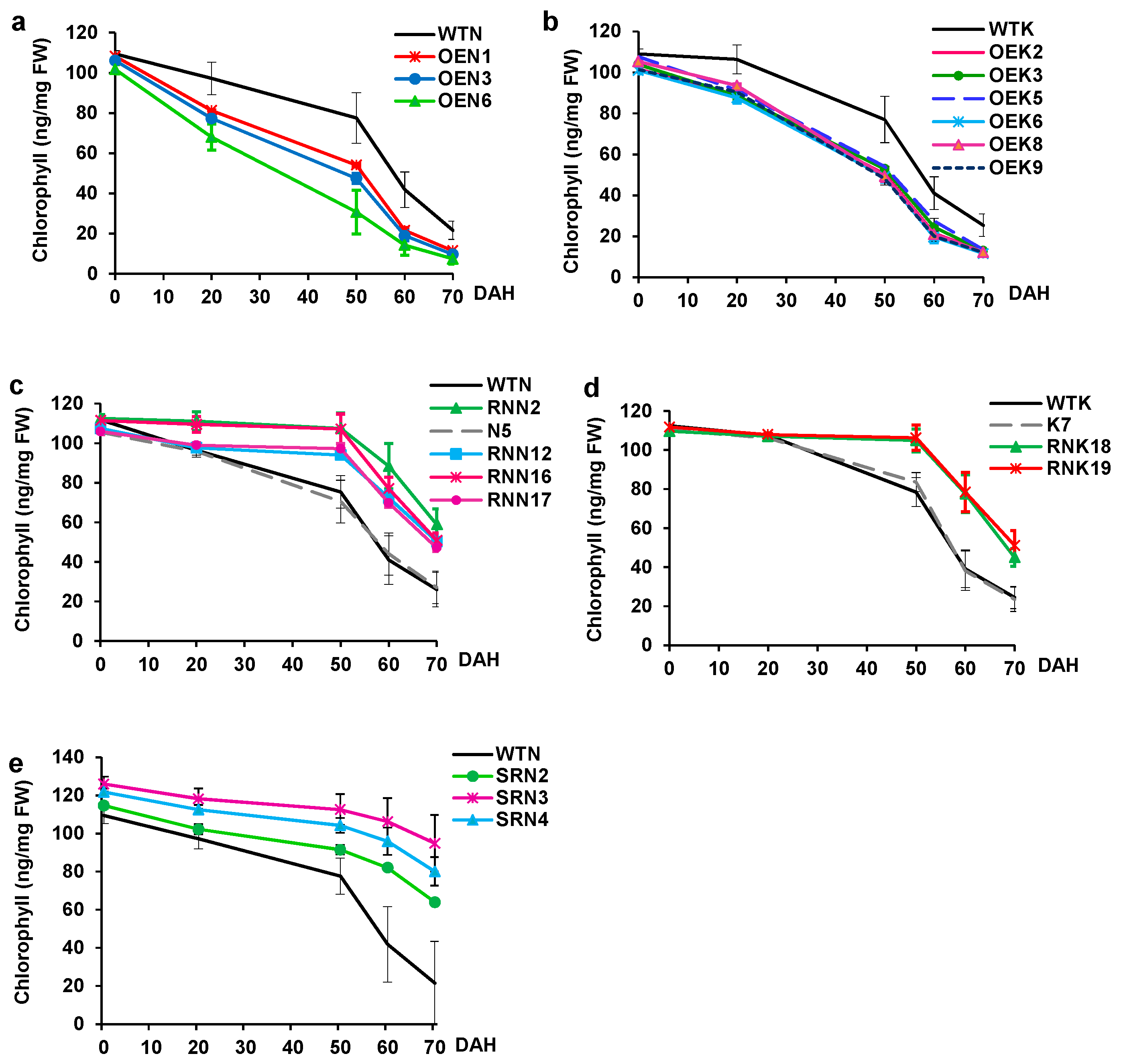
2.5. Effects of Overexpression and Knockdown of OsY37 on Heading Date and Leaf Senescence
2.6. Effects of Modulated OsY37 Versions on Vegetative Growth and Plant Development
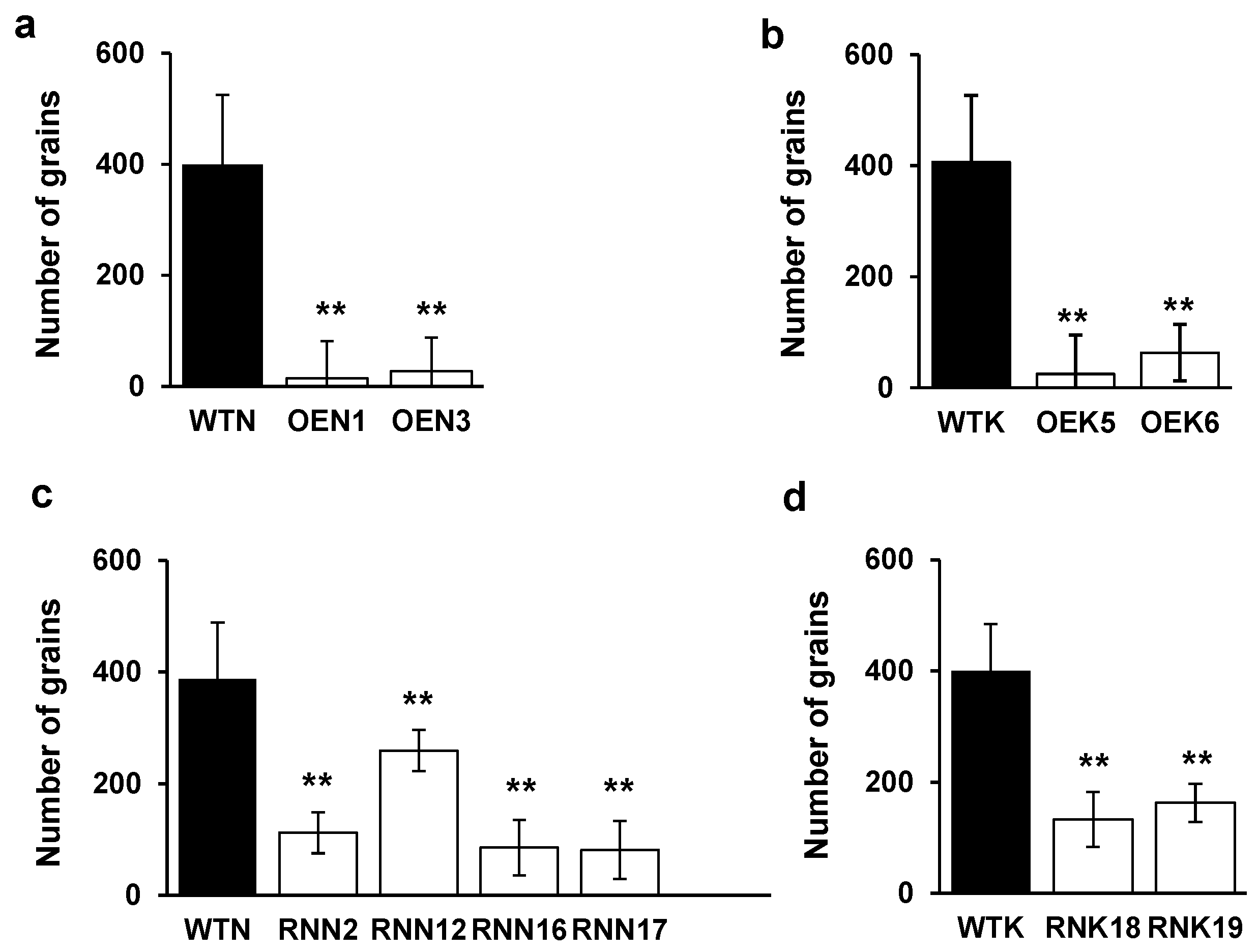
2.7. Effects of Modified OsY37 Genes on Grain Yield in Rice
3. Discussion
3.1. OsY37 Gene Expression Profile
3.2. Dual Functions of the OsY37 Gene in the Reproductive Phase
3.3. Involvement of OsY37 in Vegetative Growth and Development
3.4. Functional Comparison with the Other NAC Genes
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Plasmid Vectors and Primers
4.3. Measurement of Chlorophyll Content
4.4. RNA Extraction and Northern Hybridization Analysis
4.5. Differential Display
4.6. Full-Length cDNA Library Screening and Sub-Cloning
4.7. Generation of Recombinant Clones
4.8. Nitrogen Starvation
4.9. RT-PCR
4.10. Transformation of Rice and Arabidopsis
4.11. Particle Bombardment
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| pAct | Rice actin promoter |
| pCaMV35S | Cauliflower mosaic virus 35S promoter |
| CRES-T | Chimeric repressor silencing technology |
| GFP | Green fluorescent protein |
| HgrR | Hygromycin-resistant |
| MS | Murashige and Skoog |
| NAC | NAM, ATAF, and CUC |
| RNAi | RNA interference |
References
- Matile, P.; Ginsburg, S.; Schellenberg, M.; Thomass, H. Catabolites of chlorophyll in senescing barley leaves are localized in the vacuoles of mesophyll cells. Proc. Natl. Acad. Sci. USA 1988, 85, 9529–9532. [Google Scholar] [CrossRef] [PubMed]
- Drake, R.; John, I.; Farrell, A.; Cooper, W.; Schuch, W.; Grierson, D. Isolation and analysis of cDNAs encoding tomato cysteine proteases expressed during leaf senescence. Plant Mol. Biol. 1996, 30, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Tournaire, C.; Kushnir, S.; Bauw, G.; Inzé, D.; de la Serve, B.T.; Renaudin, J.P. A thiol protease and an anionic peroxidase are induced by lowering cytokinins during callus growth. Plant Physiol. 1996, 111, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Blank, A.; McKeon, T.A. Single-strand-preferring nuclease activity in wheat leaves is increased in senescence and is negatively photoregulated. Proc. Natl. Acad. Sci. USA 1989, 86, 3169–3173. [Google Scholar] [CrossRef] [PubMed]
- Lers, A.; Lomaniec, E.; Burd, S.; Khalchitski, A. The characterization of LeNUC1, a nucleae associated with leaf senescence of tomato. Physiol. Plant. 2001, 112, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Masclaux, C.; Valadier, M.H.; Brugiere, N.; Morot-Gaudry, J.F.; Hirel, B. Chraracterization of the sink/source transition in tobacco (Nicotiana tabacum L.) shoots in relation to nitrogen management and leaf senescence. Planta 2000, 211, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Quirino, B.F.; Reiter, W.D.; Amasino, R.D. One of two tandem Arabidopsis genes homologous to moaccharide transporters is senescence-associated. Plant Mol. Biol. 2001, 46, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Soudry, E.; Ulitzur, S.; Gepstein, S. Accumulation and remobilization of amino acids during senescence of detached and attached leaves: In planta analysis of tryptophan levels by recombinant luminescent bacteria. J. Exp. Bot. 2005, 56, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Distelfeld, A.; Avni, R.; Fischer, A.M. Senescence, nutrient remobilization, and yield in wheat and barley. J. Exp. Bot. 2014, 65, 3783–3798. [Google Scholar] [CrossRef] [PubMed]
- Abou-khalifa, A.A.B.; Misra, A.N.; Salem, A.E. Effect of leaf cutting on physiological traits and yield of two rice cultivars. Afr. J. Plant Sci. 2008, 2, 147–150. [Google Scholar]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Lu, S.; Lv, B.; Zhang, B.; Shen, J.; He, J.; Luo, L.; Xi, D.; Chen, X.; Ming, F. A rice NAC transcription factor promotes leaf senescence via ABA biosynthesis. Plant Physiol. 2017, 174, 1747–1763. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant senescence and crop productivity. Plant Mol. Biol. 2013, 82, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Zagotta, M.T.; Hicks, K.A.; Jacobs, C.I.; Young, J.C.; Hangarter, R.P.; Meeks-Wagner, D.R. The Arabidopsis ELF3 gene regulates vegetative photomorphogenesis and the photoperiodic induction of flowering. Plant J. 1996, 10, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Han, S.-H.; Yang, H.-J.; Piao, W.; Paek, N.C. Mutation of Rice Early Flowering3.1 (OsELF301) delays leaf senescence in rice. Plant Mol. Biol. 2016, 92, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.S.; Penfold, C.A.; Jenkins, D.; et al. High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.; El-Kereamy, A.; Kim, S.H.; Nambara, E.; Rothstein, S.J. ANAC032 positively regulates age-dependent and stress-induced senescence in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 2029–2046. [Google Scholar] [CrossRef] [PubMed]
- Gepstein, S.; Sabehi, G.; Carp, M.J.; Hajouj, T.; Nesher, M.F.; Yariv, I.; Dor, C.; Bassani, M. Large-scale identification of leaf senescence-associated genes. Plant J. 2003, 36, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Bhalerao, R.; Keskitalo, J.; Sterky, F.; Erlandsson, R.; Björkbacka, H.; Birve, S.J.; Karlsson, J.; Gardeström, P.; Gustafsson, P.; Lundeberg, J.; et al. Gene expression in autumn leaves. Plant Physiol. 2003, 131, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Van der Graaff, E.; Schwacke, R.; Schneider, A.; Desimone, M.; Flügge, U.I.; Kunze, R. Transcription analysis of Arabidopsis membrane transporters and hormone pathways during developmental and induced leaf senescence. Plant Physiol. 2006, 141, 776–792. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhou, Y.; Zhou, G.; Ye, R.; Zhao, L.; Li, X.; Lin, Y. Identification of early senescence-associated genes in rice flag leaves. Plant Mol. Biol. 2008, 67, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, J.; Wen, X.; Guo, H. Gene network analysis and functional studies of senescence-associated genes reveal novel regulators of Arabidopsis leaf senescence. J. Integr. Plant Biol. 2012, 54, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryva sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; You, J.; Xie, K.; Xie, W.; Xiong, L. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Sakuraba, Y.; Han, S.-H.; Yoo, S.C.; Paek, N.C. Mutation of the Arabidopsis NAC016 transcription factor delays leaf senescence. Plant Cell Physiol. 2013, 54, 1660–1672. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, W.; Liu, L.; Chen, T.; Zhou, F.; Lin, Y. Identification and functional characterization of a rice NAC gene involved in the regulation of leaf senescence. BMC Plant Biol. 2013, 13, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Balazadeh, S.; Kwasniewski, M.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Xue, G.P.; Mueller-Roeber, B. ORS1, an H2O2-responsive NAC transcription factor, controls senescence in Arabidopsis thaliana. Mol. Plant 2011, 4, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.I.; Asensi-Fabado, A.; Munné-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 2012, 24, 482–506. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.D.; Seo, P.J.; Yoon, H.K.; Park, C.M. The Arabidopsis NAC transcription factor VNI2 integrates abscisic acid signals into leaf senescence via the COR/RD genes. Plant Cell 2011, 23, 2155–2168. [Google Scholar] [CrossRef] [PubMed]
- Agüera, E.; Cabello, P.; de la Haba, P. Induction of leaf senescence by low nitrogen nutrition in sunflower (Helianthus annuus) plants. Physiol. Plant. 2010, 138, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Hiratsu, K.; Matsui, K.; Koyama, T.; Ohme-Takagi, M. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. 2003, 34, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.; Saliba-Colombani, V.; Loudet, O.; Bellunomo, P.; Moreau, L.; Daniel-Vedele, F.; Morot-Gaudry, J.F.; Masclaux-Daubresse, C. Leaf yellowing and anthocyanin accumulation are two genetically independent strategies in response to nitrogen limitation in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Oda-Yamamizo, C.; Mitsuda, N.; Sakamoto, S.; Ogawa, D.; Ohme-Takagi, M.; Ohmiya, A. The NAC transcription factor ANAC046 is a positive regulator of chlorophyll degradation and senescence in Arabidopsis leaves. Sci. Rep. 2016, 6, 2–12. [Google Scholar]
- Procházková, D.; Wilhelmová, N. Leaf senescence and activities of the antioxidant enzymes. Biol. Plant. 2007, 51, 401–406. [Google Scholar] [CrossRef]
- Lai, A.G.; Doherty, C.J.; Mueller-Roeber, B.; Kay, S.A.; Schippers, J.H.M.; Dijkwel, P.P. Circadian clock-associated1 regulates ROS homeostasis and oxidative stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 17129–17134. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; de Tullio, M.; Conklin, P.L. The role of ascorbic acid in the control of flowering time and the onset of senescence. J. Exp. Bot. 2006, 57, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Vergara, B.S.; Lilis, R.; Tanaka, A. Relationship between length of growing period and yield of rice plants under a limited nitrogen supply. Soil Sci. Plant Nutr. 1964, 10, 15–21. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J.; et al. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Gan, S. AtNAP, a NAC transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Woo, H.R.; Kim, J.; Lim, P.O.; Lee, I.C.; Choi, S.H.; Hwang, D.; Nam, H.G. Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science 2009, 323, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Piao, W.; Lim, J.-H.; Han, S.-H.; Kim, Y.-S.; An, A.; Paek, N.-C. Rice ONAC106 inhibits leaf senescence and increases salt tolerance and tiller angle. Plant Cell Physiol. 2015, 56, 2325–2339. [Google Scholar] [CrossRef] [PubMed]
- Khanna-Chopra, R. Leaf senescence and abiotic stresses share reactive oxygen species-mediated chloroplast degradation. Protoplasma 2012, 249, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Rogers, H.; Munné-Bosch, S. Production and scavenging of reactive oxygen species and redox signaling during leaf and flower senescence: Similar but different. Plant Physiol. 2016, 171, 1560–1568. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Niwa, Y.; Hirano, T.; Yoshimoto, K.; Shimizu, M.; Kobayashi, H. Non-invasive quantitative detection and applications of non-toxic, S65T-type green fluorescent protein in living plants. Plant J. 1999, 18, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Bruinsma, J. A comment on the spectrophotometric determination of chlorophyll. Biochim. Biophys. Acta 1961, 52, 576–578. [Google Scholar] [CrossRef]
- Gális, I.; Šimek, P.; van Onckelen, H.A.; Kakiuchi, Y.; Wabiko, H. Resistance of transgenic tobacco seedlings expressing the Agrobacterium tumefaciens C58-6b gene, to growth-inhibitory levels of cytokinin is associated with elevated IAA levels and activation of phenylpropanoid metabolism. Plant Cell Physiol. 2002, 43, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA.
- Becker, D. Binary vectors which allow the exchange of plant selectable markers and reporter genes. Nucleic Acids Res. 1990, 18, 203. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequce |
|---|---|
| FwOsGFP | AGGAGGTGTACAAGATGAGCGGGATGAATTCGCTT |
| M13Rv | CAGGAAACAGCTATGAC |
| IndFw | TCGAGGAGGAGGGGGTGGGC |
| IndRv | GCAAATAATGTGAACTAGAATGGTAC |
| OsYIFw | ATGAGCGGGATGAATTCGCT |
| OsYIRv | ACTGAGTGAGTTCCACATTT |
| Fw24 | TCGAGGAGGAGGGGGTGGGCATGA |
| Rv869 | AGTTCCACATTTGTGTGAAGGCATGCTCAGCCAAGTAATCAAATCCATC |
| FwOsSR | ATGAGCGGGATGAATTCGCT |
| RvSRDX | ACTGAGTGAGTTCCACATTT |
| osy-2 | GCATGAGCGGGATGAATTCG |
| osy752 | GATCGCCCTACCCATATTGT |
| PromRv | GAATTCATCCGCTCATCCCGGGCCCCTCCTCCTCGAATTCCT |
| UPSPE | GGATTGTAAGATAAGTCCTC |
| OsyCRESTFw | GAATTCGAGGAGGAGGGGCCCGGGATGAGCGGGATGAATTCGCT |
| OsyCRESTRv | TATATATCTCCTTGGAAATTCCCGGGACTGAGTGAGTTCCACAT |
| hph1 | ATGTCCTGCGGGTAAATAGC |
| hph2 | CGTCTGCTGCTCCATACAAG |
| OsActinF318 | GGTATCGTCAGCAACTGGGATGATATGG |
| OsActinR675 | GCTCCGTCAGGATCTTCATGAGGTAATC |
| OsyPri1 | TGCCCGGTGATGGTCGACGT |
| SRDX-R | TTAAGCGAAACCCAAACGGAGTTCTAG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Mannai, Y.; Akabane, K.; Hiratsu, K.; Satoh-Nagasawa, N.; Wabiko, H. The NAC Transcription Factor Gene OsY37 (ONAC011) Promotes Leaf Senescence and Accelerates Heading Time in Rice. Int. J. Mol. Sci. 2017, 18, 2165. https://doi.org/10.3390/ijms18102165
El Mannai Y, Akabane K, Hiratsu K, Satoh-Nagasawa N, Wabiko H. The NAC Transcription Factor Gene OsY37 (ONAC011) Promotes Leaf Senescence and Accelerates Heading Time in Rice. International Journal of Molecular Sciences. 2017; 18(10):2165. https://doi.org/10.3390/ijms18102165
Chicago/Turabian StyleEl Mannai, Yousra, Kenta Akabane, Keiichiro Hiratsu, Namiko Satoh-Nagasawa, and Hiroetsu Wabiko. 2017. "The NAC Transcription Factor Gene OsY37 (ONAC011) Promotes Leaf Senescence and Accelerates Heading Time in Rice" International Journal of Molecular Sciences 18, no. 10: 2165. https://doi.org/10.3390/ijms18102165
APA StyleEl Mannai, Y., Akabane, K., Hiratsu, K., Satoh-Nagasawa, N., & Wabiko, H. (2017). The NAC Transcription Factor Gene OsY37 (ONAC011) Promotes Leaf Senescence and Accelerates Heading Time in Rice. International Journal of Molecular Sciences, 18(10), 2165. https://doi.org/10.3390/ijms18102165