Refined Deep-Sea Water Suppresses Inflammatory Responses via the MAPK/AP-1 and NF-κB Signaling Pathway in LPS-Treated RAW 264.7 Macrophage Cells

Abstract
:1. Introduction
2. Results
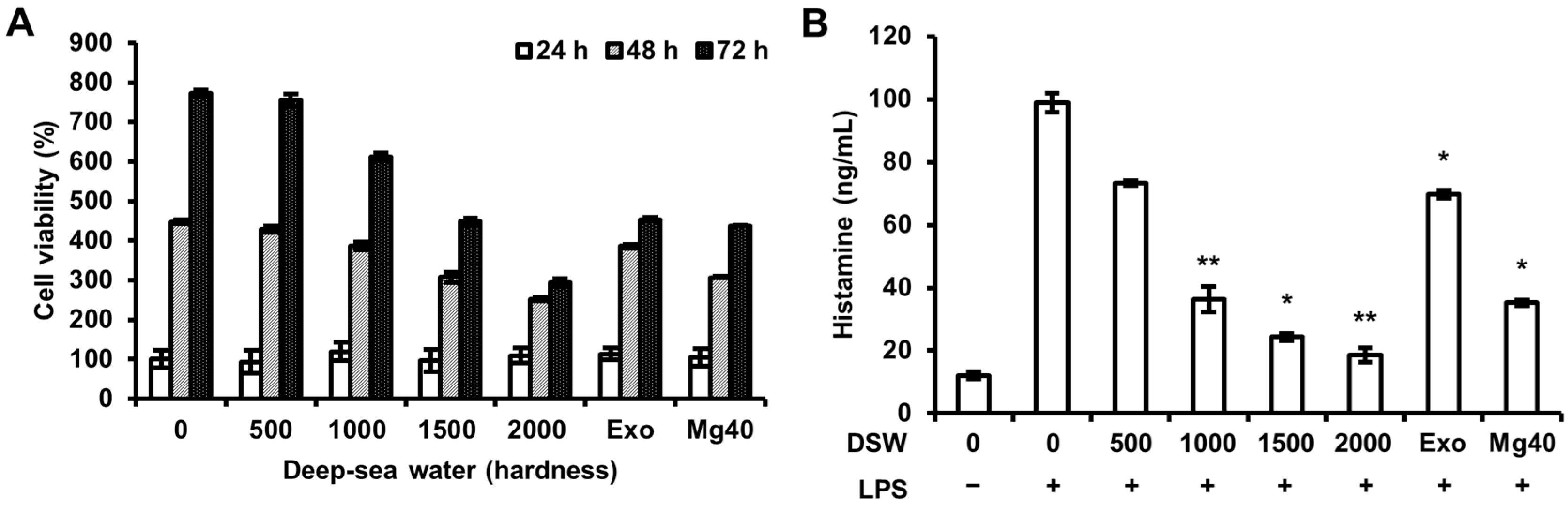
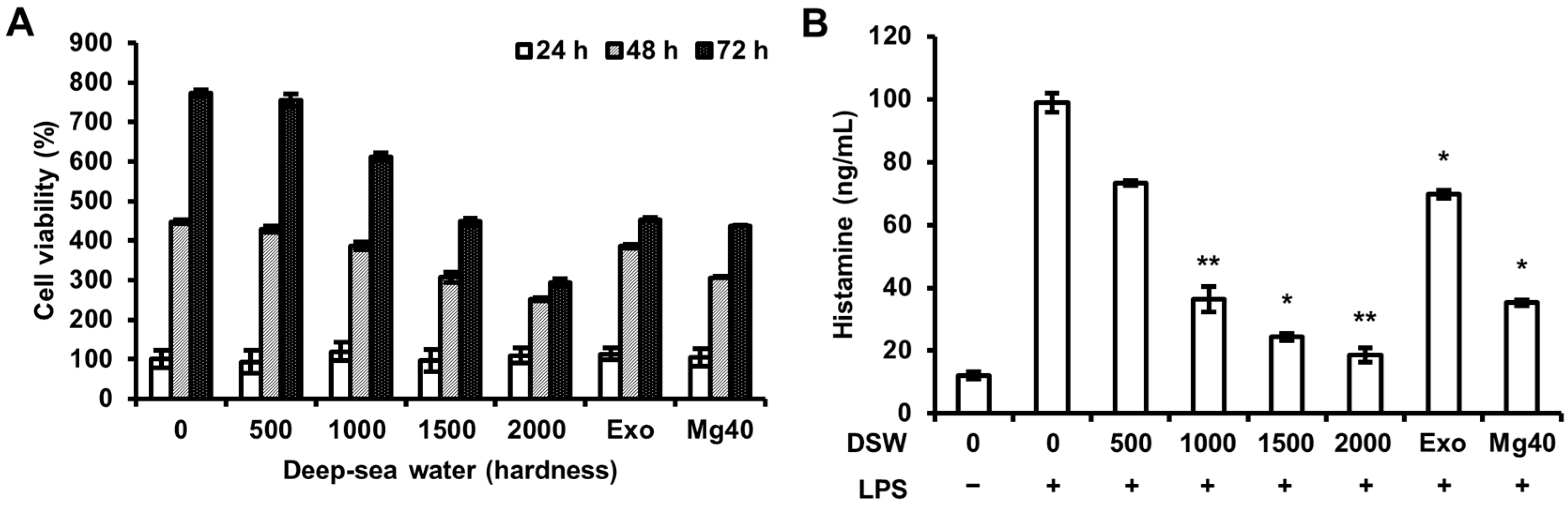
2.1. Histamine Release was Inhibited by Refined Deep-Sea Water (RDSW) on LPS-Induced RAW 264.7 Macrophage Cells
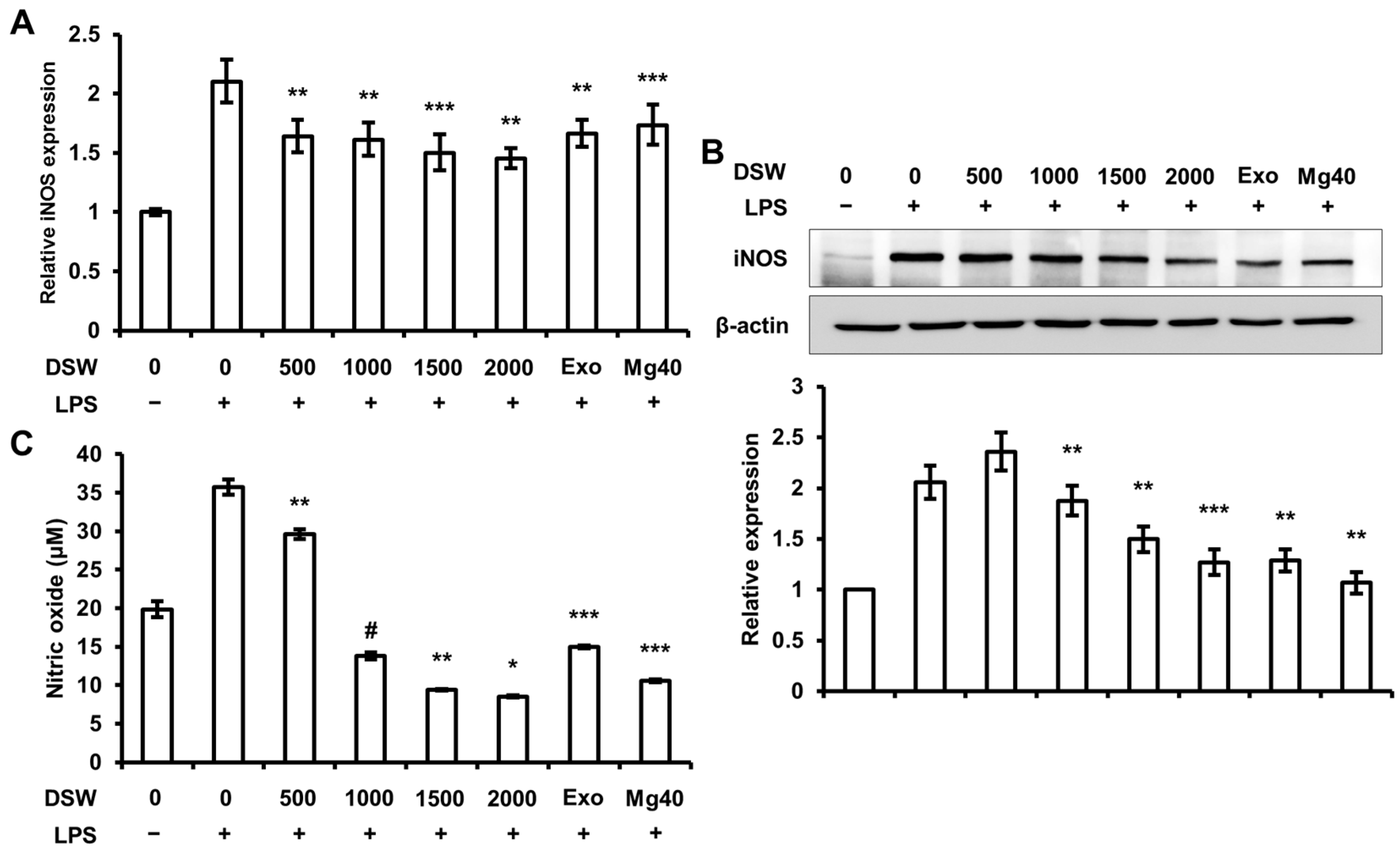
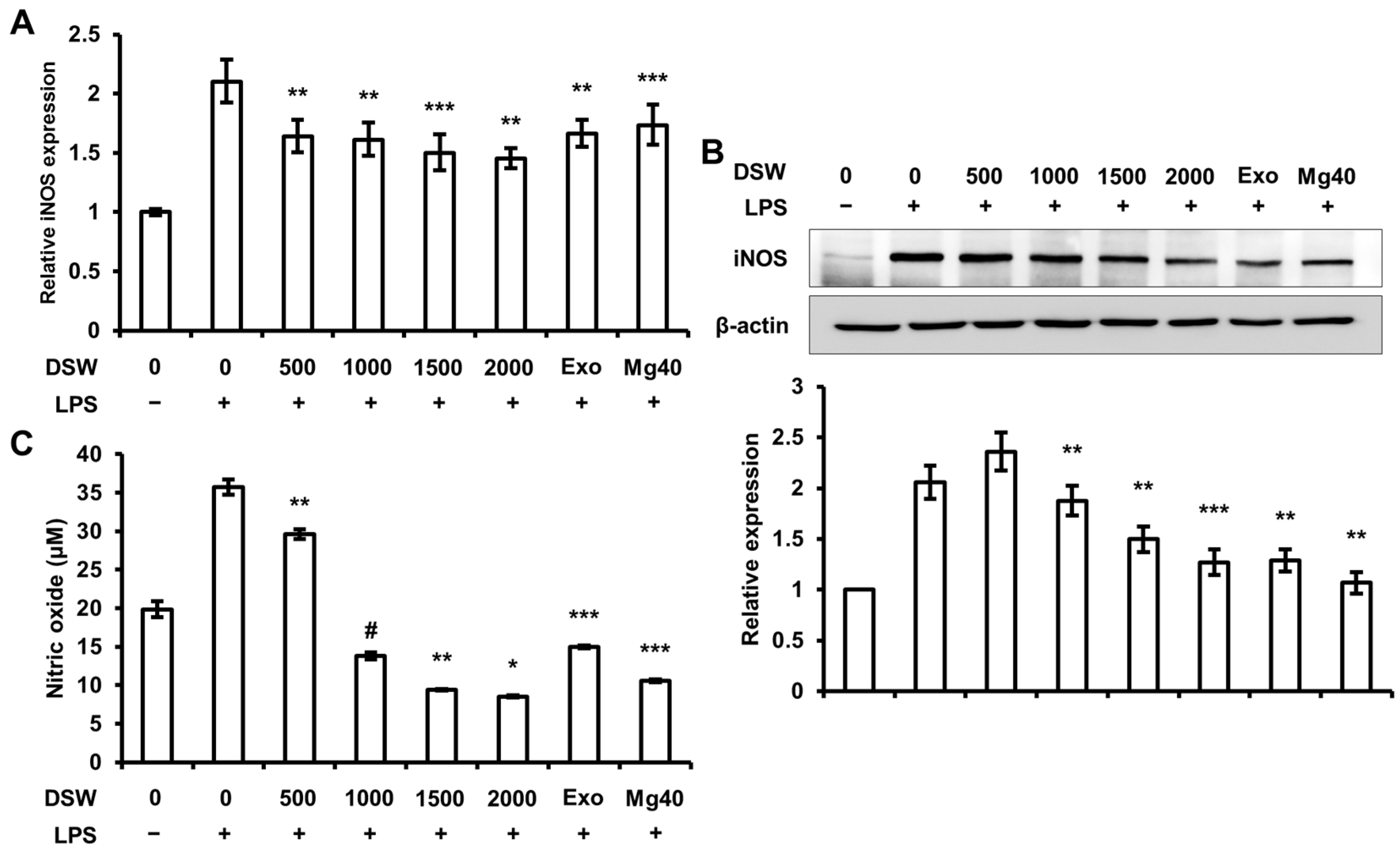
2.2. RDSW Inhibited iNOS Expressions and NO Production in LPS-Stimulated RAW 264.7 Macrophage Cells
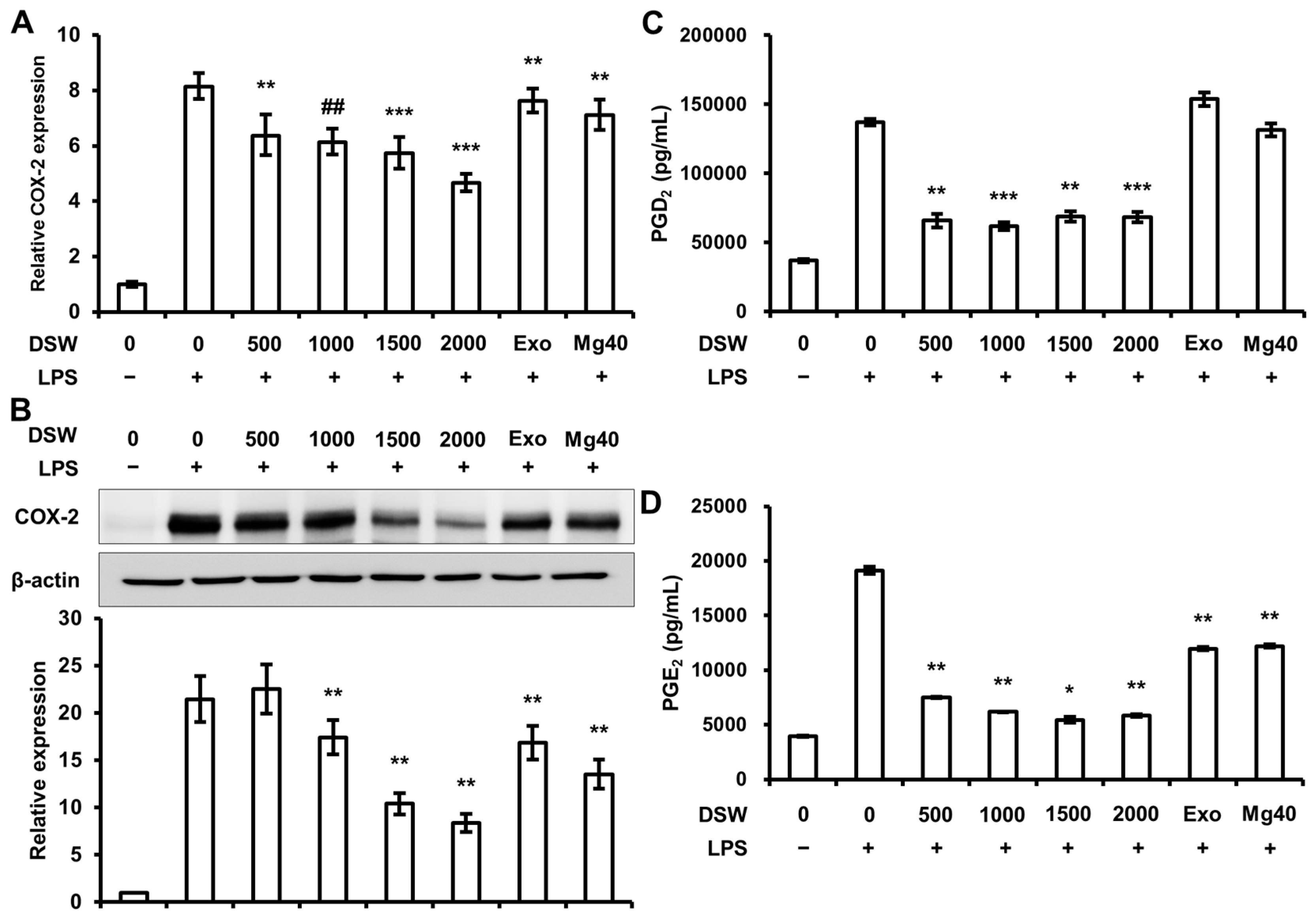
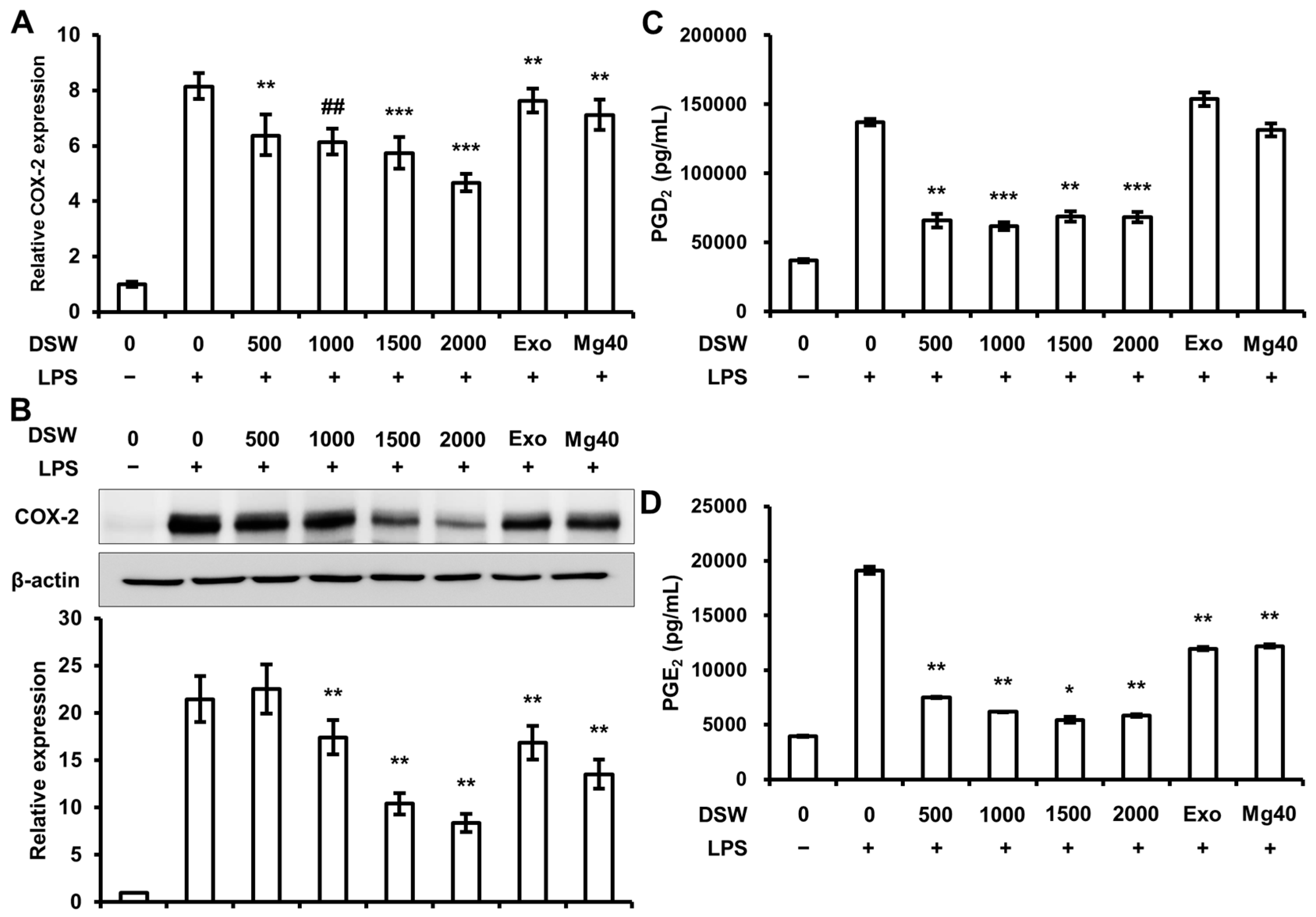
2.3. RDSW Also Suppressed COX-2 Expressions, PGD2 and PGE2 Secretions in LPS-Induced RAW 264.7 Macrophage Cells
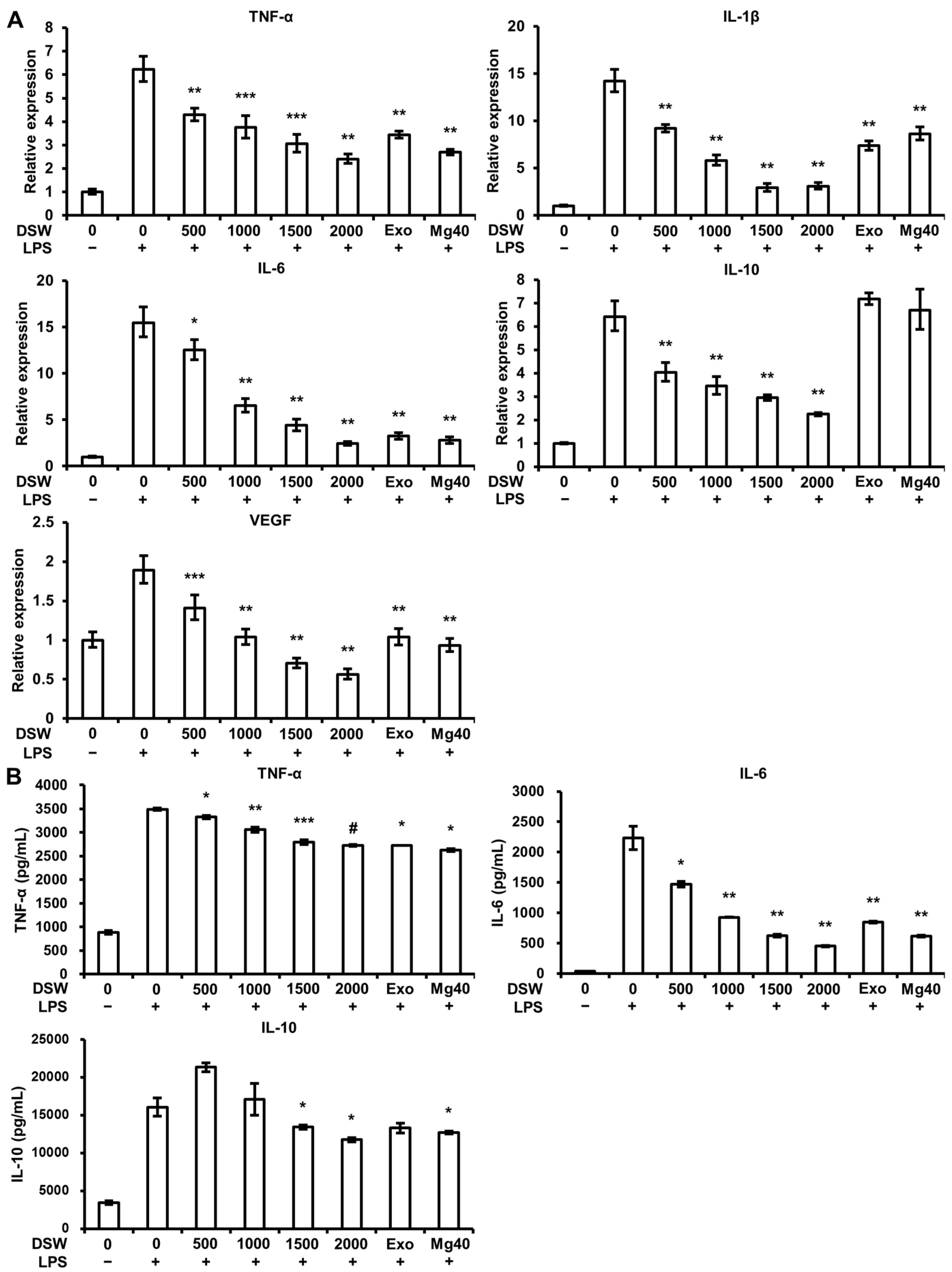
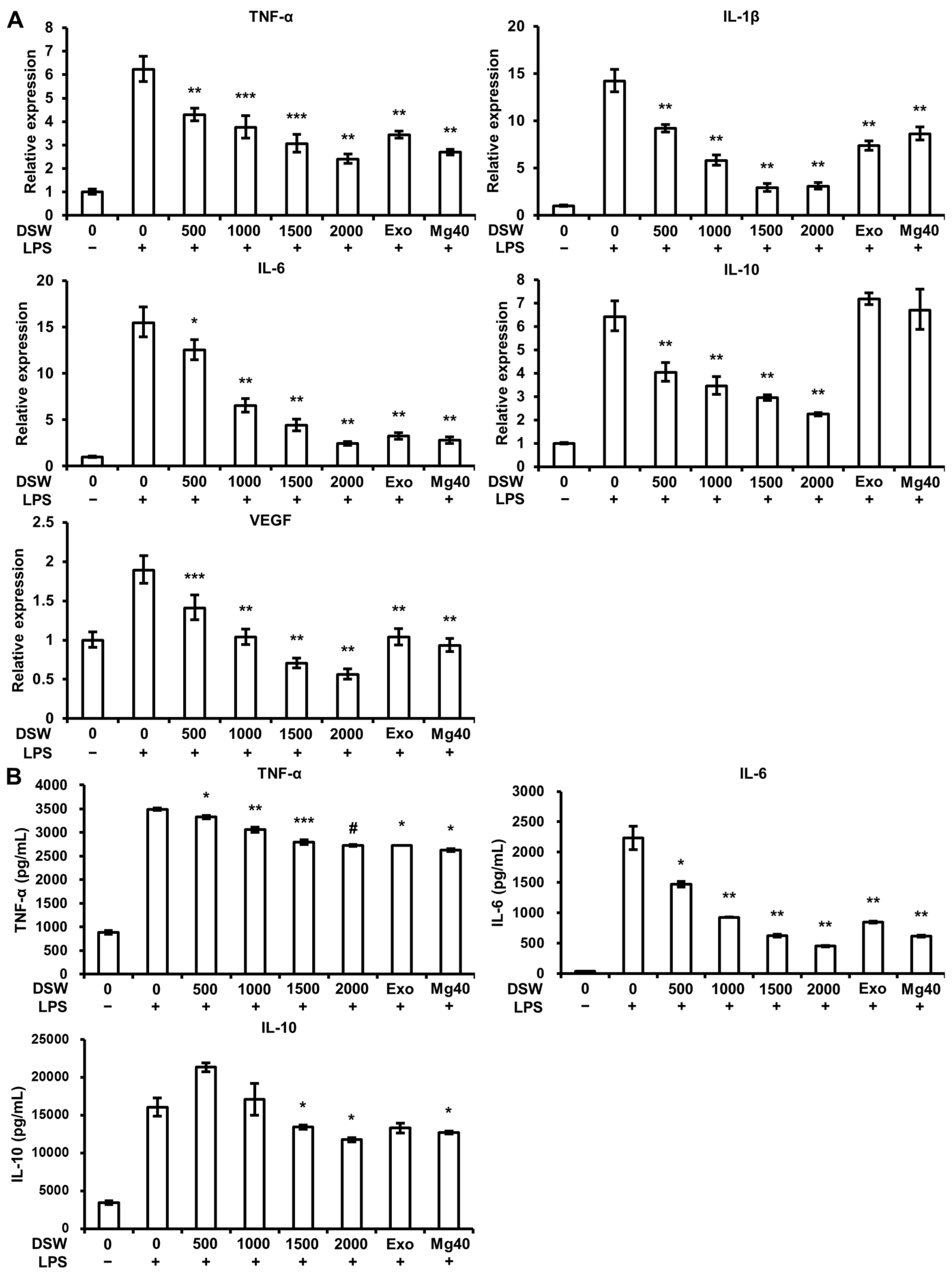
2.4. Production and Expression of Pro-Inflammatory Cytokines and VEGF Expression were Abolished by RDSW in LPS-Induced RAW 264.7 Macrophage Cells
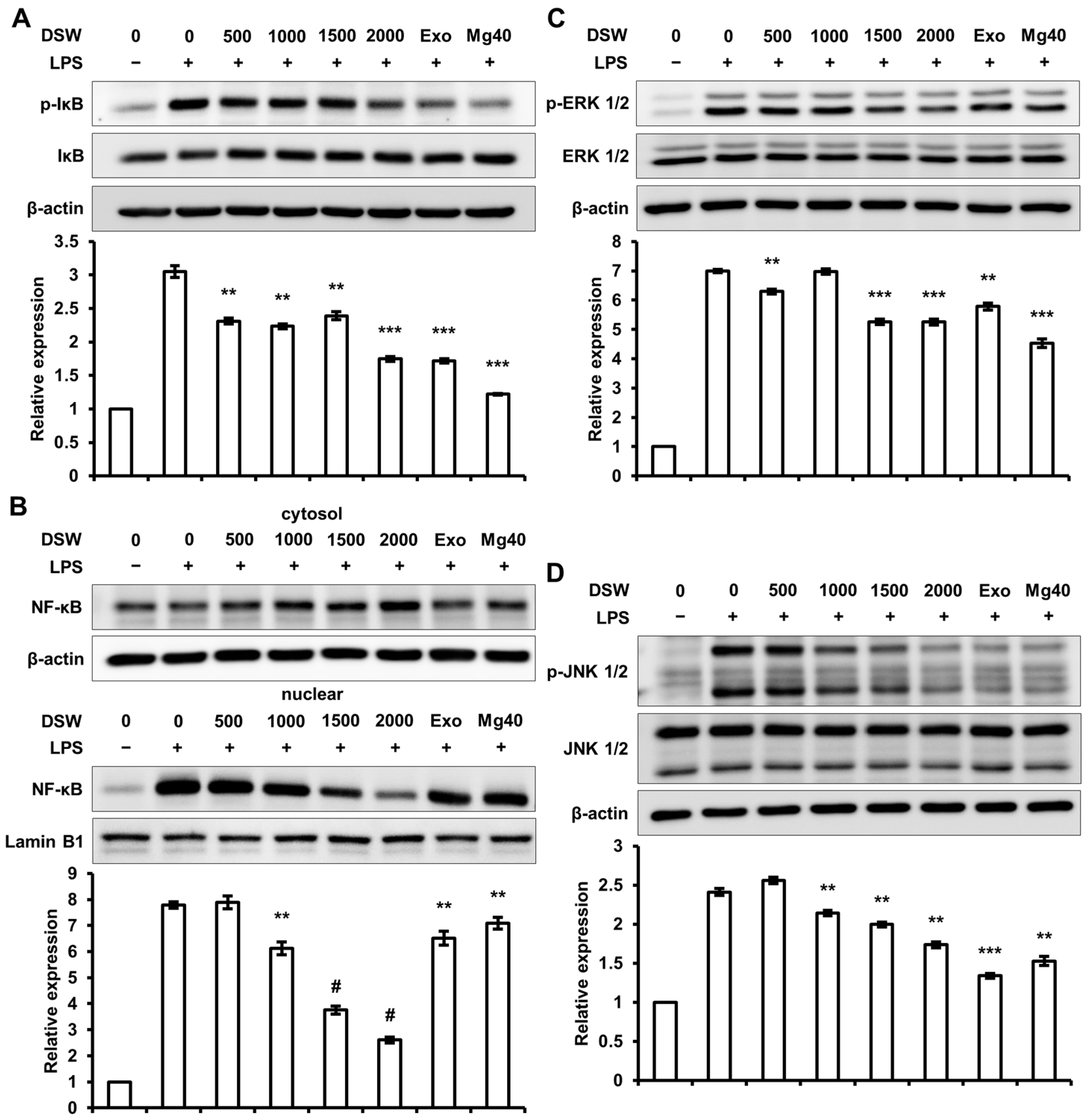
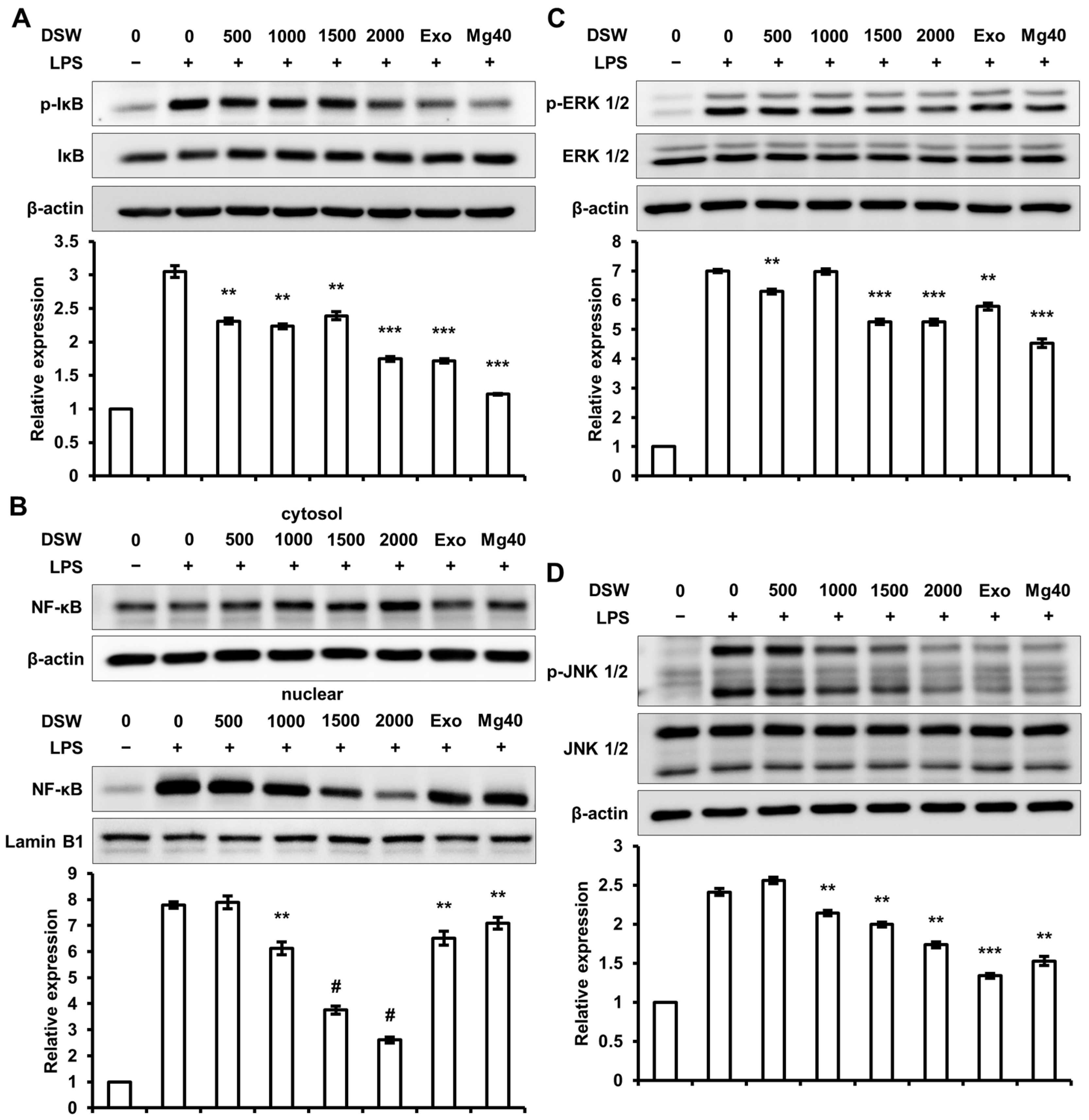
2.5. RDSW Diminished LPS-Stimulated IκB Phosphorylation, NF-κB Nuclear Translocation, and Activations of ERK 1/2 and JNK 1/2 in RAW 264.7 Macrophage Cells
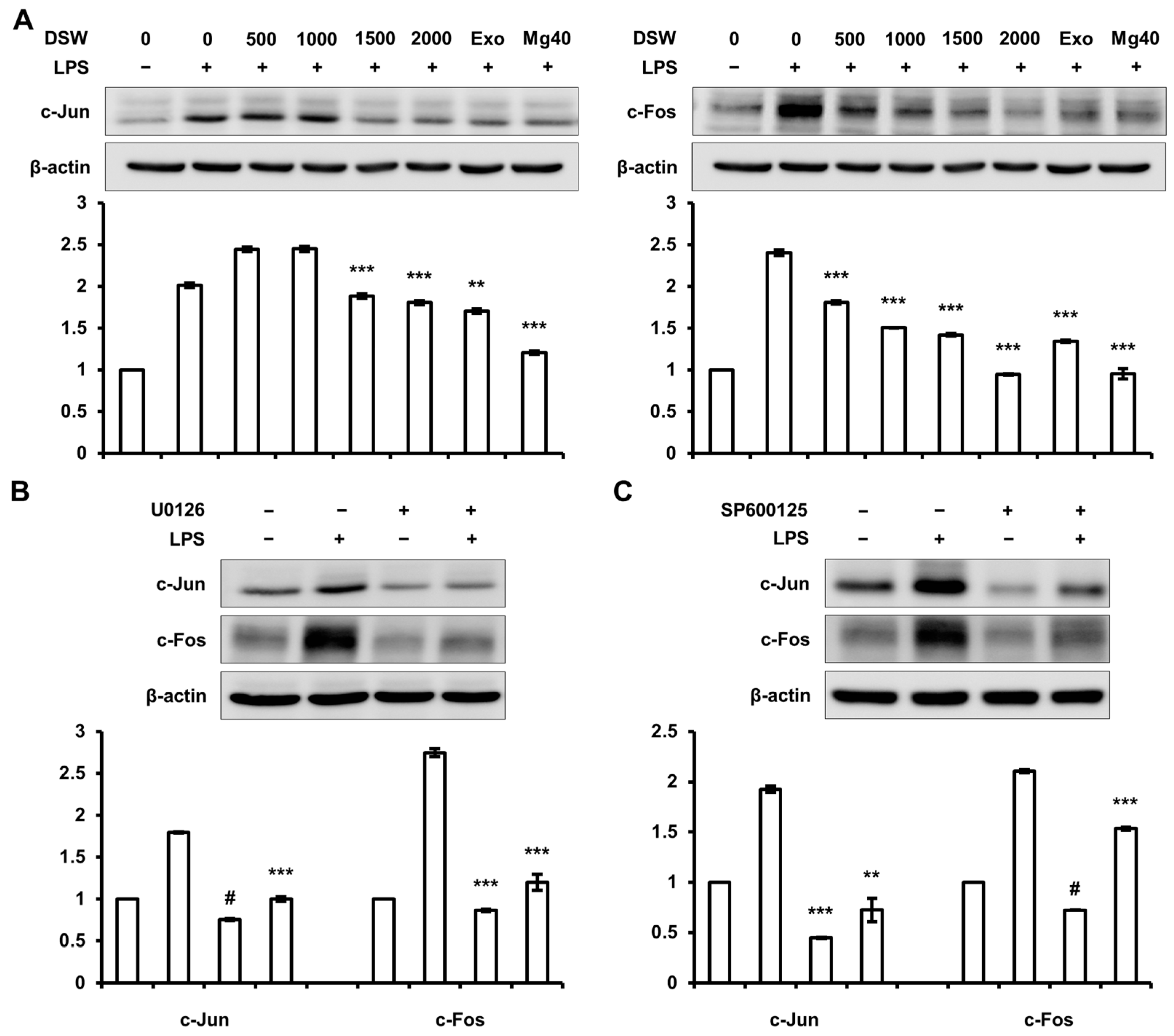
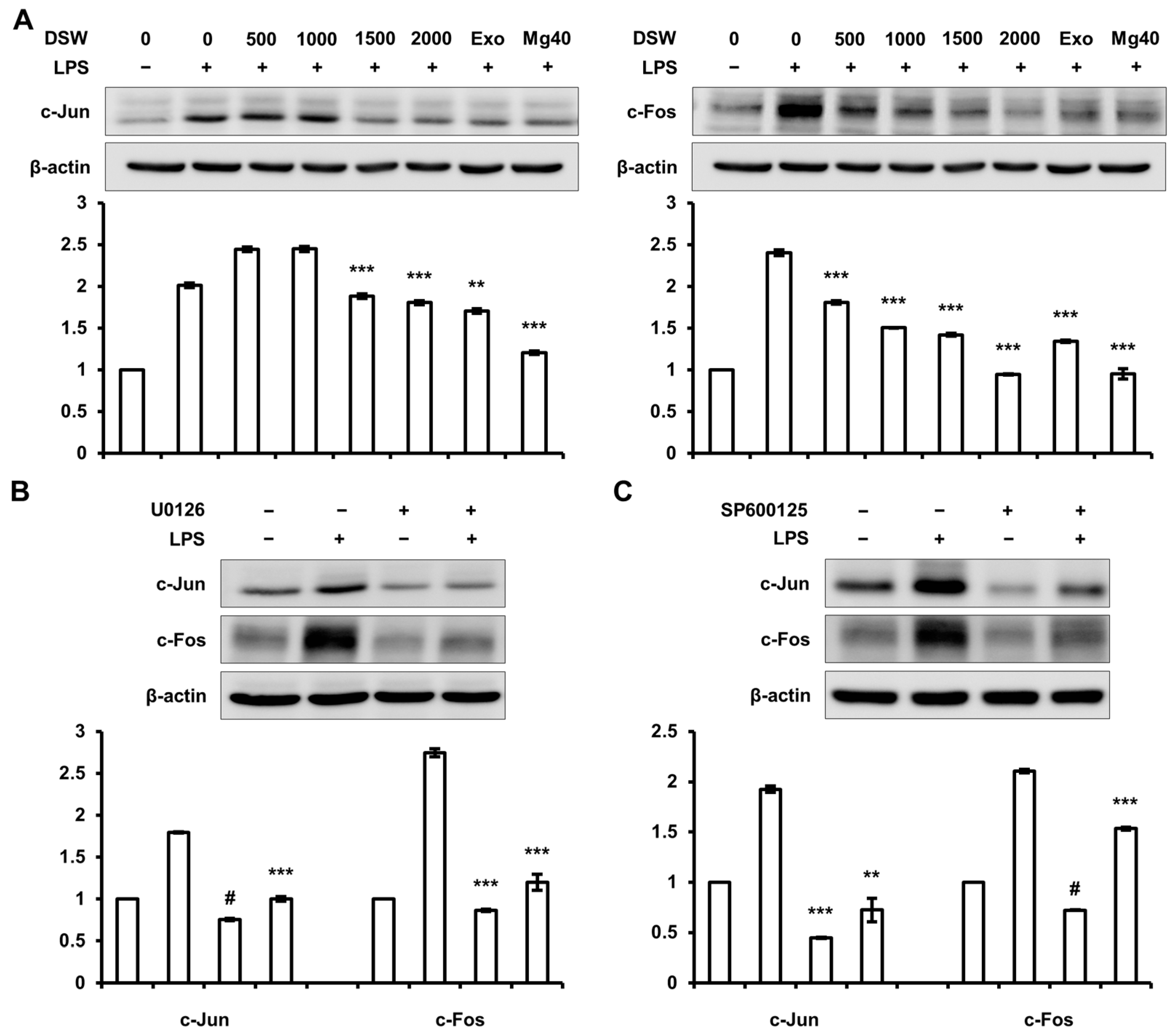
2.6. Inhibition of c-Jun and c-Fos Expressions by RDSW was Regulated by ERK 1/2 and JNK 1/2 Signaling Pathway
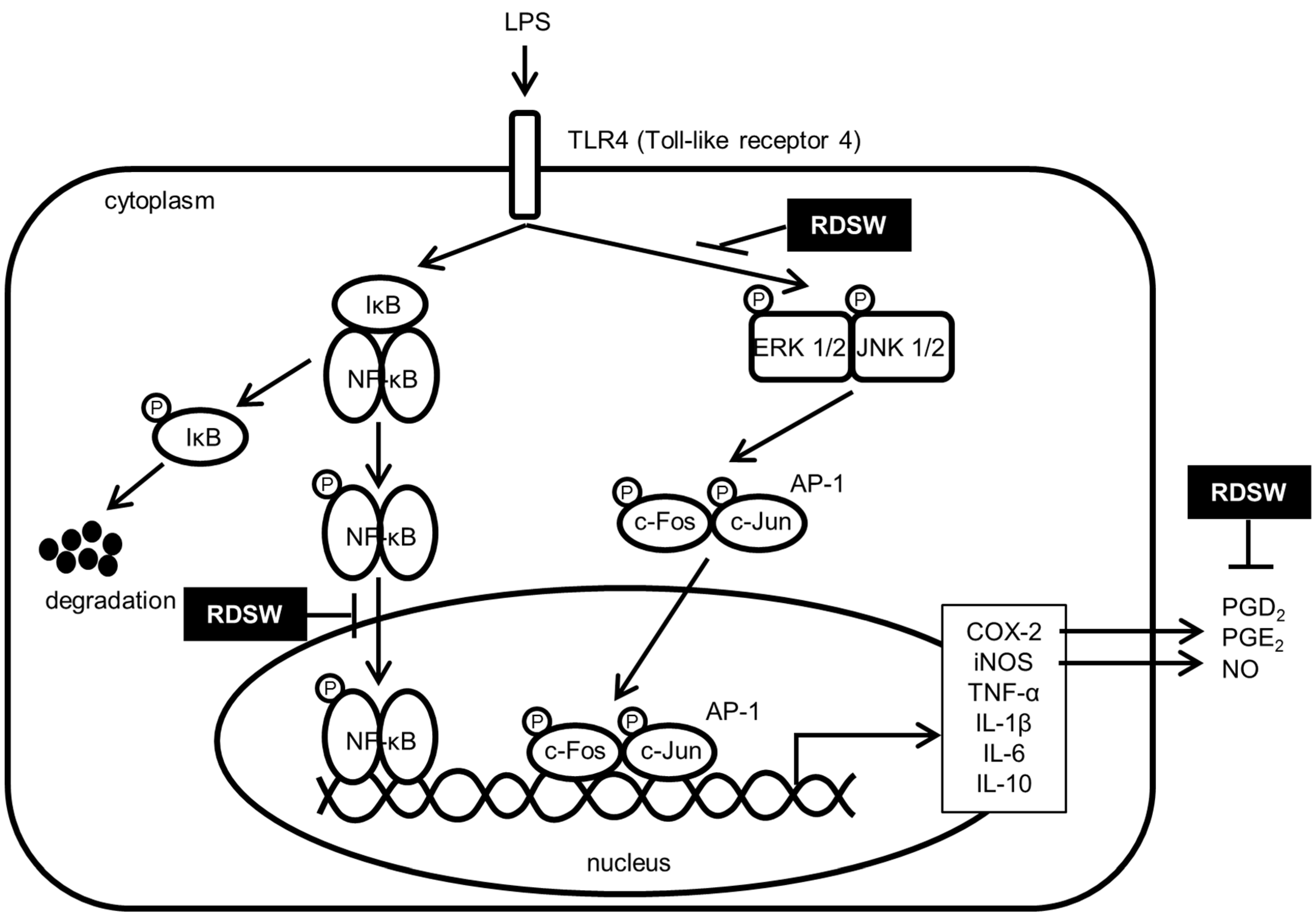
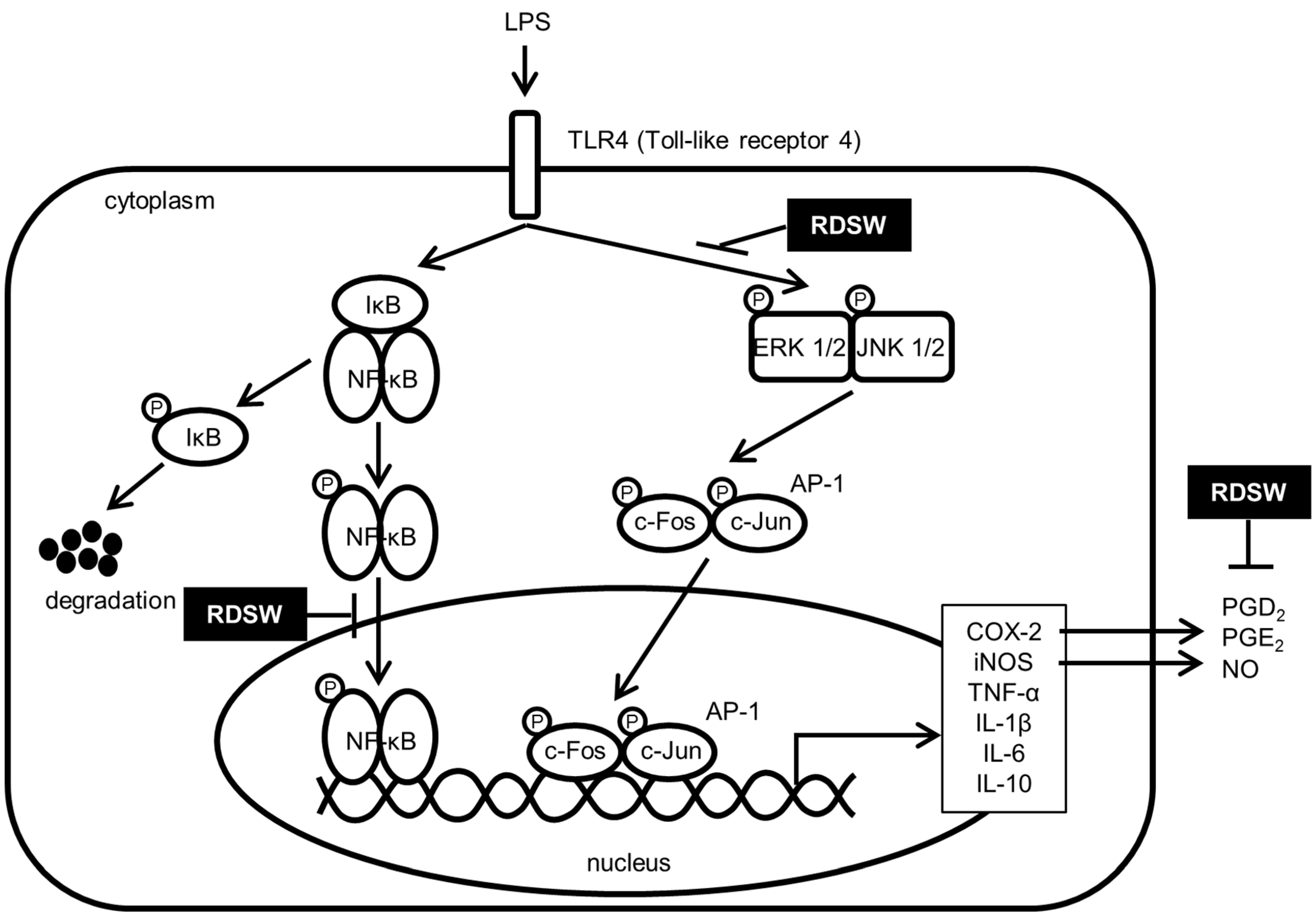
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of RDSW
4.3. Cell Treatment
4.4. Cell Viability Assay
4.5. Measurement of NO Production
4.6. Measurement of Histamine Release, PGD2, PGE2, and Various Cytokines’ Production
4.7. RNA Isolation and Quantitative Real-Time Reverse Transcriptase-Polymerase Chain Reaction (qRT-PCR)
4.8. Western Blotting
4.9. Nuclear Fractionation
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thomsen, S.F. Atopic dermatitis: Natural history, diagnosis, and treatment. ISRN Allergy 2014, 2014, 354250. [Google Scholar] [CrossRef] [PubMed]
- Bieber, T. Atopic dermatitis. Ann. Dermatol. 2010, 22, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, Y.; Guttman-Yassky, E. Immune pathways in atopic dermatitis, and definition of biomarkers through broad and targeted therapeutics. J. Clin. Med. 2015, 4, 858–873. [Google Scholar] [CrossRef] [PubMed]
- Odhiambo, J.A.; Williams, H.C.; Clayton, T.O.; Robertson, C.F.; Asher, M.I.; Group, I.P.T.S. Global variations in prevalence of eczema symptoms in children from isaac phase three. J. Allergy Clin. Immunol. 2009, 124, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Kim, J.Y.; Jeong, M.S.; Park, K.Y.; Park, K.H.; Lee, M.W.; Joo, S.S.; Seo, S.J. Effect of topical application of quercetin-3-o-(2″-gallate)-alpha-l-rhamnopyranoside on atopic dermatitis in NC/Nga mice. J. Dermatol. Sci. 2015, 77, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Bak, J.P.; Kim, Y.M.; Son, J.; Kim, C.J.; Kim, E.H. Application of concentrated deep sea water inhibits the development of atopic dermatitis-like skin lesions in NC/Nga mice. BMC Complement. Altern. Med. 2012, 12, 108. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Seo, C.S.; Ha, H.; Lee, H.; Lee, J.K.; Lee, M.Y.; Shin, H. Effect of alpinia katsumadai hayata on house dust mite-induced atopic dermatitis in NC/Nga mice. Evid.-Based Complement. Altern. Med. 2012, 2012, 705167. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Choi, S.E.; Lee, M.W.; Lee, C.S. Taxifolin glycoside inhibits dendritic cell responses stimulated by lipopolysaccharide and lipoteichoic acid. J. Pharm. Pharmacol. 2008, 60, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Kasraie, S.; Werfel, T. Role of macrophages in the pathogenesis of atopic dermatitis. Mediat. Inflamm. 2013, 2013, 942375. [Google Scholar] [CrossRef] [PubMed]
- Valledor, A.F.; Comalada, M.; Santamaria-Babi, L.F.; Lloberas, J.; Celada, A. Macrophage proinflammatory activation and deactivation: A question of balance. Adv. Immunol. 2010, 108, 1–20. [Google Scholar] [PubMed]
- Yoo, M.S.; Shin, J.S.; Choi, H.E.; Cho, Y.W.; Bang, M.H.; Baek, N.I.; Lee, K.T. Fucosterol isolated from undaria pinnatifida inhibits lipopolysaccharide-induced production of nitric oxide and pro-inflammatory cytokines via the inactivation of nuclear factor-κB and p38 mitogen-activated protein kinase in raw264.7 macrophages. Food Chem. 2012, 135, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Tajima, T.; Murata, T.; Aritake, K.; Urade, Y.; Hirai, H.; Nakamura, M.; Ozaki, H.; Hori, M. Lipopolysaccharide induces macrophage migration via prostaglandin d(2) and prostaglandin e(2). J. Pharmacol. Exp. Ther. 2008, 326, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Xue, M.Y.; Geng, Z.R.; Chen, P.Y. The suppressive effects of bursopentine (bp5) on oxidative stress and NF-κB activation in lipopolysaccharide-activated murine peritoneal macrophages. Cell Physiol. Biochem. 2012, 29, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.C.; Cho, W.K.; Jeong, Y.H.; Im, G.Y.; Kim, A.; Hwang, Y.H.; Kim, T.; Song, K.H.; Ma, J.Y. A novel herbal medicine kiom-ma exerts an anti-inflammatory effect in lps-stimulated raw 264.7 macrophage cells. Evid.-Based Complement. Altern. Med. 2012, 2012, 462383. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K. Regulation by prostaglandin e2 and histamine of angiogenesis in inflammatory granulation tissue. Yakugaku Zasshi 2003, 123, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Stander, S.; Steinhoff, M. Pathophysiology of pruritus in atopic dermatitis: An overview. Exp. Dermatol. 2002, 11, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, G.; Cohen, T.; Gengrinovitch, S.; Poltorak, Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J. 1999, 13, 9–22. [Google Scholar] [PubMed]
- Nakasone, T.; Akeda, S. The application of deep sea water in japan. UJNR Tech. Rep. 1999, 28, 69–75. [Google Scholar]
- Ha, B.G.; Shin, E.J.; Park, J.E.; Shon, Y.H. Anti-diabetic effect of balanced deep-sea water and its mode of action in high-fat diet induced diabetic mice. Mar. Drugs 2013, 11, 4193–4212. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, G.; Yamamoto, M.; Maeda, H.; Nakagawa, A.; KatareGopalrao, R.; Okada, H.; Nishimori, H.; Wariishi, S.; Toda, E.; Ogawa, H.; et al. Intake of dissolved organic matter from deep seawater inhibits atherosclerosis progression. Biochem. Biophys. Res. Commun. 2009, 387, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Kim, H.A.; Lee, S.H.; Yun, J.W. Anti-obesity and antidiabetic effects of deep sea water on ob/ob mice. Mar. Biotechnol. 2009, 11, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.G.; Park, J.E.; Shin, E.J.; Shon, Y.H. Effects of balanced deep-sea water on adipocyte hypertrophy and liver steatosis in high-fat, diet-induced obese mice. Obesity 2014, 22, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chun, S.Y.; Lee, D.H.; Lee, K.S.; Nam, K.S. Mineral-enriched deep-sea water inhibits the metastatic potential of human breast cancer cell lines. Int. J. Oncol. 2013, 43, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, S.; Nam, K.S. Protective effects of deep sea water against doxorubicininduced cardiotoxicity in H9c2 cardiac muscle cells. Int. J. Oncol. 2014, 45, 2569–2575. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Kwon, Y.S.; Kim, S.; Moon, D.S.; Kim, H.J.; Nam, K.S. Regulatory mechanism of mineral-balanced deep sea water on hypocholesterolemic effects in HepG2 hepatic cells. Biomed. Pharmacother. 2017, 86, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Chun, S.Y.; Kwon, Y.S.; Kim, S.; Nam, K.S. Deep sea water improves hypercholesterolemia and hepatic lipid accumulation through the regulation of hepatic lipid metabolic gene expression. Mol. Med. Rep. 2017, 15, 2814–2822. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, J.; Romani, A.M.; Valentin-Torres, A.M.; Luciano, A.A.; Ramirez Kitchen, C.M.; Funderburg, N.; Mesiano, S.; Bernstein, H.B. Magnesium decreases inflammatory cytokine production: A novel innate immunomodulatory mechanism. J. Immunol. 2012, 188, 6338–6346. [Google Scholar] [CrossRef] [PubMed]
- Hataguchi, Y.; Tai, H.; Nakajima, H.; Kimata, H. Drinking deep-sea water restores mineral imbalance in atopic eczema/dermatitis syndrome. Eur. J. Clin. Nutr. 2005, 59, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lu, H.; Wang, J.; Qi, X.; Liu, X.; Zhang, X. The effect of toll-like receptor 4 on macrophage cytokines during endotoxin induced uveitis. Int. J. Mol. Sci. 2012, 13, 7508–7520. [Google Scholar] [CrossRef] [PubMed]
- Orita, K.; Hiramoto, K.; Kobayashi, H.; Ishii, M.; Sekiyama, A.; Inoue, M. Inducible nitric oxide synthase (inos) and alpha-melanocyte-stimulating hormones of inos origin play important roles in the allergic reactions of atopic dermatitis in mice. Exp. Dermatol. 2011, 20, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; de Heer, H.J.; Soullie, T.; Hoogsteden, H.C.; Trottein, F.; Lambrecht, B.N. Prostaglandin d2 inhibits airway dendritic cell migration and function in steady state conditions by selective activation of the d prostanoid receptor 1. J. Immunol. 2003, 171, 3936–3940. [Google Scholar] [CrossRef] [PubMed]
- Trebino, C.E.; Stock, J.L.; Gibbons, C.P.; Naiman, B.M.; Wachtmann, T.S.; Umland, J.P.; Pandher, K.; Lapointe, J.M.; Saha, S.; Roach, M.L.; et al. Impaired inflammatory and pain responses in mice lacking an inducible prostaglandin e synthase. Proc. Natl. Acad. Sci. USA 2003, 100, 9044–9049. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.K.; Adams, D.H. Angiogenesis and chronic inflammation; the potential for novel therapeutic approaches in chronic liver disease. J. Hepatol. 2005, 42, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Valledor, A.F.; Arpa, L.; Sanchez-Tillo, E.; Comalada, M.; Casals, C.; Xaus, J.; Caelles, C.; Lloberas, J.; Celada, A. Ifn-{gamma}-mediated inhibition of mapk phosphatase expression results in prolonged mapk activity in response to m-csf and inhibition of proliferation. Blood 2008, 112, 3274–3282. [Google Scholar] [CrossRef] [PubMed]
- de Martin, R.; Vanhove, B.; Cheng, Q.; Hofer, E.; Csizmadia, V.; Winkler, H.; Bach, F.H. Cytokine-inducible expression in endothelial cells of an I kappa B alpha-like gene is regulated by NF kappa b. EMBO J. 1993, 12, 2773–2779. [Google Scholar] [PubMed]
- Mohd Nani, S.Z.; Majid, F.A.; Jaafar, A.B.; Mahdzir, A.; Musa, M.N. Potential health benefits of deep sea water: A review. Evid.-Based Complement. Altern. Med. 2016, 2016, 6520475. [Google Scholar] [CrossRef] [PubMed]
- Del Rosso, J.Q.; Bhambri, S.; Michaels, B. An aluminum magnesium hydroxide stearate-based skin barrier protection cream used for the management of eczematous dermatitis: A summary of completed studies. J. Clin. Aesth. Dermatol. 2008, 1, 18–21. [Google Scholar]
- Proksch, E.; Nissen, H.P.; Bremgartner, M.; Urquhart, C. Bathing in a magnesium-rich dead sea salt solution improves skin barrier function, enhances skin hydration, and reduces inflammation in atopic dry skin. Int. J. Dermatol. 2005, 44, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Nourbakhsh, S.M.; Rouhi-Boroujeni, H.; Kheiri, M.; Mobasheri, M.; Shirani, M.; Ahrani, S.; Karami, J.; Hafshejani, Z.K. Effect of topical application of the cream containing magnesium 2% on treatment of diaper dermatitis and diaper rash in children a clinical trial study. J. Clin. Diagn. Res. 2016, 10, WC04–WC06. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward | Reverse | Product Size (bp) |
|---|---|---|---|
| COX-2 | 5′-CCTGCTGCCCGACACCTTCA-3′ | 5′-AGCAACCCGGCCAGCAATCT-3′ | 139 |
| iNOS | 5′-CCTCCTCCACCCTACCAAGT-3′ | 5′-CACCCAAAGTGCTTCAGTCA-3′ | 119 |
| TNF-α | 5′-ATAGCTCCCAGAAAAGCAAGC-3′ | 5′-CACCCCGAAGTTCAGTAGACA-3′ | 258 |
| IL-1β | 5′-GCCTTGGGCCTCAAAGGAAAGAATC-3′ | 5′-GGAAGACACCGATTCCATGGTGAAG-3′ | 282 |
| IL-6 | 5′-TGGAGTCACAGAAGGAGTGGCTAAG-3′ | 5′-TCTGACCACAGTGAGGAATGTCCAC-3′ | 155 |
| IL-10 | 5′-CCCTTTGCTATGGTGTCCTT-3′ | 5′-TGGTTTCTCTTCCCAAGACC-3′ | 97 |
| VEGF | 5′-GTACCTCCACCATGCCAAGT-3′ | 5′-GCATTCACATCTGCTGTGCT-3′ | 340 |
| GAPDH | 5′-GTATGACTCCACTCACGGCAAA-3′ | 5′-GGTCTCGCTCCTGGAAGATG-3′ | 101 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chun, S.-Y.; Lee, K.-S.; Nam, K.-S. Refined Deep-Sea Water Suppresses Inflammatory Responses via the MAPK/AP-1 and NF-κB Signaling Pathway in LPS-Treated RAW 264.7 Macrophage Cells. Int. J. Mol. Sci. 2017, 18, 2282. https://doi.org/10.3390/ijms18112282
Chun S-Y, Lee K-S, Nam K-S. Refined Deep-Sea Water Suppresses Inflammatory Responses via the MAPK/AP-1 and NF-κB Signaling Pathway in LPS-Treated RAW 264.7 Macrophage Cells. International Journal of Molecular Sciences. 2017; 18(11):2282. https://doi.org/10.3390/ijms18112282
Chicago/Turabian StyleChun, So-Young, Kyu-Shik Lee, and Kyung-Soo Nam. 2017. "Refined Deep-Sea Water Suppresses Inflammatory Responses via the MAPK/AP-1 and NF-κB Signaling Pathway in LPS-Treated RAW 264.7 Macrophage Cells" International Journal of Molecular Sciences 18, no. 11: 2282. https://doi.org/10.3390/ijms18112282
APA StyleChun, S.-Y., Lee, K.-S., & Nam, K.-S. (2017). Refined Deep-Sea Water Suppresses Inflammatory Responses via the MAPK/AP-1 and NF-κB Signaling Pathway in LPS-Treated RAW 264.7 Macrophage Cells. International Journal of Molecular Sciences, 18(11), 2282. https://doi.org/10.3390/ijms18112282