Icariin Regulates Cellular Functions and Gene Expression of Osteoarthritis Patient-Derived Human Fibroblast-Like Synoviocytes

Abstract
1. Introduction
2. Results
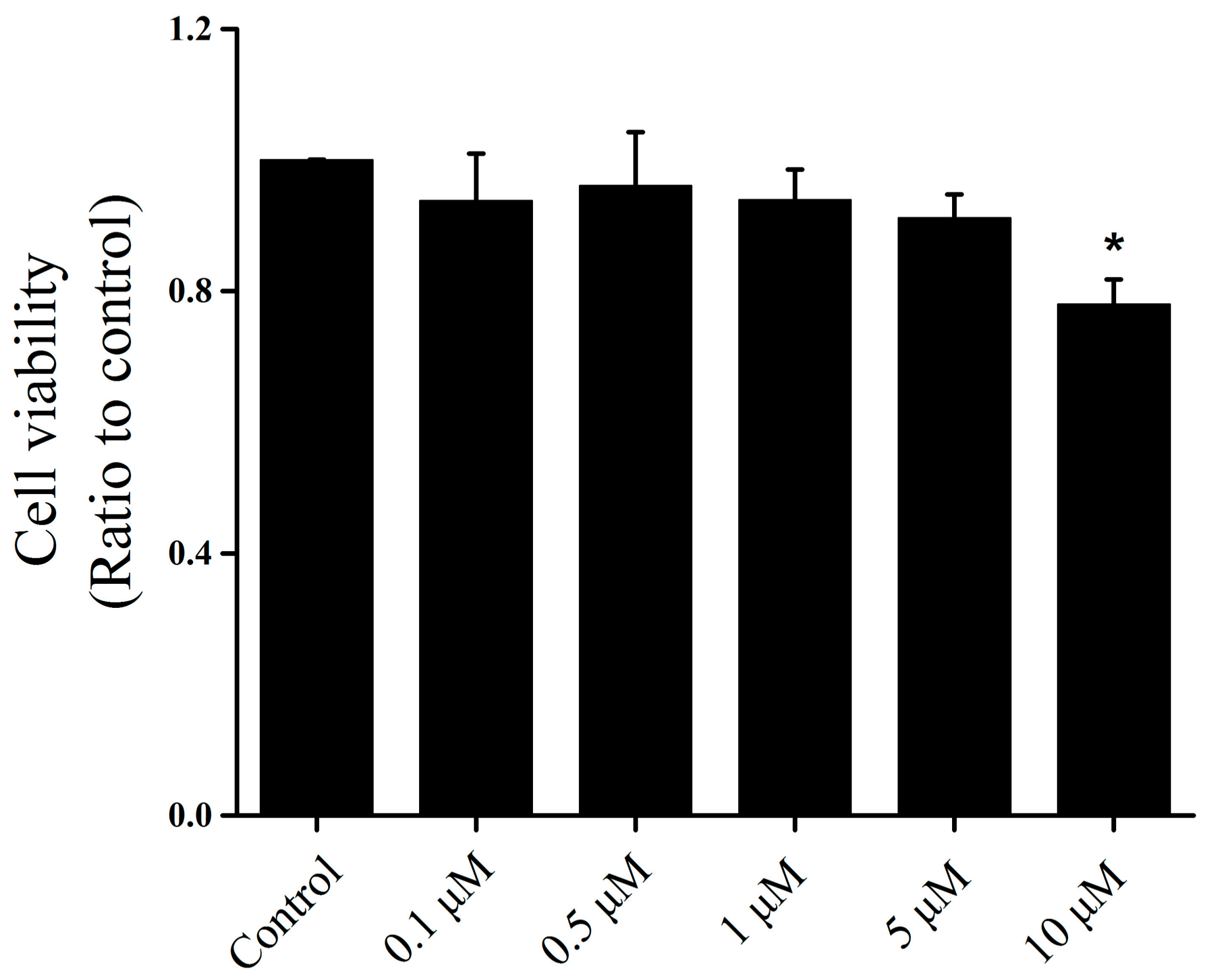
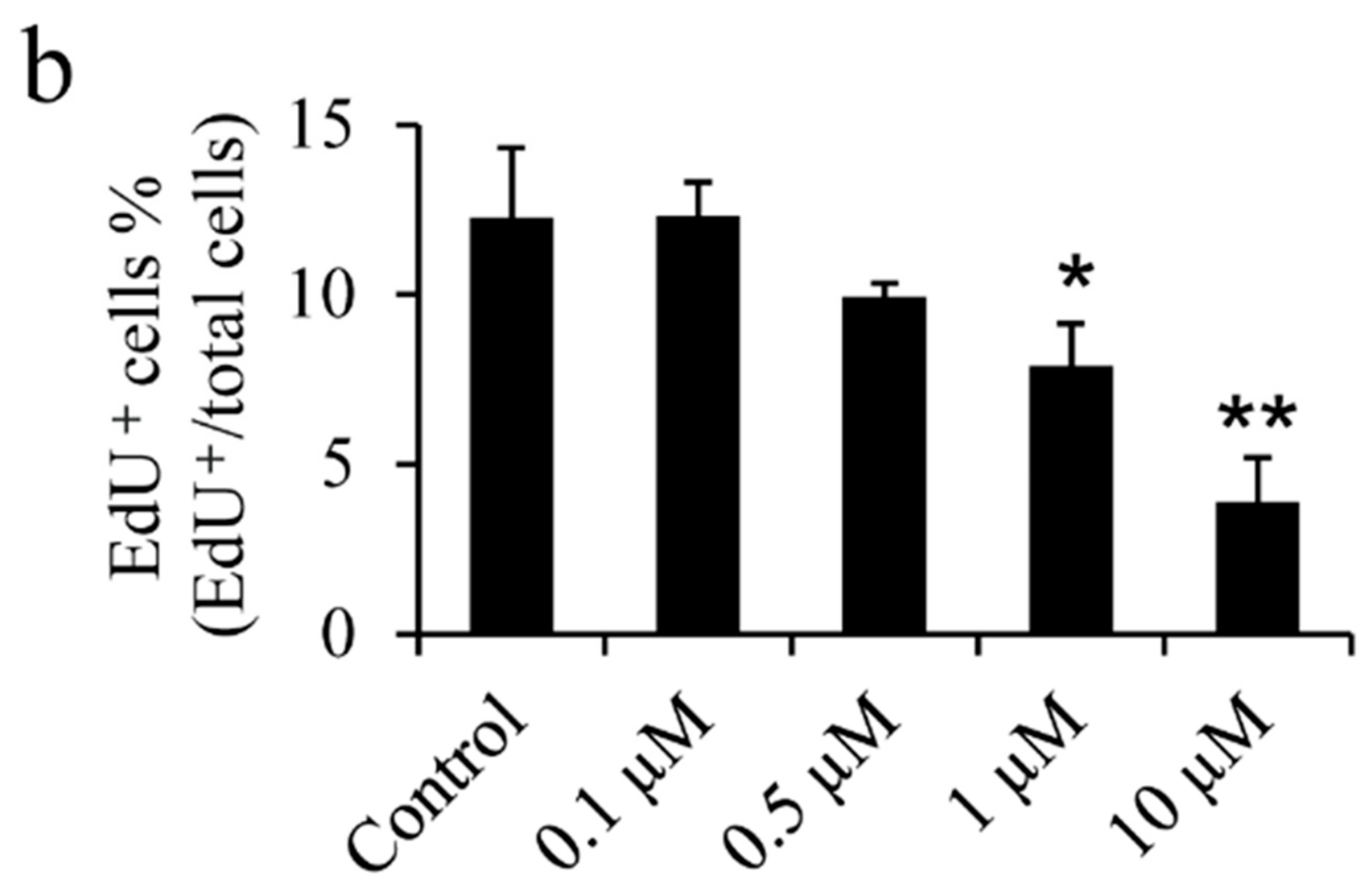
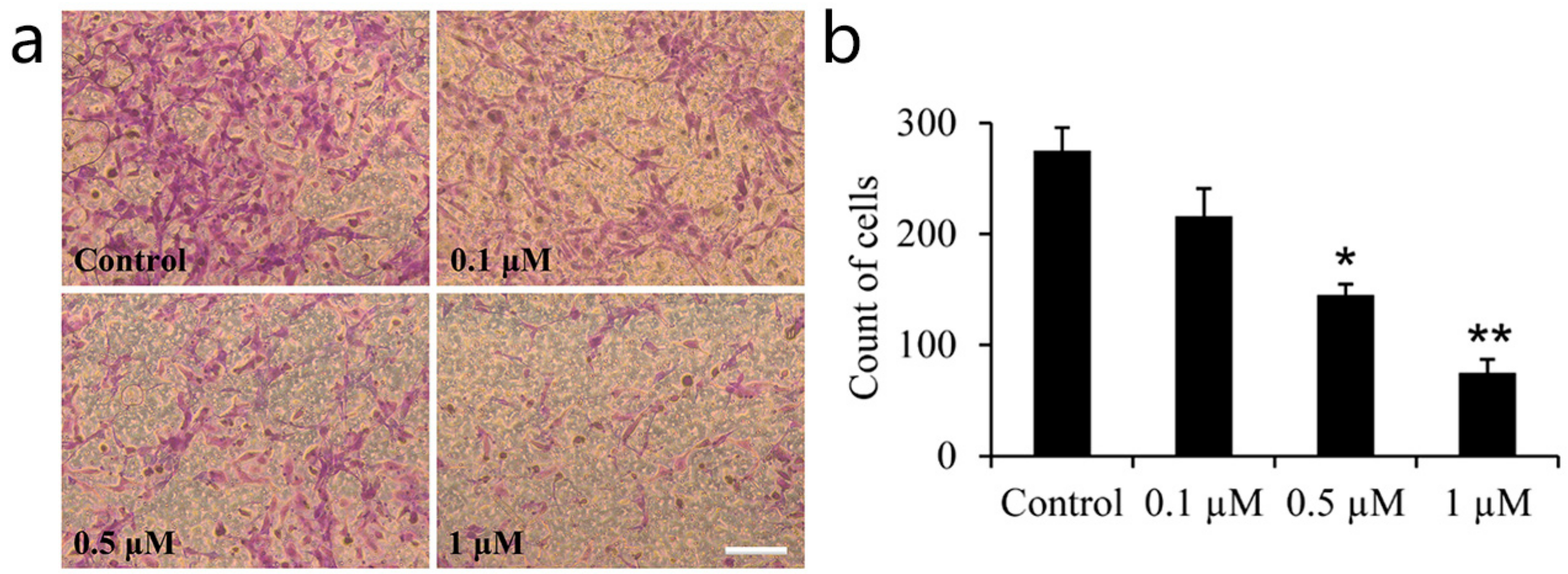
2.1. Effect of Icariin on the Osteoarthritis Fibroblast-Like Synoviocytes (OA–FLSs) Viability, Proliferation and Migration
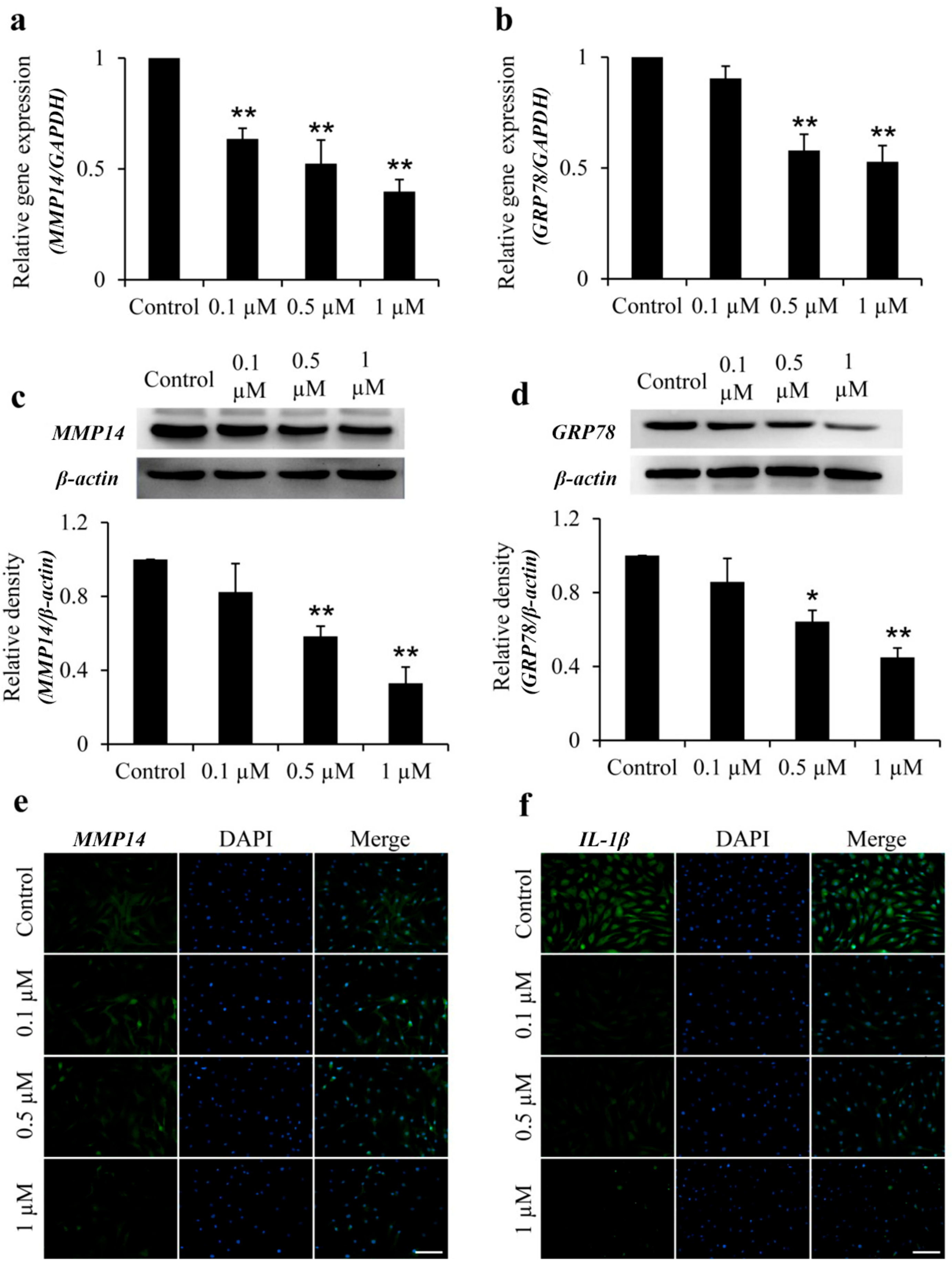
2.2. Icariin Inhibits MMP14 and GRP78 Gene and Protein Expression in OA–FLSs
2.3. Icariin Inhibits IL-1β and MMP14 Expression in OA–FLSs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cytotoxicity Assay
4.3. EdU Assay
4.4. Transwell Assay
4.5. Immunofluorescence Assay
4.6. Quantitative Real-Time Polymerase Chain Reaction
4.7. Western Blotting
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflict of Interest
References
- Guermazi, A.; Roemer, F.W.; Burstein, D.; Hayashi, D. Why radiography should no longer be considered a surrogate outcome measure for longitudinal assessment of cartilage in knee osteoarthritis. Arthritis Res. Ther. 2011, 13, 247. [Google Scholar] [CrossRef] [PubMed]
- Heidari, B. Knee osteoarthritis prevalence, risk factors, pathogenesis and features: Part I. Casp. J. Intern. Med. 2011, 2, 205–212. [Google Scholar]
- Benito, M.J.; Veale, D.J.; FitzGerald, O.; van den Berg, W.B.; Bresnihan, B. Synovial tissue inflammation in early and late osteoarthritis. Ann. Rheum. Dis. 2005, 64, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Dziak, R. Articular Cartilage and Osteoarthritis. Instr. Course Lect. 2005, 54, 465–480. [Google Scholar] [CrossRef]
- Peter, B.; Novikoff, A.B.; David, H. Electron Microscopy of the Human Synovial Membrane. J. Cell Biol. 1962, 14, 207–220. [Google Scholar]
- Pelletier, J.P.; Martel-Pelletier, J.; Abramson, S.B. Osteoarthritis, an inflammatory disease: Potential implication for the selection of new therapeutic targets. Arthritis Rheumatol. 2001, 44, 1237–1247. [Google Scholar] [CrossRef]
- Smith, M.D.; Triantafillou, S.; Parker, A.; Youssef, P.P.; Coleman, M. Synovial membrane inflammation and cytokine production in patients with early osteoarthritis. J. Rheumatol. 1997, 24, 365–371. [Google Scholar] [PubMed]
- Rollín, R.; Marco, F.; Jover, J.A.; García-Asenjo, J.A.; Rodríguez, L.; López-Durán, L.; Fernández-Gutiérrez, B. Early lymphocyte activation in the synovial microenvironment in patients with osteoarthritis: Comparison with rheumatoid arthritis patients and healthy controls. Rheumatol. Int. 2008, 28, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Kanda, V.; Bush-Joseph, C.; Verma, N.; Chubinskaya, S.; Mikecz, K.; Glant, T.T.; Malfait, A.M.; Crow, M.K.; Spear, G.T.; et al. Synovial fluid from patients with early osteoarthritis modulates fibroblast-like synoviocyte responses to toll-like receptor 4 and toll-like receptor 2 ligands via soluble CD14. Arthritis Rheumatol. 2012, 64, 2268–2277. [Google Scholar] [CrossRef] [PubMed]
- Kloesch, B.; Liszt, M.; Krehan, D.; Broell, J.; Kiener, H.; Steiner, G. High concentrations of hydrogen sulphide elevate the expression of a series of pro-inflammatory genes in fibroblast-like synoviocytes derived from rheumatoid and osteoarthritis patients. Immunol. Lett. 2012, 141, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Liu, P.; Yang, D.; Wang, F.; Yuan, Y.; Lin, Z.; Jiang, J. Interleukin-18-induced inflammatory responses in synoviocytes and chondrocytes, from osteoarthritic patients. Int. J. Mol. Med. 2012, 30, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.P.; Dibattista, J.A.; Roughley, P.; McCollum, R.; Martel-Pelletier, J. Cytokines and inflammation in cartilage degradation. Rheum. Dis. Clin. N. Am. 1993, 19, 545–568. [Google Scholar]
- Goldring, M.B. Osteoarthritis and cartilage: The role of cytokines. Curr. Rheumatol. Rep. 2000, 2, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.; Martel-Pelletier, J.; Pelletier, J.P. The role of cytokines in osteoarthritis pathophysiology. Biorheology 2002, 39, 237–246. [Google Scholar] [PubMed]
- Hassanali, S.H.; Oyoo, G.O. Osteoarthritis: A look at pathophysiology and approach to new treatments: A review. East Afr. Orthop. J. 2011, 5, 51–57. [Google Scholar]
- Hasegawa, A.; Nakahara, H.; Kinoshita, M.; Asahara, H.; Koziol, J.; Lotz, M.K. Cellular and extracellular matrix changes in anterior cruciate ligaments during human knee aging and osteoarthritis. Arthritis Res. Ther. 2013, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Seiki, M. MT1-MMP: A potent modifier of pericellular microenvironment. J. Cell. Physiol. 2006, 206, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Petrow, P.K.; Wernicke, D.; Westhoff, C.S.; Hummel, K.M.; Bräuer, R.; Kriegsmann, J.; Gromnica-Ihle, E.; Gay, R.E.; Gay, S. Characterisation of the cell type-specificity of collagenase 3 mRNA expression in comparison with membrane type 1 matrix metalloproteinase and gelatinase A in the synovial membrane in rheumatoid arthritis. Ann. Rheum. Dis. 2002, 61, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Kiener, H.P.; Watts, G.F.; Cui, Y.; Wright, J.; Thornhill, T.S.; Skold, M.; Behar, S.M.; Niederreiter, B.; Lu, J.; Cernadas, M.; et al. Synovial fibroblasts self-direct multicellular lining architecture and synthetic function in three-dimensional organ culture. Arthritis Rheumatol. 2010, 62, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Bánhegyi, G.; Baumeister, P.; Benedetti, A.; Dong, D.; Fu, Y.; Lee, A.S.; Li, J.; Mao, C.; Margittai, E.; Ni, M.; et al. Endoplasmic Reticulum Stress. Ann. N. Y. Acad. Sci. 2007, 1113, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.; Emi, M.; Tanabe, K.; Murakami, S. Role of the unfolded protein response in cell death. Apoptosis 2006, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.J.; Jiang, T.C.; Zhou, C.Y.; Yu, C.L.; Shen, Y.J.; Li, J.; Shen, Y.X. Activated macrophage-like synoviocytes are resistant to endoplasmic reticulum stress-induced apoptosis in antigen-induced arthritis. Inflamm. Res. 2014, 63, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Sze, S.C.; Tong, Y.; Ng, T.B.; Cehng, C.L.; Cheung, H.P. Herba epimedii: Anti-oxidative properties and its medical implications. Molecules 2010, 15, 7861–7870. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.Q.; Liu, B.J.; Wu, J.F.; Xu, Y.C.; Duan, X.H.; Cao, Y.X.; Dong, J.C. Icariin attenuates LPS-induced acute inflammatory responses: Involvement of PI3K/Akt and NF-κB signaling pathway. Eur. J. Pharmacol. 2010, 642, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.P.; Sheu, S.Y.; Sun, J.S.; Chen, M.H. Icariin inhibits osteoclast differentiation and bone resorption by suppression of MAPKs/NF-κB regulated HIF-1α and PGE2 synthesis. Phytomedicine 2011, 18, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.H.; Sun, J.S.; Tsai, S.W.; Sheu, S.Y.; Chen, M.H. Icariin protects murine chondrocytes from lipopolysaccharide-induced inflammatory responses and extracellular matrix degradation. Nutr. Res. 2010, 30, 57–65. [Google Scholar] [CrossRef] [PubMed]
- De Lange-Brokaar, B.J.E.; Ioanfacsinay, A.; van Osch, G.J.V.M.; Zuurmond, A.-M.; Schoones, J.; Toes, R.E.M.; Huizinga, T.W.J.; Kloppenburg, M. Synovial inflammation, immune cells and their cytokines in osteoarthritis: A review. Osteoarthr. Cartil. 2012, 20, 1484–1499. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell. Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Zwerina, J.; Redlich, K.; Polzer, K.; Joosten, L.; Krönke, G.; Distler, J.; Hess, A.; Pundt, N.; Pap, T.; Hoffmann, O.; et al. TNF-induced structural joint damage is mediated by IL-1. Proc. Natl. Acad. Sci. USA 2007, 104, 11742–11747. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.W.; Dodge, G.R. Effects of interleukin-1β and tumor necrosis factor-α on expression of matrix-related genes by cultured equine articular chondrocytes. Am. J. Vet. Res. 2000, 61, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Birkhead, J.; Sandell, L.J.; Kimura, T.; Krane, S.M. Interleukin 1 suppresses expression of cartilage-specific types II and IX collagens and increases types I and III collagens in human chondrocytes. J. Clin. Investig. 1988, 82, 2026–2037. [Google Scholar] [CrossRef] [PubMed]
- Sadouk, M.B.; Pelletier, J.P.; Tardif, G.; Kiansa, K.; Cloutier, J.M.; Martel-Pelletier, J. Human synovial fibroblasts coexpress IL-1 receptor type I and type II mRNA. The increased level of the IL-1 receptor in osteoarthritic cells is related to an increased level of the type I receptor. Lab. Investig. 1995, 73, 347–355. [Google Scholar]
- Johnson, E.O.; Charchandi, A.; Babis, G.C.; Soucacos, P.N. Apoptosis in osteoarthritis: Morphology, mechanisms, and potential means for therapeutic intervention. J. Surg. Orthop. Adv. 2008, 17, 147–152. [Google Scholar] [PubMed]
- Zhang, L.; Zhang, X.; Li, K.F.; Li, D.X.; Xiao, Y.M.; Fan, Y.J.; Zahng, X.D. Icariin promotes extracellular matrix synthesis and gene expression of chondrocytes in vitro. Phytother. Res. 2012, 26, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Baillymaitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. Int. J. Endocrinol. Metab. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.V.; Ellerby, H.M.; Bredesen, D.E. Coupling endoplasmic reticulum stress to the cell death program. J. Biol. Chem. 2001, 276, 33869–33874. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cao, Y.; Yang, X.; Shan, P.; Liu, H. Tauroursodeoxycholic acid suppresses endoplasmic reticulum stress in the chondrocytes of patients with osteoarthritis. Int. J. Mol. Med. 2015, 36, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, Z.; Xue, R.; Singh, G.K.; Lv, Y.; Shi, K.; Cai, K.; Deng, L.; Yang, L. TGF-β1 promoted MMP-2 mediated wound healing of anterior cruciate ligament fibroblasts through NF-κB. Connect. Tissue Res. 2011, 52, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Singh, G.K.; Zhang, F.; Wang, P.; Liu, W.; Zhong, L.; Yang, L. Comparative study of normal and rheumatoid arthritis fibroblast-like synoviocytes proliferation under cyclic mechanical stretch: Role of prostaglandin E2. Connect. Tissue Res. 2012, 53, 246–254. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (Forward/Reverse) | Product Size (bp) |
|---|---|---|
| MMP14 | 5′-CCGATGTGGTGTTCCAGACA-3′ | 153 |
| 5′-CGTATGTGGCATACTCGCC-3′ | ||
| GRP78 | 5′-CTTAAGCTGCCACCATGAAG | 256 |
| CTCTCCCTGGTGGCCGCG-3′ | ||
| 5′-AGGCCTCGAGCTACAACT | ||
| CATCTTTTTCTGCTGT-3 | ||
| GAPDH | 5′-AAATTCCATGGCACCGTCAAGGCT-3′ | 183 |
| 5′-CTCATGGTTCACACCCATGACGAA-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, L.; Zhang, Y.; Chen, N.; Yang, L. Icariin Regulates Cellular Functions and Gene Expression of Osteoarthritis Patient-Derived Human Fibroblast-Like Synoviocytes. Int. J. Mol. Sci. 2017, 18, 2656. https://doi.org/10.3390/ijms18122656
Pan L, Zhang Y, Chen N, Yang L. Icariin Regulates Cellular Functions and Gene Expression of Osteoarthritis Patient-Derived Human Fibroblast-Like Synoviocytes. International Journal of Molecular Sciences. 2017; 18(12):2656. https://doi.org/10.3390/ijms18122656
Chicago/Turabian StylePan, Lianhong, Yonghui Zhang, Na Chen, and Li Yang. 2017. "Icariin Regulates Cellular Functions and Gene Expression of Osteoarthritis Patient-Derived Human Fibroblast-Like Synoviocytes" International Journal of Molecular Sciences 18, no. 12: 2656. https://doi.org/10.3390/ijms18122656
APA StylePan, L., Zhang, Y., Chen, N., & Yang, L. (2017). Icariin Regulates Cellular Functions and Gene Expression of Osteoarthritis Patient-Derived Human Fibroblast-Like Synoviocytes. International Journal of Molecular Sciences, 18(12), 2656. https://doi.org/10.3390/ijms18122656