miR-34a Regulates Sperm Motility in Zebrafish


Abstract
:1. Introduction
2. Results
2.1. Expression of miR-34a during Zebrafish Testis Development
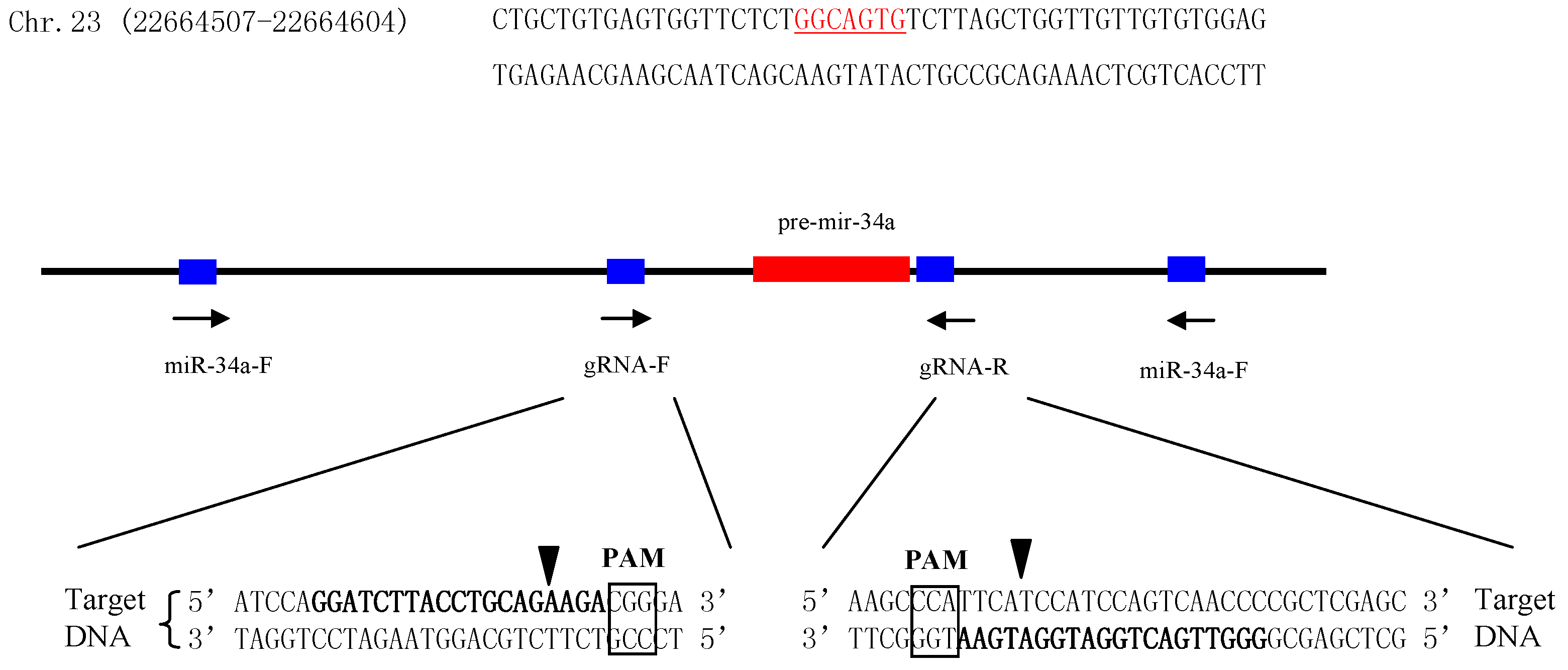
2.2. Establishment of miR-34a Knockout Zebrafish Model
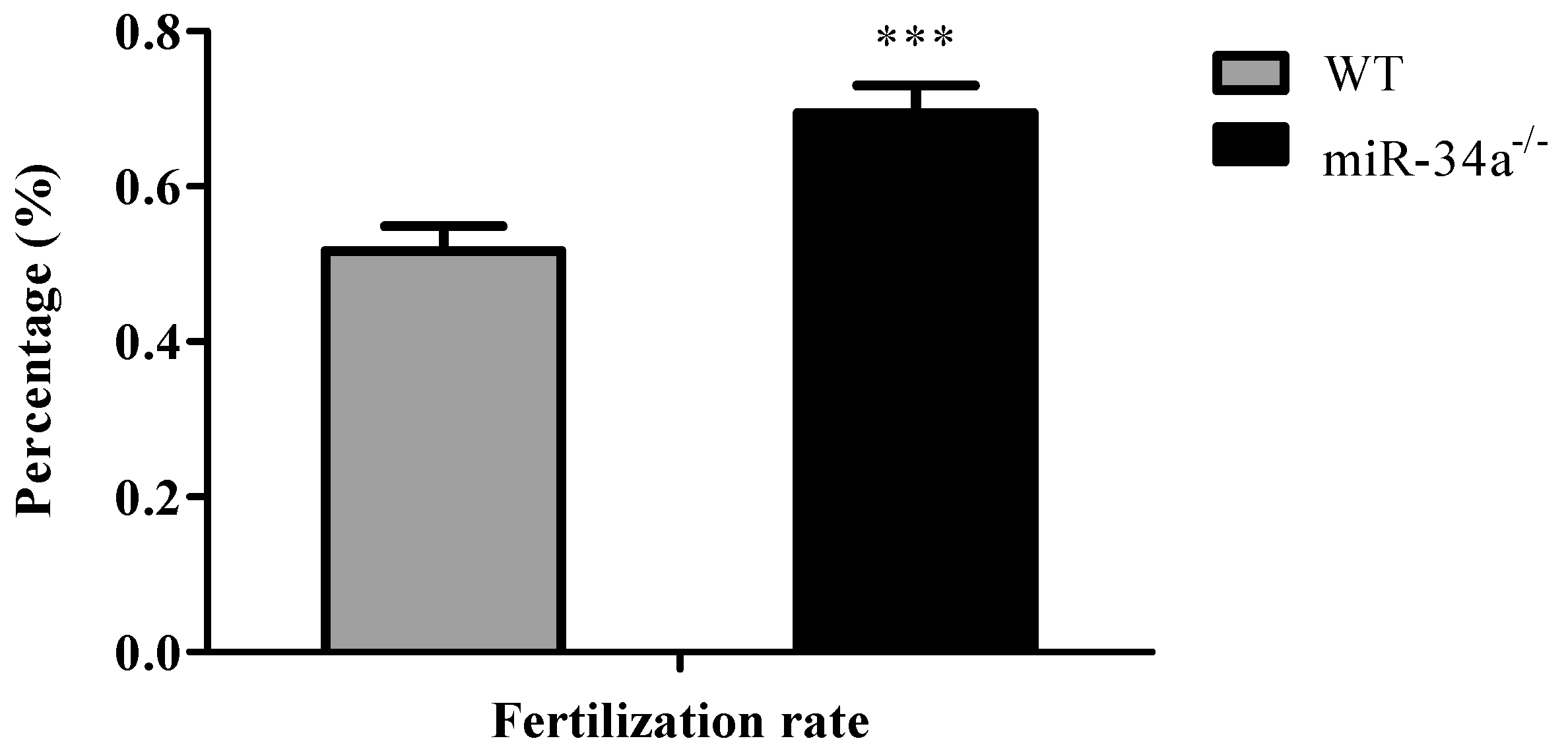
2.3. Effects of miR-34a Knockout on Zebrafish Sperm Motility
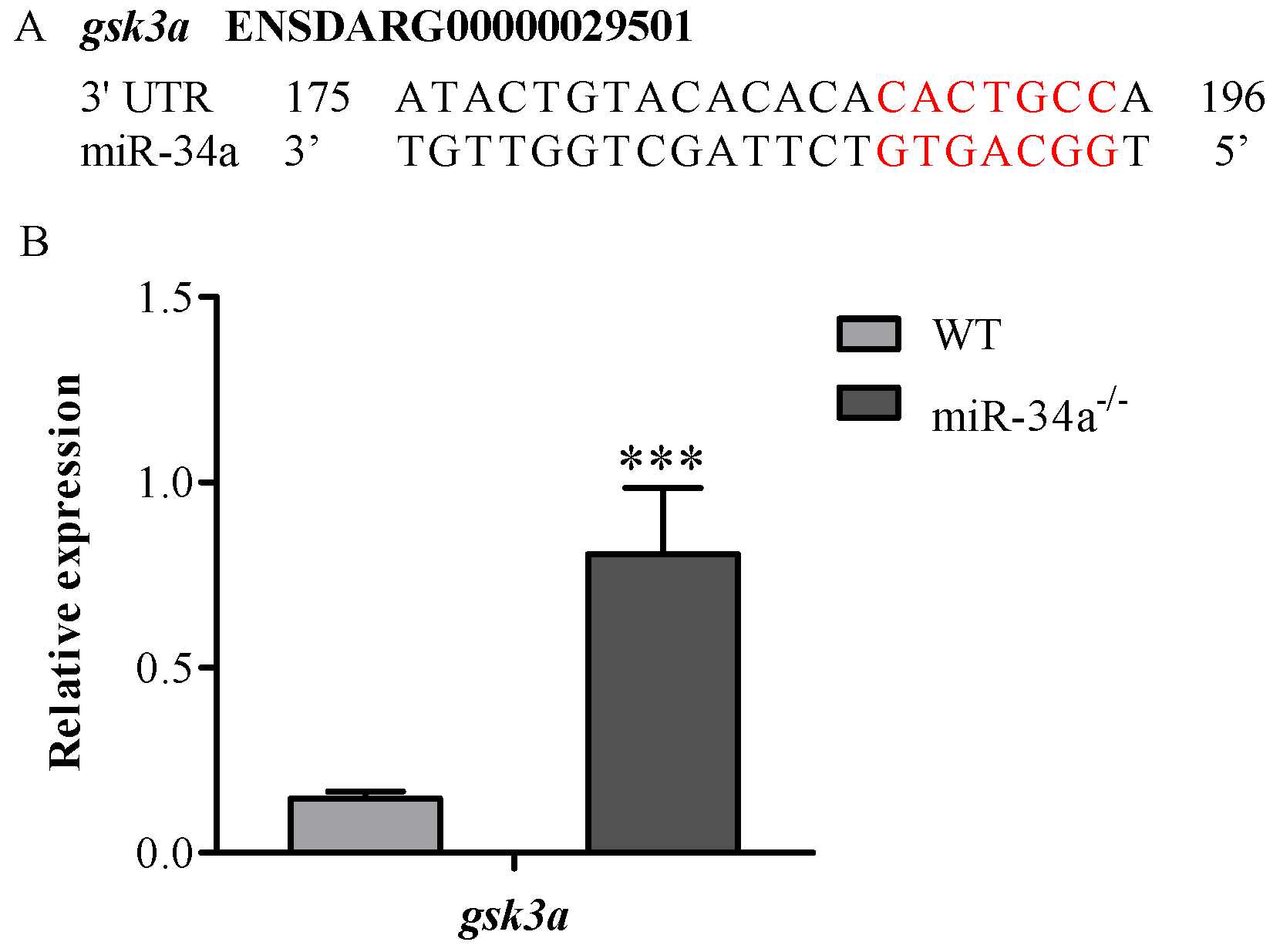
2.4. Potential Target Genes of miR-34a
2.5. Luciferase Reporter Assays
3. Discussion
4. Materials and Methods
4.1. Zebrafish Strain
4.2. Design of CRISPR/Cas9 Target Site and Single Guide RNA (sgRNA) Synthesis
4.3. Capped mRNA Synthesis
4.4. Microinjection, Mutation Analyses, and Mutant Lines Establishment
4.5. Evaluation of Sperm Motility and Fertilization Rate
4.6. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR) Analysis
4.7. Luciferase Reporter Assay
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, D.H.; Hu, J.R.; Wang, L.Y.; Hu, Y.J.; Tan, F.Q.; Zhou, H.; Shao, J.Z.; Yang, W.X. The Apoptotic Function Analysis of p53, Apaf1, Caspase3 and Caspase7 during the Spermatogenesis of the Chinese Fire-Bellied Newt Cynops orientalis. PLoS ONE 2012, 7, e39920. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P.T. Germ cell-somatic cell interactions during spermatogenesis. Reproduction 2003, 61, 91–101. [Google Scholar] [PubMed]
- Costoya, J.A.; Hobbs, R.M.; Barna, M.; Cattoretti, G.; Manova, K.; Sukhwani, M.; Orwig, K.E.; Wolgemuth, D.J.; Pandolfi, P.P. Essential role of Plzf in maintenance of spermatogonial stem cells. Nat. Genet. 2004, 36, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Naughton, C.K.; Jain, S.; Strickland, A.M.; Gupta, A.; Milbrandt, J. Glial cell-line derived neurotrophic factor-mediated RET signaling regulates spermatogonial stem cell fate. Biol. Reprod. 2006, 74, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Pang, A.; Rennert, O.M. Protein acetylation and spermatogenesis. Reprod. Syst. Sex. Disord. Curr. Res. 2013. [Google Scholar] [CrossRef]
- Shima, J.E.; McLean, D.J.; McCarrey, J.R.; Griswold, M.D. The murine testicular transcriptome: Characterizing gene expression in the testis during the progression of spermatogenesis. Biol. Reprod. 2004, 71, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, M.C.; Santangeli, S.; Maradonna, F.; Gioacchini, G.; Verdenelli, C.; Gallego, V.; Penaranda, D.S.; Tveiten, H.; Perez, L.; Carnevali, O.; et al. Effect of the probiotic Lactobacillus rhamnosus on the expression of genes involved in European eel spermatogenesis. Theriogenology 2015, 84, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, N.; Amiri, I.; Farimani, M.; Najafi, R.; Hadeie, J. Correlation between sperm parameters and sperm DNA fragmentation in fertile and infertile men. Int. J. Reprod. Med. 2008, 6, 13–18. [Google Scholar]
- Labas, V.; Grasseau, I.; Cahier, K.; Gargaros, A.; Harichaux, G.; Teixeira-Gomes, A.P.; Alves, S.; Bourin, M.; Gerard, N.; Blesbois, E. Data for chicken semen proteome and label free quantitative analyses displaying sperm quality biomarkers. Data Brief 2014, 1, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Montjean, D.; Zini, A.; Ravel, C.; Belloc, S.; Dalleac, A.; Copin, H.; Boyer, P.; McElreavey, K.; Benkhalifa, M. Sperm global DNA methylation level: Association with semen parameters and genome integrity. Andrology 2015, 3, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Sassone-Corsi, P. CREM: A master-switch regulating the balance between differentiation and apoptosis in male germ cells. Mol. Reprod. Dev. 2000, 56 (Suppl. 2), 228–229. [Google Scholar] [CrossRef]
- Sapiro, R.; Kostetskii, I.; Olds-Clarke, P.; Gerton, G.L.; Radice, G.L.; Strauss, I.J. Male infertility, impaired sperm motility, and hydrocephalus in mice deficient in sperm-associated antigen 6. Mol. Cell. Biol. 2002, 22, 6298–6305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kostetskii, I.; Tang, W.; Haig-Ladewig, L.; Sapiro, R.; Wei, Z.; Patel, A.M.; Bennett, J.; Gerton, G.L.; Moss, S.B.; et al. Deficiency of SPAG16L causes male infertility associated with impaired sperm motility. Biol. Reprod. 2006, 74, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Danshina, P.V.; Geyer, C.B.; Dai, Q.; Goulding, E.H.; Willis, W.D.; Kitto, G.B.; McCarrey, J.R.; Eddy, E.M.; O’Brien, D.A. Phosphoglycerate kinase 2 (PGK2) is essential for sperm function and male fertility in mice. Biol. Reprod. 2010, 82, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Robles, V.; Herraez, P.; Labbe, C.; Cabrita, E.; Psenicka, M.; Valcarce, D.G.; Riesco, M.F. Molecular basis of spermatogenesis and sperm quality. Gen. Comp. Endocrinol. 2017, 245, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Bassett, A.R.; Azzam, G.; Wheatley, L.; Tibbit, C.; Rajakumar, T.; McGowan, S.; Stanger, N.; Ewels, P.A.; Taylor, S.; Ponting, C.P.; et al. Understanding functional miRNA-target interactions in vivo by site-specific genome engineering. Nat. Commun. 2014, 5, 4640. [Google Scholar] [CrossRef] [PubMed]
- Shcherbata, H.R.; Hatfield, S.; Ward, E.J.; Reynolds, S.; Fischer, K.A.; Ruohola-Baker, H. The MicroRNA pathway plays a regulatory role in stem cell division. Cell Cycle 2006, 5, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Korpal, M.; Lee, E.S.; Hu, G.; Kang, Y. The miR-200 family inhibits epithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin transcriptional repressors ZEB1 and ZEB2. J. Boil. Chem. 2008, 283, 14910–14914. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.M.; Byrom, M.W.; Shelton, J.; Ford, L.P. Antisense inhibition of human miRNAs and indications for an involvement of miRNA in cell growth and apoptosis. Nucleic Acids Res. 2005, 33, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Presslauer, C.; Bizuayehu, T.T.; Kopp, M.; Fernandes, J.M.O.; Babiak, I. Dynamics of miRNA transcriptome during gonadal development of zebrafish. Sci. Rep. 2017, 7, 43850. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, V.R.; Kasimanickam, R.K. Differential expression of microRNAs in sexually immature and mature canine testes. Theriogenology 2015, 83, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Wang, Y.; Fu, J.; Cheng, X.; Miao, S.; Wang, L. Cyclin T2: A novel miR-15a target gene involved in early spermatogenesis. FEBS Lett. 2011, 585, 2493–2500. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bao, J.; Wang, L.; Hu, Y.; Xu, C. MicroRNA-184 downregulates nuclear receptor corepressor 2 in mouse spermatogenesis. BMC Dev. Boil. 2011, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Navarro, B.; Perez, G.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A sperm ion channel required for sperm motility and male fertility. Nature 2001, 413, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Jamsai, D.; O’Connor, A.E.; Borg, C.; Clark, B.J.; Whisstock, J.C.; Field, M.C.; Adams, V.; Ishikawa, T.; Aitken, R.J.; et al. RAB-like 2 has an essential role in male fertility, sperm intra-flagellar transport, and tail assembly. PLoS Genet. 2012, 8, e1002969. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, R.; Goswami, S.; Dudiki, T.; Popkie, A.P.; Phiel, C.J.; Kline, D.; Vijayaraghavan, S. Targeted disruption of glycogen synthase kinase 3A (GSK3A) in mice affects sperm motility resulting in male infertility. Biol. Reprod. 2015, 92, 65. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.Y. CFTR gene and male fertility. Mol. Hum. Reprod. 1998, 4, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Curry, E.; Safranski, T.J.; Pratt, S.L. Differential expression of porcine sperm microRNAs and their association with sperm morphology and motility. Theriogenology 2011, 76, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.H.; Zhou, Q.Z.; Lyu, X.M.; Zhu, T.; Chen, Z.J.; Chen, M.K.; Xia, H.; Wang, C.Y.; Qi, T.; Li, X.; et al. The Expression of Cysteine-Rich Secretory Protein 2 (CRISP2) and Its Specific Regulator miR-27b in the Spermatozoa of Patients with Asthenozoospermia. Biol. Reprod. 2015, 92, 28. [Google Scholar] [CrossRef] [PubMed]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.L.; Jin, F.; Ye, X.; Zhu, L.; Yang, J.S.; Yang, W.J. Expression profiles of miRNAs and involvement of miR-100 and miR-34 in regulation of cell cycle arrest in Artemia. Biochem. J. 2015, 470, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Disayabutr, S.; Kim, E.K.; Cha, S.I.; Green, G.; Naikawadi, R.P.; Jones, K.D.; Golden, J.A.; Schroeder, A.; Matthay, M.A.; Kukreja, J.; et al. miR-34 miRNAs Regulate Cellular Senescence in Type II Alveolar Epithelial Cells of Patients with Idiopathic Pulmonary Fibrosis. PLoS ONE 2016, 11, e0158367. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.C.; Wentzel, E.A.; Kent, O.A.; Ramachandran, K.; Mullendore, M.; Lee, K.H.; Feldmann, G.; Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J.; et al. Transactivation of miR-34a by p53 broadly influences gene expression and promotes apoptosis. Mol. Cell 2007, 26, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Bouhallier, F.; Allioli, N.; Lavial, F.; Chalmel, F.; Perrard, M.H.; Durand, P.; Samarut, J.; Pain, B.; Rouault, J.P. Role of miR-34c microRNA in the late steps of spermatogenesis. RNA 2010, 16, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.W.; Bao, J.Q.; Kim, M.; Yuan, S.Q.; Tang, C.; Zheng, H.L.; Mastick, G.S.; Xu, C.; Yan, W. Two miRNA clusters, miR-34b/c and miR-449, are essential for normal brain development, motile ciliogenesis, and spermatogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, E2851–E2857. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Ma’ayan, A.; Jenkins, S.L.; Neves, S.; Hasseldine, A.; Grace, E.; Dubin-Thaler, B.; Eungdamrong, N.J.; Weng, G.; Ram, P.T.; Rice, J.J.; et al. Formation of regulatory patterns during signal propagation in a Mammalian cellular network. Science 2005, 309, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Boehm, M.; Slack, F. A developmental timing microRNA and its target regulate life span in C. elegans. Science 2005, 310, 1954–1957. [Google Scholar] [CrossRef] [PubMed]
- Hebert, S.S.; Horre, K.; Nicolai, L.; Papadopoulou, A.S.; Mandemakers, W.; Silahtaroglu, A.N.; Kauppinen, S.; Delacourte, A.; De Strooper, B. Loss of microRNA cluster miR-29a/b-1 in sporadic Alzheimer’s disease correlates with increased BACE1/beta-secretase expression. Proc. Natl. Acad. Sci. USA 2008, 105, 6415–6420. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [PubMed]
- Agostini, M.; Knight, R.A. miR-34: From bench to bedside. Oncotarget 2014, 5, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Zhou, X.; Chen, X.; Hu, D.N.; Dong, X.D.; Wang, J.; Lu, F.; Tu, L.; Qu, J. MicroRNA-34a inhibits uveal melanoma cell proliferation and migration through downregulation of c-Met. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.J.; Weng, S.L.; Hsieh, J.Y.; Wang, T.Y.; Chang, M.D.; Wang, H.W. MicroRNA-34a modulates genes involved in cellular motility and oxidative phosphorylation in neural precursors derived from human umbilical cord mesenchymal stem cells. BMC Med. Genom. 2011, 4, 65. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Martello, G.; Piccolo, S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol. 2010, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Bachoo, R.; Zhang, C.L. MicroRNA-146a inhibits glioma development by targeting Notch1. Mol. Cell. Boil. 2011, 31, 3584–3592. [Google Scholar] [CrossRef] [PubMed]
- Subtelny, A.O.; Eichhorn, S.W.; Chen, G.R.; Sive, H.; Bartel, D.P. Poly (A)-tail profiling reveals an embryonic switch in translational control. Nature 2014, 508, 66–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado de Llera, A.; Martin-Hidalgo, D.; Gil, M.C.; Garcia-Marin, L.J.; Bragado, M.J. New insights into transduction pathways that regulate boar sperm function. Theriogenology 2016, 85, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; Li, B.; Huang, C.; Wei, Z.; Zhou, Y.; Liu, J.; Zhang, H. miR-34a is Involved in the Decrease of ATP Contents Induced by Resistin Through Target on ATP5S in HepG2 Cells. Biochem. Genet. 2015, 53, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Bukeirat, M.; Sarkar, S.N.; Hu, H.; Quintana, D.D.; Simpkins, J.W.; Ren, X. miR-34a regulates blood-brain barrier permeability and mitochondrial function by targeting cytochrome c. J. Cereb. Blood Flow Metab. 2016, 36, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Nam, K.H. Regulation of brassinosteroid signaling by a GSK3/SHAGGY-like kinase. Science 2002, 295, 1299–1301. [Google Scholar] [PubMed]
- Xiong, S.T.; Wu, J.J.; Jing, J.; Huang, P.P.; Li, Z.; Mei, J.; Gui, J.F. Loss of stat3 function leads to spine malformation and immune disorder in zebrafish. Sci. Bull. 2017, 62, 185–196. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, W.; Xie, B.; Gui, J.F.; Mei, J. Beneficial effect and potential molecular mechanism of chloroquine on sperm motility and fertilizing ability in yellow catfish. Aquaculture 2017, 468, 307–313. [Google Scholar] [CrossRef]
- Jing, J.; Xiong, S.; Li, Z.; Wu, J.; Zhou, L.; Gui, J.F.; Mei, J. A feedback regulatory loop involving p53/miR-200 and growth hormone endocrine axis controls embryo size of zebrafish. Sci. Rep. 2015, 5, 15906. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Wu, J.; Xiong, S.; Zhang, X.; Zhang, J.; Mei, J. MicroRNA-203a regulates fast muscle differentiation by targeting dmrt2a in zebrafish embryos. Gene 2017, 625, 49–54. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Sequences (5′–3′) | PAM |
|---|---|---|
| gRNA-F | GGATCTTACCTGCAGAAGAC | GGG |
| gRNA-R | GGGTTGACTGGATGGATGAA | TGG |
| g-miR-34a-F | GGACTTGTGACTGCTGTAATTCC | |
| g-miR-34a-R | CTAATGAAATGACTCAGGCTAC |
| Primers | Sequences (5′-3′) | Size of the Products (bp) |
|---|---|---|
| miR-203a stem loop | CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAG | |
| miR-34a-F | GCGTGGCAGTGTCTTAGCTG | 57 |
| miR-34a-R | ACTGGTGTCGTGGAGTCGGC | |
| U6-F | TGCTCGCTACGGTGGCACA | 111 |
| U6-R | AAAACAGCAATATGGAGCGC | |
| gsk3a-F | TAAAGGGGCACAAGAGGTTC | 187 |
| gsk3a-R | TGTCGCTGATAGATATTTCGTC | |
| β-actin-F | CGAGCAGGAGATGGGAACC | 102 |
| β-actin-R | CAACGGAAACGCTCATTGC |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, W.; Xie, B.; Xiong, S.; Liang, X.; Gui, J.-F.; Mei, J. miR-34a Regulates Sperm Motility in Zebrafish. Int. J. Mol. Sci. 2017, 18, 2676. https://doi.org/10.3390/ijms18122676
Guo W, Xie B, Xiong S, Liang X, Gui J-F, Mei J. miR-34a Regulates Sperm Motility in Zebrafish. International Journal of Molecular Sciences. 2017; 18(12):2676. https://doi.org/10.3390/ijms18122676
Chicago/Turabian StyleGuo, Wenjie, Binyue Xie, Shuting Xiong, Xufang Liang, Jian-Fang Gui, and Jie Mei. 2017. "miR-34a Regulates Sperm Motility in Zebrafish" International Journal of Molecular Sciences 18, no. 12: 2676. https://doi.org/10.3390/ijms18122676
APA StyleGuo, W., Xie, B., Xiong, S., Liang, X., Gui, J.-F., & Mei, J. (2017). miR-34a Regulates Sperm Motility in Zebrafish. International Journal of Molecular Sciences, 18(12), 2676. https://doi.org/10.3390/ijms18122676