
Transcriptional and Behavioral Responses of Zebrafish Larvae to Microcystin-LR Exposure


Abstract
:1. Introduction
2. Results
2.1. Alterations of Protein Phosphatase 2A Activity in Zebrafish Larvae Following Exposure to Microcystin-LR
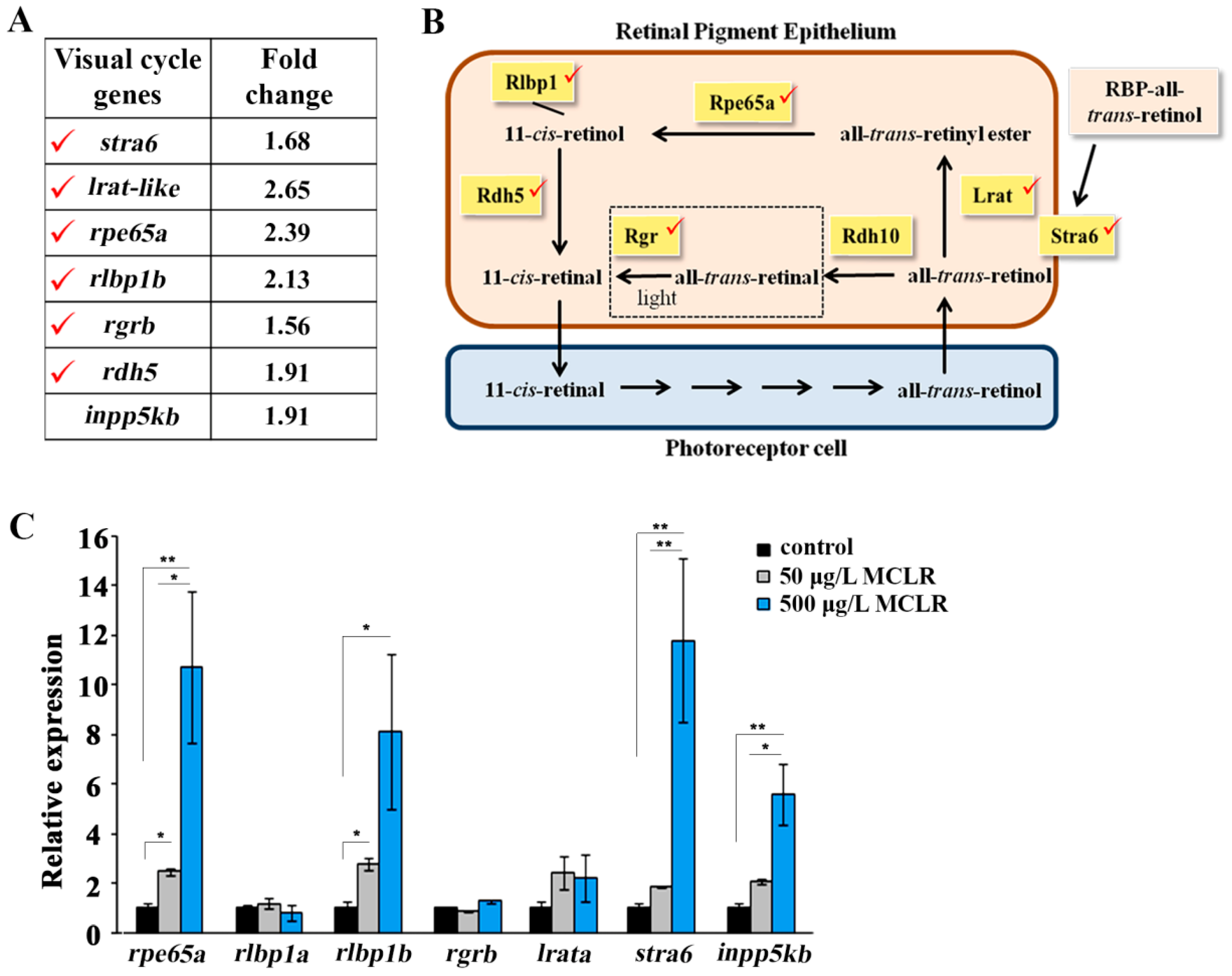
2.2. Gene Expression Analysis
2.3. Behavioral Alterations upon Microcystin-LR Exposure of Zebrafish Larvae
3. Discussion
4. Materials and Methods
4.1. Zebrafish Maintenance
4.2. Exposure of Zebrafish Larvae to Microcystin-LR
4.3. Protein Phosphatase 2A Activity Assay
4.4. RNA Preparation and cDNA Synthesis
4.5. Microarray Analysis
4.6. Quantitative Real-Time PCR
4.7. Behavior Assay
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferrao-Filho Ada, S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Tsuji, K.; Watanabe, M.F. Stability of microcystins from cyanobacteria III. Effect of pH and temperature. Phycologia 1996, 35, 83–88. [Google Scholar] [CrossRef]
- Dziga, D.; Wladyka, B.; Zielinska, G.; Meriluoto, J.; Wasylewski, M. Heterologous expression and characterisation of microcystinase. Toxicon 2012, 59, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E and FN Spon; WHO: London, UK, 1999. [Google Scholar]
- Pavagadhi, S.; Balasubramanian, R. Toxicological evaluation of microcystins in aquatic fish species: Current knowledge and future directions. Aquat. Toxicol. 2013, 142–143, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef]
- Feurstein, D.; Holst, K.; Fischer, A.; Dietrich, D.R. Oatp-associated uptake and toxicity of microcystins in primary murine whole brain cells. Toxicol. Appl. Pharmacol. 2009, 234, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.J.; Chien, M.S.; Wu, F.J.; Chou, H.N.; Lee, S.J. Inhibition of embryonic development by microcystin-LR in zebrafish, Danio rerio. Toxicon 2005, 45, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, T.; Armeni, E.; Stalikas, C.D.; Kagalou, I.; Leonardos, I.D. Detection of microcystins in pamvotis lake water and assessment of cyanobacterial bloom toxicity. Environ. Monit. Assess. 2012, 184, 3043–3052. [Google Scholar] [CrossRef] [PubMed]
- Malbrouck, C.; Kestemont, P. Effects of microcystins on fish. Environ. Toxicol. Chem. 2006, 25, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Wickstrom, M.L.; Haschek, W.M.; Schaeffer, D.J.; Ghosh, S.; Beasley, V.R. Microcystin-LR and kinetics of cytoskeletal reorganization in hepatocytes, kidney cells, and fibroblasts. Nat. Toxins 1996, 4, 206–214. [Google Scholar] [CrossRef]
- Wickstrom, M.L.; Khan, S.A.; Haschek, W.M.; Wyman, J.F.; Eriksson, J.E.; Schaeffer, D.J.; Beasley, V.R. Alterations in microtubules, intermediate filaments, and microfilaments induced by microcystin-LR in cultured cells. Toxicol. Pathol. 1995, 23, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Tu, W.W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon 2015, 101, 92–100. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- Janssens, V.; Goris, J.; van Hoof, C. PP2A: The expected tumor suppressor. Curr. Opin. Genet. Dev. 2005, 15, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Perrotti, D.; Neviani, P. Protein phosphatase 2A (PP2A), a drugable tumor suppressor in Ph1(+) leukemias. Cancer Metastasis Rev. 2008, 27, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Xu, Y.; Chen, Y.; Jeffrey, P.D.; Chao, Y.; Lin, Z.; Li, Z.; Strack, S.; Stock, J.B.; Shi, Y. Structure of protein phosphatase 2A core enzyme bound to tumor-inducing toxins. Cell 2006, 127, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zheng, Q.; Sun, Y.T.; Huang, P.; Guo, Z.L.; Xu, L.H. Microcystin-LR induces protein phosphatase 2a alteration in a human liver cell line. Environ. Toxicol. 2014, 29, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin-LR: The first step of detoxication. Biochim. Biophys. Acta 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Schmidt, J.R.; Wilhelm, S.W.; Boyer, G.L. The fate of microcystins in the environment and challenges for monitoring. Toxins 2014, 6, 3354–3387. [Google Scholar] [CrossRef] [PubMed]
- Kist, L.W.; Rosemberg, D.B.; Pereira, T.C.; de Azevedo, M.B.; Richetti, S.K.; de Castro Leao, J.; Yunes, J.S.; Bonan, C.D.; Bogo, M.R. Microcystin-LR acute exposure increases AChE activity via transcriptional ache activation in zebrafish (Danio rerio) brain. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 247–252. [Google Scholar] [CrossRef]
- Zeng, C.; Sun, H.; Xie, P.; Wang, J.; Zhang, G.; Chen, N.; Yan, W.; Li, G. The role of apoptosis in MCLR-induced developmental toxicity in zebrafish embryos. Aquat. Toxicol. 2014, 149, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kabashi, E.; Brustein, E.; Champagne, N.; Drapeau, P. Zebrafish models for the functional genomics of neurogenetic disorders. Biochim. Biophys. Acta 2011, 1812, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.J.; Jia, Y.F.; Chen, N.; Bian, W.P.; Li, Q.K.; Ma, Y.B.; Chen, Y.L.; Pei, D.S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Stahl, B.J. Early and recent primitive brain forms. Ann. N. Y. Acad. Sci. 1977, 299, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Moser, V.C. Functional assays for neurotoxicity testing. Toxicol. Pathol. 2011, 39, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Fleisch, V.C.; Neuhauss, S.C. Visual behavior in zebrafish. Zebrafish 2006, 3, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Neuhauss, S.C. Behavioral genetic approaches to visual system development and function in zebrafish. J. Neurobiol. 2003, 54, 148–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleisch, V.C.; Neuhauss, S.C. Parallel visual cycles in the zebrafish retina. Prog. Retin. Eye Res. 2010, 29, 476–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhang, X.; Zhou, W.; Qiao, Q.; Liang, H.; Li, G.; Wang, J.; Cai, F. The interactive effects of cytoskeleton disruption and mitochondria dysfunction lead to reproductive toxicity induced by microcystin-LR. PLoS ONE 2013, 8, e53949. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, L.; Liu, W.; Qiao, Q.; Wu, K.; Wen, J.; Huang, C.; Tang, R.; Zhang, X. Involvement of oxidative stress and cytoskeletal disruption in microcystin-induced apoptosis in cik cells. Aquat. Toxicol. 2015, 165, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Redmond, T.M.; Yu, S.; Lee, E.; Bok, D.; Hamasaki, D.; Chen, N.; Goletz, P.; Ma, J.X.; Crouch, R.K.; Pfeifer, K. rpe65 is necessary for production of 11-cis-vitamin a in the retinal visual cycle. Nat. Genet. 1998, 20, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Marlhens, F.; Bareil, C.; Griffoin, J.M.; Zrenner, E.; Amalric, P.; Eliaou, C.; Liu, S.Y.; Harris, E.; Redmond, T.M.; Arnaud, B.; et al. Mutations in rpe65 cause leber’s congenital amaurosis. Nat. Genet. 1997, 17, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Travis, G.H.; Golczak, M.; Moise, A.R.; Palczewski, K. Diseases caused by defects in the visual cycle: Retinoids as potential therapeutic agents. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 469–512. [Google Scholar] [CrossRef] [PubMed]
- Rajala, R.V.; Rajala, A.; Morris, A.J.; Anderson, R.E. Phosphoinositides: Minor lipids make a major impact on photoreceptor cell functions. Sci. Rep. 2014, 4, 5463. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Fan, H.; Xie, P.; He, J. A review of neurotoxicity of microcystins. Environ. Sci. Pollut. Res. Int. 2016, 23, 7211–7219. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yan, W.; Liu, C.; Li, L.; Yu, L.; Zhao, S.; Li, G. Microcystin-LR exposure induces developmental neurotoxicity in zebrafish embryo. Environ. Pollut. 2016, 213, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Emran, F.; Rihel, J.; Dowling, J.E. A behavioral assay to measure responsiveness of zebrafish to changes in light intensities. J. Vis. Exp. 2008, 20, 923. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Patterson, J.; Kimmel, R.O. The development and behavioral characteristics of the startle response in the zebra fish. Dev. Psychobiol. 1974, 7, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Higdon, C.W.; Mitra, R.D.; Johnson, S.L. Gene expression analysis of zebrafish melanocytes, iridophores, and retinal pigmented epithelium reveals indicators of biological function and developmental origin. PLoS ONE 2013, 8, e67801. [Google Scholar] [CrossRef] [PubMed]
- Fleisch, V.C.; Schonthaler, H.B.; von Lintig, J.; Neuhauss, S.C. Subfunctionalization of a retinoid-binding protein provides evidence for two parallel visual cycles in the cone-dominant zebrafish retina. J. Neurosci. 2008, 28, 8208–8216. [Google Scholar] [CrossRef] [PubMed]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed]
- Rogers, E.D.; Henry, T.B.; Twiner, M.J.; Gouffon, J.S.; McPherson, J.T.; Boyer, G.L.; Sayler, G.S.; Wilhelm, S.W. Global gene expression profiling in larval zebrafish exposed to microcystin-LR and microcystis reveals endocrine disrupting effects of cyanobacteria. Environ. Sci. Technol. 2011, 45, 1962–1969. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Leung, H.T.; Pak, W.L.; Carl, Y.T.; Wadzinski, B.E.; Shieh, B.H. Role of protein phosphatase 2A in regulating the visual signaling in drosophila. J. Neurosci. 2008, 28, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Chen, J.; Xie, P.; Jiang, Y.; Wu, L.; Zhang, X. Protein expression profiling in the zebrafish (Danio rerio) embryos exposed to the microcystin-LR. Proteomics 2011, 11, 2003–2018. [Google Scholar] [CrossRef] [PubMed]
- Baganz, D.; Staaks, G.; Pflugmacher, S.; Steinberg, C.E. Comparative study of microcystin-LR-induced behavioral changes of two fish species, Danio rerio and Leucaspius delineatus. Environ. Toxicol. 2004, 19, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Kist, L.W.; Piato, A.L.; da Rosa, J.G.; Koakoski, G.; Barcellos, L.J.; Yunes, J.S.; Bonan, C.D.; Bogo, M.R. Acute exposure to microcystin-producing cyanobacterium microcystis aeruginosa alters adult zebrafish (Danio rerio) swimming performance parameters. J. Toxicol. 2011, 2011, 280304. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Noldus, L.P.J.J.; Tegelenbosch, R.A.J.; Richardson, M.K. Zebrafish embryos and larvae in behavioural assays. Behaviour 2012, 149, 1241–1281. [Google Scholar] [CrossRef]
- Baran, R.; Castelblanco, L.; Tang, G.; Shapiro, I.; Goncharov, A.; Jin, Y. Motor neuron synapse and axon defects in a C. elegans alpha-tubulin mutant. PLoS ONE 2010, 5, e9655. [Google Scholar] [CrossRef] [PubMed]
- Rihel, J.; Prober, D.A.; Arvanites, A.; Lam, K.; Zimmerman, S.; Jang, S.; Haggarty, S.J.; Kokel, D.; Rubin, L.L.; Peterson, R.T.; et al. Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation. Science 2010, 327, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Connon, R.E.; Geist, J.; Pfeiff, J.; Loguinov, A.V.; D’Abronzo, L.S.; Wintz, H.; Vulpe, C.D.; Werner, I. Linking mechanistic and behavioral responses to sublethal esfenvalerate exposure in the endangered delta smelt; Hypomesus transpacificus (Fam. Osmeridae). BMC Genom. 2009, 10, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeti, S.; O’Farrell, S.; Kennedy, B.N. Early safety assessment of human oculotoxic drugs using the zebrafish visualmotor response. J. Pharmacol. Toxicol. Methods 2014, 69, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Transcript ID | Gene Symbol | Gene Description | NCBI Gene ID | p-Value | Fold Change |
|---|---|---|---|---|---|
| Upregulated genes | |||||
| 13122067 | tm6sf2 | transmembrane 6 superfamily member 2 | 791179 | 3.21 × 10−2 | 2.95 |
| 12945361 | zgc:194355 (lrat-like) | zgc:194355 (lecithin retinol acyltransferase-like) | 556575 | 3.15 × 10−2 | 2.65 |
| 13223317 | rdh20 | retinol dehydrogenase 20 | 555864 | 1.42 × 10−2 | 2.42 |
| 13085946 | rpe65a | retinal pigment epithelium-specific protein 65a | 393724 | 7.28 × 10−3 | 2.39 |
| 12985711 | gch2 | GTP cyclohydrolase 2 | 64263 | 9.80 × 10−4 | 2.14 |
| 13263303 | dct | dopachrome tautomerase | 58074 | 1.50 × 10−2 | 2.13 |
| 13156297 | rlbp1b | retinaldehyde binding protein 1b | 402990 | 6.80 × 10−3 | 2.13 |
| 13135621 | pmelb | premelanosome protein b | 562810 | 6.42 × 10−3 | 2.06 |
| 13305424 | slc16a8 | solute carrier family 16 (monocarboxylate transporter), member 8 | 557884 | 3.47 × 10−2 | 1.96 |
| 13112995 | LOC103911087 | uncharacterized LOC103911087 | 103911087 | 7.87 × 10−3 | 1.95 |
| 12968503 | inpp5kb | inositol polyphosphate-5-phosphatase Kb | 566188 | 2.05 × 10−3 | 1.91 |
| 13126385 | rdh5 | retinol dehydrogenase 5 (11-cis/9-cis) | 556528 | 3.12 × 10−2 | 1.91 |
| 13279630 | stra6 | stimulated by retinoic acid 6 | 724007 | 2.31 × 10−2 | 1.73 |
| 12972154 | pmela | premelanosome protein a | 321239 | 1.63 × 10−3 | 1.72 |
| 12946322 | tyrp1b | tyrosinase-related protein 1b | 437022 | 6.72 × 10−5 | 1.70 |
| 13284986/ 13022340 | tyr | tyrosinase | 30207 | 9.79 × 10−3 5.54 × 10−3 | 1.69 1.55 |
| 13156897 | stra6 | stimulated by retinoic acid 6 | 724007 | 7.76 × 10−3 | 1.68 |
| 12988859 | si:ch1073-13h15.3 (LOC563241) | si:ch1073-13h15.3 (putative all-trans-retinol 13,14-reductase) | 563241 | 2.07 × 10−2 | 1.68 |
| 13109427 | slc45a2 | solute carrier family 45, member 2 | 558311 | 3.11 × 10−2 | 1.66 |
| 13129757 | LOC101882639 | uncharacterized LOC101882639 | 101882639 | 3.57 × 10−2 | 1.66 |
| 13133349 | LOC566587 | Erb-b2 receptor tyrosine kinases (ERBB) receptor feedback inhibitor1-like | 566587 | 1.79 × 10−2 | 1.60 |
| 13182209 | si:dkey-31f5.1 (slc26a3.2) | solute carrier family 26 (anion exchanger), member 3, tandem duplicate 2 | 563896 | 9.96 × 10−3 | 1.59 |
| 13274399 | bco1l | β-carotene oxygenase 1, like | 393580 | 2.98 × 10−2 | 1.57 |
| 12994027 | rgrb | retinal G protein coupled receptor b | 554142 | 3.02 × 10−2 | 1.56 |
| 13054709 | LOC100535423 | urokinase plasminogen activator surface receptor-like | 100535423 | 1.69 × 10−2 | 1.56 |
| 13216589 | mct1b (slc16a1a) | solute carrier family 16 (monocarboxylate transporter), member 1a | 100534752 | 4.29 × 10−2 | 1.55 |
| 13233501 | tyrp1a | tyrosinase-related protein 1a | 100333145 | 7.52 × 10−4 | 1.51 |
| 13071881 | slc39a4 | solute carrier family 39 (zinc transporter), member 4 | 562762 | 4.36 × 10−2 | 1.51 |
| 12958158 | zgc:154142 | zgc:154142 | 555481 | 1.50 × 10−2 | 1.50 |
| 13215240 | oca2 | oculocutaneous albinism II | 567419 | 5.33 × 10−3 | 1.50 |
| Downregulated genes | |||||
| 13089117 | sepp1b | selenoprotein P, plasma, 1b | 791479 | 5.75 × 10−3 | 0.66 |
| 13124494/ 13279064 | cp | ceruloplasmin | 84702 | 7.16 × 10−3 2.04 × 10−2 | 0.66 0.66 |
| 12993720 | LOC568930 | uncharacterized LOC568930 | 568930 | 3.97 × 10−2 | 0.64 |
| 13185986 | LOC100331497 | U2 small nuclear ribonucleoprotein auxiliary factor 35 kDa subunit-related protein 1-like | 100331497 | 1.11 × 10−3 4.63 × 10−2 | 0.64 0.56 |
| 13269248/ 13285056 | ttn.2 (ttna) | titin, tandem duplicate 2 (titin a) | 317731 | 1.54 × 10−2 4.63 × 10−2 | 0.63 0.56 |
| 13269083 | ttnb | titin b | 100001684 | 1.40 × 10−2 | 0.63 |
| 13070223 | si:ch211-250g4.3 | si:ch211-250g4.3 | 557772 | 3.90 × 10−2 | 0.63 |
| 12959767 | LOC100330916 | uncharacterized LOC100330916 | 100330916 | 3.61 × 10−2 | 0.60 |
| 13283642 | si:dkey-7c18.24 | si:dkey-7c18.24 | 562950 | 3.36 × 10−3 | 0.60 |
| 13037425 | si:dkey-8k3.2 | si:dkey-8k3.2 | 794635 | 3.54 × 10−2 | 0.60 |
| 13191393 | LOC100004951 | stonus toxin subunit β-like (neoverrucotoxin subunit beta-like) | 100004951 | 2.96 × 10−2 | 0.60 |
| 13002727 | si:ch211-270n8.1 | si:ch211-270n8.1 | 792467 | 3.27 × 10−2 | 0.59 |
| 13283794 | zgc:172075 | zgc:172075 | 555875 | 4.83 × 10−2 | 0.58 |
| 13104731 | si:dkey-1j5.4 | si:dkey-1j5.4 | 563949 | 3.26 × 10−3 | 0.54 |
| 13072283 | si:ch211-133n4.9 | si:ch211-133n4.9 | 100000061 | 2.22 × 10−2 | 0.53 |
| 12993635 | zmp:0000001031 (LOC568241) | zmp:0000001031 (uncharacterized LOC568241) | 568241 | 3.33 × 10−2 | 0.44 |
| 13282574/ 13284122 | lgals1l1 | lectin, galactoside-binding, soluble, 1 (galectin 1)-like 1 | 326706 | 4.84 × 10−3 4.84 × 10−3 | 0.33 0.33 |
| 13076755 | grn2 | granulin 2 | 336575 | 4.55 × 10−2 | 0.32 |
| 13121248 | krt96 | keratin 96 | 321502 | 3.79 × 10−2 | 0.26 |
| Gene Symbol | Gene ID | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|---|
| stra6 | 724007 | CAAGCAATTGTTGTGTTTTGTGTC | TGGTGGGATAACTTCGACAGG |
| lrata | 553239 | TACGCGTCGATTCGGTTGAA | TTACAAAACTTGTCGGTCTGCTC |
| rpe65a | 393724 | AGAGACGGGACGGTCTACAA | CCGTCATCCCAAAACTGTGC |
| rlbp1a | 393678 | TTGAACATCTGACTGTGAAAGACC | GCCTGCCTTGTCTTTAATCATGG |
| rlbp1b | 402990 | TGAGCTTGCTAAAGGTGTTCAGG | TCAGGATAATCCCGTCTGAAGC |
| rgrb | 554142 | GGAGCTTTAAAACGCGCACA | CTCTCGAACCCTGAGGAACG |
| inpp5k | 566188 | GGTTTGTATAAGCCATAGCAAGATG | GTGCAATCTGAAGGACTCTCTG |
| tyr | 30207 | GCGCTGGAAGGTTTTGCTAAT | AAATGGGGTCGTTGGCAGAT |
| dct | 58074 | TGTCTAAAGAGTGCTGCCCG | CCGGCAAAGTTTCCAAAGCA |
| pmela | 321239 | CTCCTGCTCCAGTTACAGATGA | CGTTGGCTACAACTCCCTCC |
| pmelb | 562810 | CACACAGTTTCACGAAGGCG | GCCAGTATTTGCCCCAGGTT |
| cp | 84702 | GAAAGAAAGCCCAGGCAACG | ATATCGGCGGTCCTCTCCTT |
| sepp1b | 791479 | TCTACAGTGGTTGAAGTCCAGC | TCCTCGAACCACTGCTTTCC |
| tuba1b | 373080 | AATCACCAATGCTTGCTTCGAGCC | TTCACGTCTTTGGGTACCACGTCA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzima, E.; Serifi, I.; Tsikari, I.; Alzualde, A.; Leonardos, I.; Papamarcaki, T. Transcriptional and Behavioral Responses of Zebrafish Larvae to Microcystin-LR Exposure. Int. J. Mol. Sci. 2017, 18, 365. https://doi.org/10.3390/ijms18020365
Tzima E, Serifi I, Tsikari I, Alzualde A, Leonardos I, Papamarcaki T. Transcriptional and Behavioral Responses of Zebrafish Larvae to Microcystin-LR Exposure. International Journal of Molecular Sciences. 2017; 18(2):365. https://doi.org/10.3390/ijms18020365
Chicago/Turabian StyleTzima, Eleni, Iliana Serifi, Ioanna Tsikari, Ainhoa Alzualde, Ioannis Leonardos, and Thomais Papamarcaki. 2017. "Transcriptional and Behavioral Responses of Zebrafish Larvae to Microcystin-LR Exposure" International Journal of Molecular Sciences 18, no. 2: 365. https://doi.org/10.3390/ijms18020365
APA StyleTzima, E., Serifi, I., Tsikari, I., Alzualde, A., Leonardos, I., & Papamarcaki, T. (2017). Transcriptional and Behavioral Responses of Zebrafish Larvae to Microcystin-LR Exposure. International Journal of Molecular Sciences, 18(2), 365. https://doi.org/10.3390/ijms18020365