
Clinical and Molecular Evidence of ABCC11 Protein Expression in Axillary Apocrine Glands of Patients with Axillary Osmidrosis

 , ,
, ,  ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Generation and Validation of Anti-ABCC11 Antibody
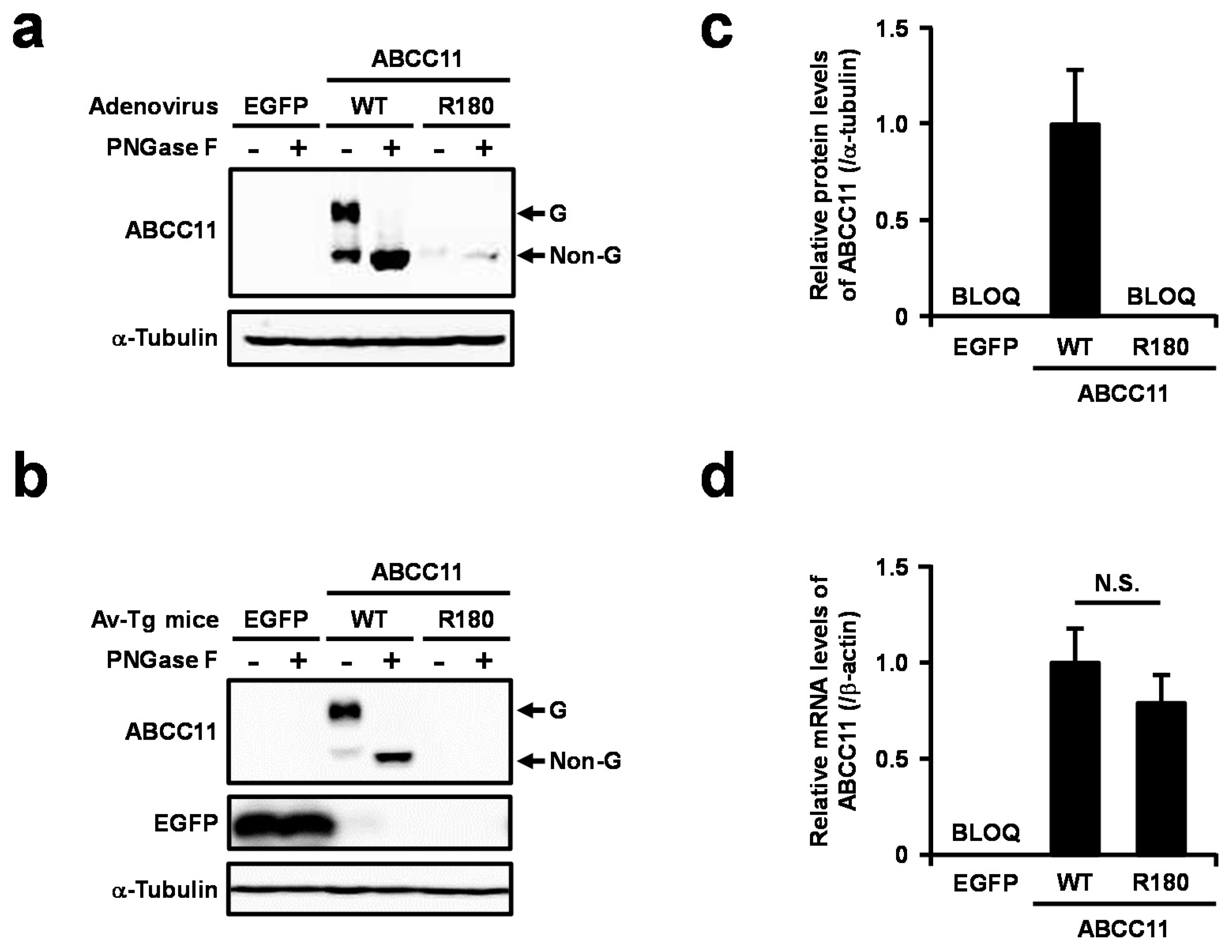
2.2. Generation and Analysis of Transiently ABCC11-Expressing Transgenic Mice
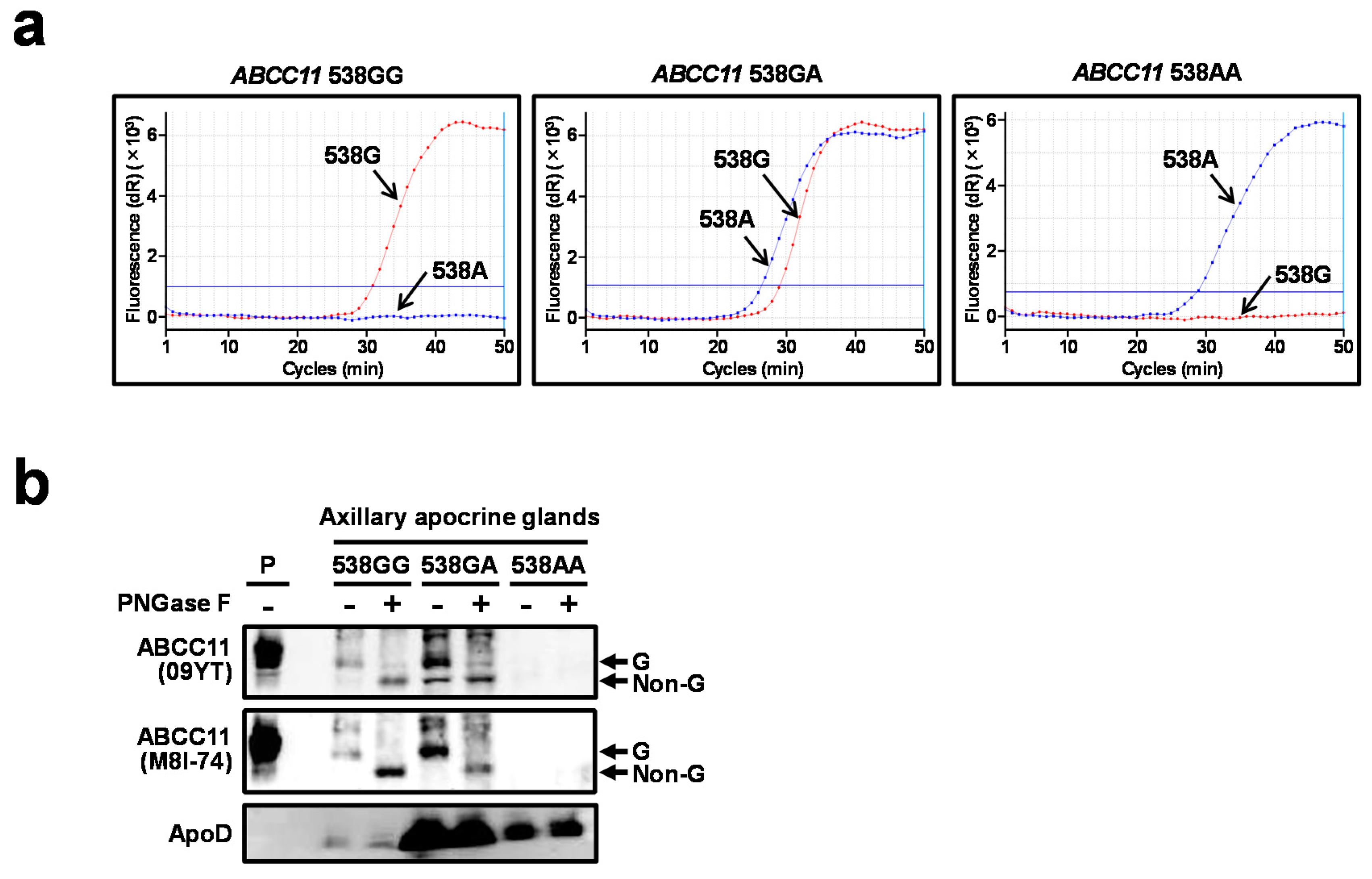
2.3. Expression of ABCC11 WT Protein in Axillary Apocrine Glands Derived from AO Patient
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Construction of and Infection with Recombinant Adenovirus
4.4. Preparation of Plasma Membrane Vesicles and the In Vitro Transport Experiment
4.5. Animals
4.6. Immunoblotting Analysis
4.7. Quantification of mRNA Expression
4.8. Patients and Sample Collection
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| AO | Axillary osmidrosis |
| ERAD | Endoplasmic reticulum-associated protein degradation |
| E1S | Estrone sulfate |
| MRP8 | Multidrug resistance-associated protein 8 |
| QPCR | Quantitative polymerase chain reaction |
| SNP | Single nucleotide polymorphism |
| WT | Wild-type |
References
- Toyoda, Y.; Gomi, T.; Nakagawa, H.; Nagakura, M.; Ishikawa, T. Diagnosis of Human Axillary Osmidrosis by Genotyping of the Human ABCC11 Gene: Clinical Practice and Basic Scientific Evidence. BioMed Res. Int. 2016, 2016, 7670483. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Steer, C.D.; Farrow, A.; Golding, J.; Day, I.N. Dependence of deodorant usage on ABCC11 genotype: Scope for personalized genetics in personal hygiene. J. Investig. Dermatol. 2013, 133, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Leyden, J.J.; McGinley, K.J.; Holzle, E.; Labows, J.N.; Kligman, A.M. The microbiology of the human axilla and its relationship to axillary odor. J. Investig. Dermatol. 1981, 77, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Morioka, D.; Ohkubo, F.; Amikura, Y. Clinical features of axillary osmidrosis: A retrospective chart review of 723 Japanese patients. J. Dermatol. 2013, 40, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Bang, Y.H.; Kim, J.H.; Paik, S.W.; Park, S.H.; Jackson, I.T.; Lebeda, R. Histopathology of apocrine bromhidrosis. Plast. Reconstr. Surg. 1996, 98, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Saathoff, M.; Kuhn, F.; Max, H.; Terstegen, L.; Natsch, A. A functional ABCC11 allele is essential in the biochemical formation of human axillary odor. J. Investig. Dermatol. 2010, 130, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Mori, T.; Toyoda, Y.; Sakurai, A.; Ishikawa, T.; Mitani, Y.; Hayashizaki, Y.; Yoshimura, Y.; Kurahashi, H.; Sakai, Y. Correlation of axillary osmidrosis to a SNP in the ABCC11 gene determined by the Smart Amplification Process (SmartAmp) method. J. Plast. Reconstr. Aesthet. Surg. 2010, 63, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Sakurai, A.; Mitani, Y.; Nakashima, M.; Yoshiura, K.; Nakagawa, H.; Sakai, Y.; Ota, I.; Lezhava, A.; Hayashizaki, Y.; et al. Earwax, osmidrosis, and breast cancer: Why does one SNP (538G>A) in the human ABC transporter ABCC11 gene determine earwax type? FASEB J. 2009, 23, 2001–2013. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Miwa, N.; Hirano, A.; Yoshiura, K.; Niikawa, N. A strong association of axillary osmidrosis with the wet earwax type determined by genotyping of the ABCC11 gene. BMC Genet. 2009, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Yoshiura, K.; Kinoshita, A.; Ishida, T.; Ninokata, A.; Ishikawa, T.; Kaname, T.; Bannai, M.; Tokunaga, K.; Sonoda, S.; Komaki, R.; et al. A SNP in the ABCC11 gene is the determinant of human earwax type. Nat. Genet. 2006, 38, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Takada, T.; Miyata, H.; Ishikawa, T.; Suzuki, H. Regulation of the Axillary Osmidrosis-Associated ABCC11 Protein Stability by N-Linked Glycosylation: Effect of Glucose Condition. PLoS ONE 2016, 11, e0157172. [Google Scholar] [CrossRef] [PubMed]
- Aw, W.; Ota, I.; Toyoda, Y.; Lezhava, A.; Sakai, Y.; Gomi, T.; Hayashizaki, Y.; Ishikawa, T. Pharmacogenomics of human ABC transporters: Detection of clinically important SNPs by SmartAmp2 method. Curr. Pharm. Biotechnol. 2011, 12, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Spielman, A.I.; Vowels, B.R.; Leyden, J.J.; Biemann, K.; Preti, G. A human axillary odorant is carried by apolipoprotein D. Proc. Natl. Acad. Sci. USA 1996, 93, 6626–6630. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Ito, K.; Ikebuchi, Y.; Toyoda, Y.; Takada, T.; Hisaka, A.; Oka, A.; Suzuki, H. Prediction of Drug Transfer into Milk Considering Breast Cancer Resistance Protein (BCRP)-Mediated Transport. Pharm. Res. 2015, 32, 2527–2537. [Google Scholar] [CrossRef] [PubMed]
- Yoshikado, T.; Takada, T.; Yamamoto, T.; Yamaji, H.; Ito, K.; Santa, T.; Yokota, H.; Yatomi, Y.; Yoshida, H.; Goto, J.; et al. Itraconazole-induced cholestasis: Involvement of the inhibition of bile canalicular phospholipid translocator MDR3/ABCB4. Mol. Pharmacol. 2011, 79, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Hagiya, Y.; Adachi, T.; Hoshijima, K.; Kuo, M.T.; Ishikawa, T. MRP class of human ATP binding cassette (ABC) transporters: Historical background and new research directions. Xenobiotica 2008, 38, 833–862. [Google Scholar] [CrossRef] [PubMed]
- Kruh, G.D.; Guo, Y.; Hopper-Borge, E.; Belinsky, M.G.; Chen, Z.S. ABCC10, ABCC11, and ABCC12. Pflugers Arch. 2007, 453, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Takada, T.; Suzuki, H. Halogenated hydrocarbon solvent-related cholangiocarcinoma risk: Biliary excretion of glutathione conjugates of 1,2-dichloropropane evidenced by untargeted metabolomics analysis. Sci. Rep. 2016, 6, 24586. [Google Scholar] [CrossRef] [PubMed]
- Miyata, H.; Takada, T.; Toyoda, Y.; Matsuo, H.; Ichida, K.; Suzuki, H. Identification of febuxostat as a new strong ABCG2 inhibitor: Potential applications and risks in clinical situations. Front. Pharmacol. 2016, 7, 518. [Google Scholar] [CrossRef] [PubMed]
- Takada, T.; Yamanashi, Y.; Konishi, K.; Yamamoto, T.; Toyoda, Y.; Masuo, Y.; Yamamoto, H.; Suzuki, H. NPC1L1 is a key regulator of intestinal vitamin K absorption and a modulator of warfarin therapy. Sci. Transl. Med. 2015, 7, 275ra223. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Wakabayashi-Nakao, K.; Tamura, A.; Toyoda, Y.; Koshiba, S.; Ishikawa, T. Disruption of N-linked glycosylation enhances ubiquitin-mediated proteasomal degradation of the human ATP-binding cassette transporter ABCG2. FEBS J. 2009, 276, 7237–7252. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyoda, Y.; Takada, T.; Gomi, T.; Nakagawa, H.; Ishikawa, T.; Suzuki, H. Clinical and Molecular Evidence of ABCC11 Protein Expression in Axillary Apocrine Glands of Patients with Axillary Osmidrosis. Int. J. Mol. Sci. 2017, 18, 417. https://doi.org/10.3390/ijms18020417
Toyoda Y, Takada T, Gomi T, Nakagawa H, Ishikawa T, Suzuki H. Clinical and Molecular Evidence of ABCC11 Protein Expression in Axillary Apocrine Glands of Patients with Axillary Osmidrosis. International Journal of Molecular Sciences. 2017; 18(2):417. https://doi.org/10.3390/ijms18020417
Chicago/Turabian StyleToyoda, Yu, Tappei Takada, Tsuneaki Gomi, Hiroshi Nakagawa, Toshihisa Ishikawa, and Hiroshi Suzuki. 2017. "Clinical and Molecular Evidence of ABCC11 Protein Expression in Axillary Apocrine Glands of Patients with Axillary Osmidrosis" International Journal of Molecular Sciences 18, no. 2: 417. https://doi.org/10.3390/ijms18020417
APA StyleToyoda, Y., Takada, T., Gomi, T., Nakagawa, H., Ishikawa, T., & Suzuki, H. (2017). Clinical and Molecular Evidence of ABCC11 Protein Expression in Axillary Apocrine Glands of Patients with Axillary Osmidrosis. International Journal of Molecular Sciences, 18(2), 417. https://doi.org/10.3390/ijms18020417