1. Introduction
Cervical cancer is the fourth most frequent cancer in women worldwide (about 530,000 new cases in 2012, 7.5% of all female cancer deaths). The leading cause of cervical cancer is a persistent infection with high-risk human papillomavirus (HR-HPV) [
1]. Specifically, the HPV subtypes 16 and 18 cause about 70% of all cancer cases [
1,
2]. A total of 170 HPV-types have been described currently [
3]. The infection with 15 types of HPV most likely leads to cancer, which is why these 15 types are called carcinogenic or high-risk types [
4]. The genome of human papillomaviruses consists of approximately 8000 base pairs and contains six “early genes” (E6, E7, E1, E2, E4, E5) and two “late genes” (L1, L2) [
5]. In case of replication of the viral gene E6, the E6 oncoprotein is expressed, which disturbs the cell cycle [
6]. E6 oncoprotein and E6-associated protein (E6-AP) form a complex which binds to p53 and causes its proteolytic degradation [
7].
During the different stages of cervical cancer development, there is an accumulation of epigenetic alterations that leads to changes in gene expression [
8]. Altered mechanisms of epigenetic regulation in cervical cancer include DNA methylation and post-translational modifications of histone proteins [
8]. It has been reported that histone modifying enzymes such as histone deacetylase (HDAC)-1 and HDAC2 are over-expressed in cervical dysplasia and invasive carcinoma [
9]. These results suggest that the dysregulation of enzymes that modify histones in cervical cancer are of importance for the biology of this tumor entity.
HR-HPVs establish persistent infection by maintaining their genomes as extrachromosomal elements—the so-called episome—that replicate, together with host DNA, in infected cells [
10]. By associating with the host chromatin, HR-HPV redirects the normal cellular control of chromatin to create a cellular environment that is beneficial for both the HR-HPV multiplication and malignant progression of the infected cell. Therefore, the investigation of HPV–host chromatin interaction will offer new insights into the importance of HPV-driven chromatin regulation in cervical cancer tissue [
10].
The state of histone modifications that are connected to the early and late HPV viral promoters—modification by acetylation and methylation—were examined in a previous study in cell culture systems using chromatin immunoprecipitation assays: in undifferentiated cells, di-methylated forms of histone H3K4 as well as acetylated histone H3 and H4 were found [
11]. Together with differentiation, the levels of di-methylated H3K4 and acetylated H3 are increased, while the acetylated H4 is also increased, which suggests that nucleosomes are activated through histone modifications to coordinate the HPV transcription during cell differentiation [
11].
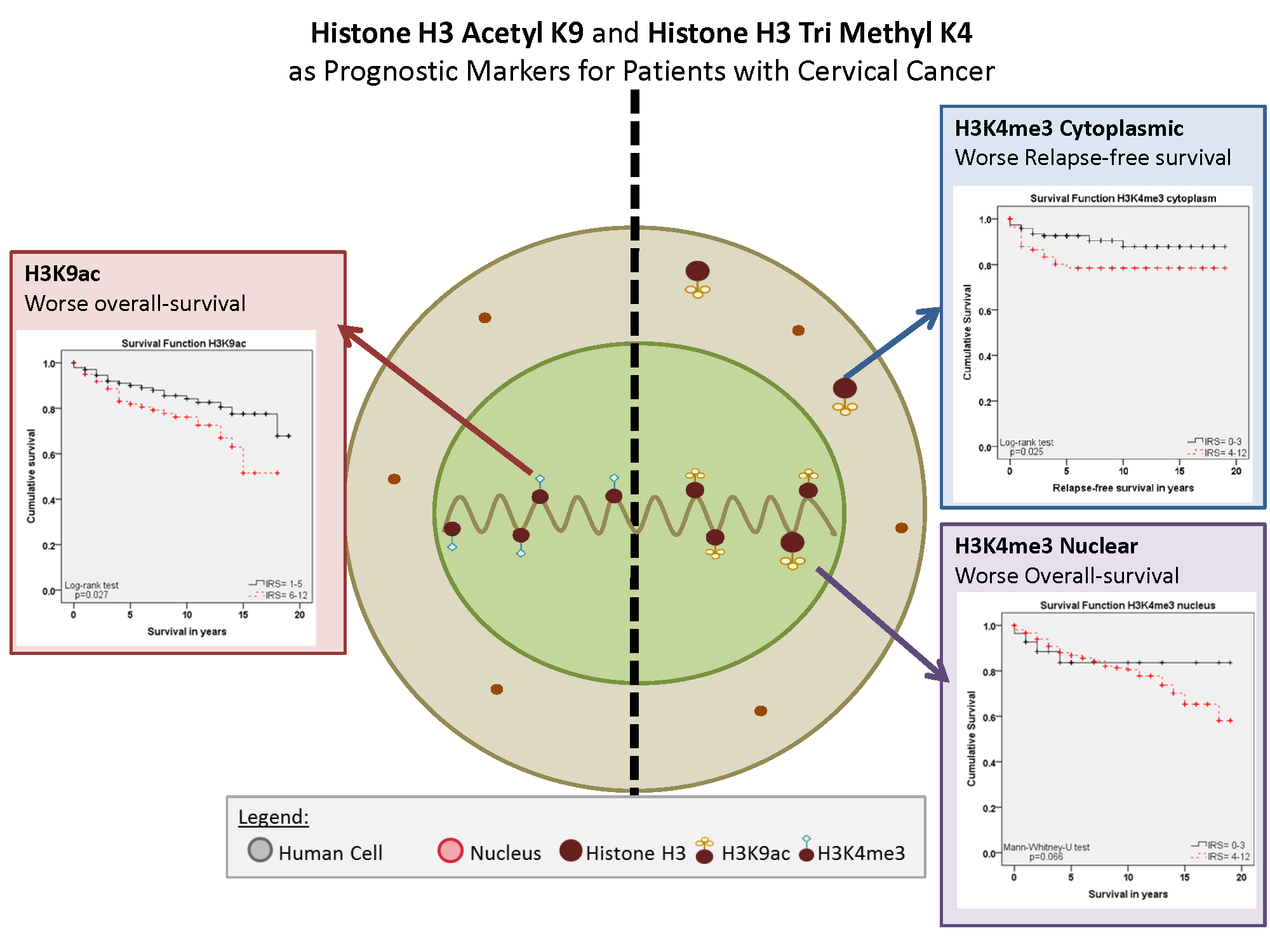
The already-mentioned studies and several other studies showed that histone protein modifications play a fundamental role in HPV driven oncogenesis. Because a systematic investigation of posttranslational changes in histone proteins, for their prognostic relevance in cervical cancer tissue, was lacking, the aim of this study was an expression analyses of histone H3 acetyl K9 (H3K9ac) and histone H3 tri methyl K4 (H3K4me3) in cervical cancer, examined in 250 cases by immunohistochemical methods and assessed by a semi-quantitative score.
3. Discussion
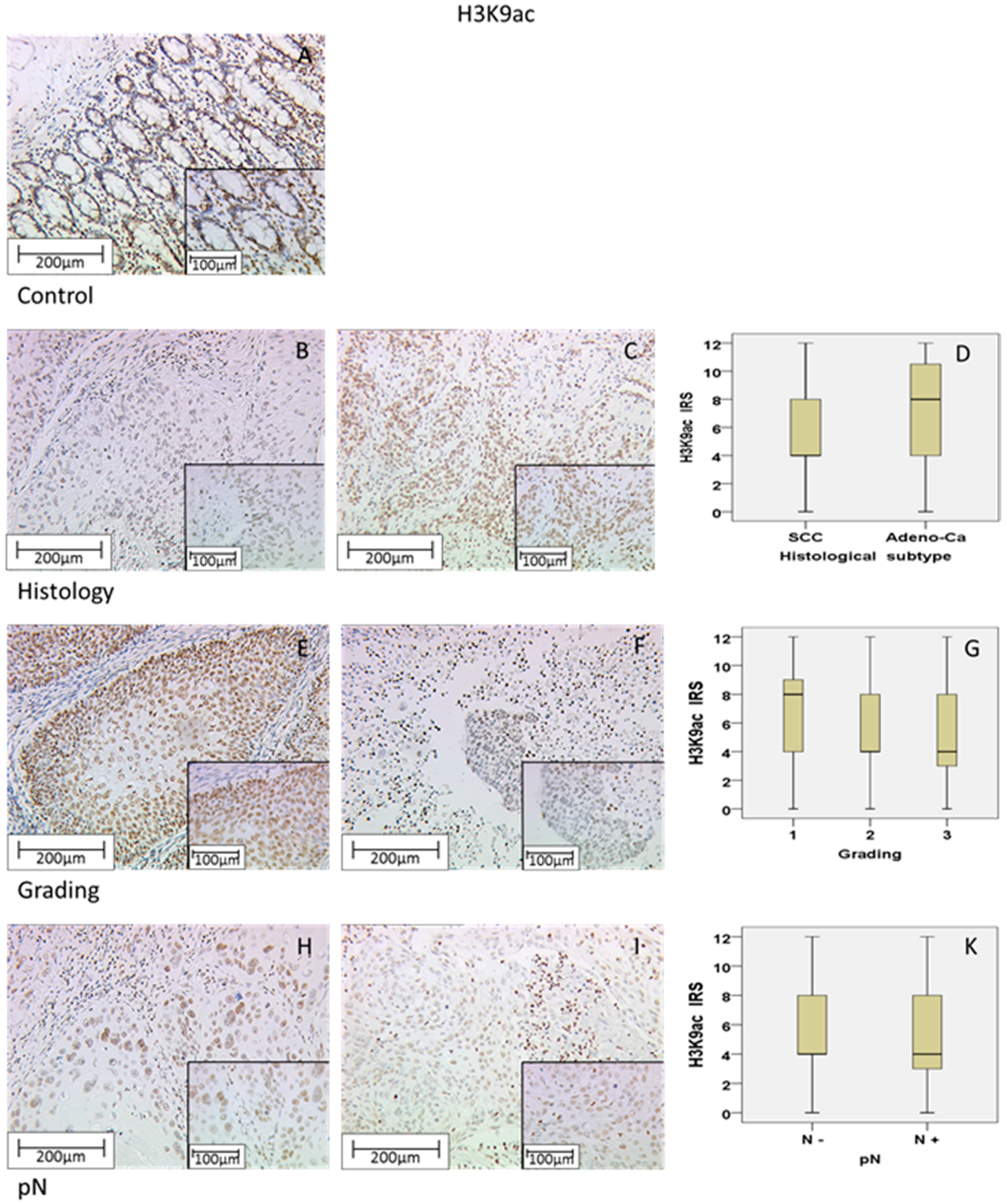
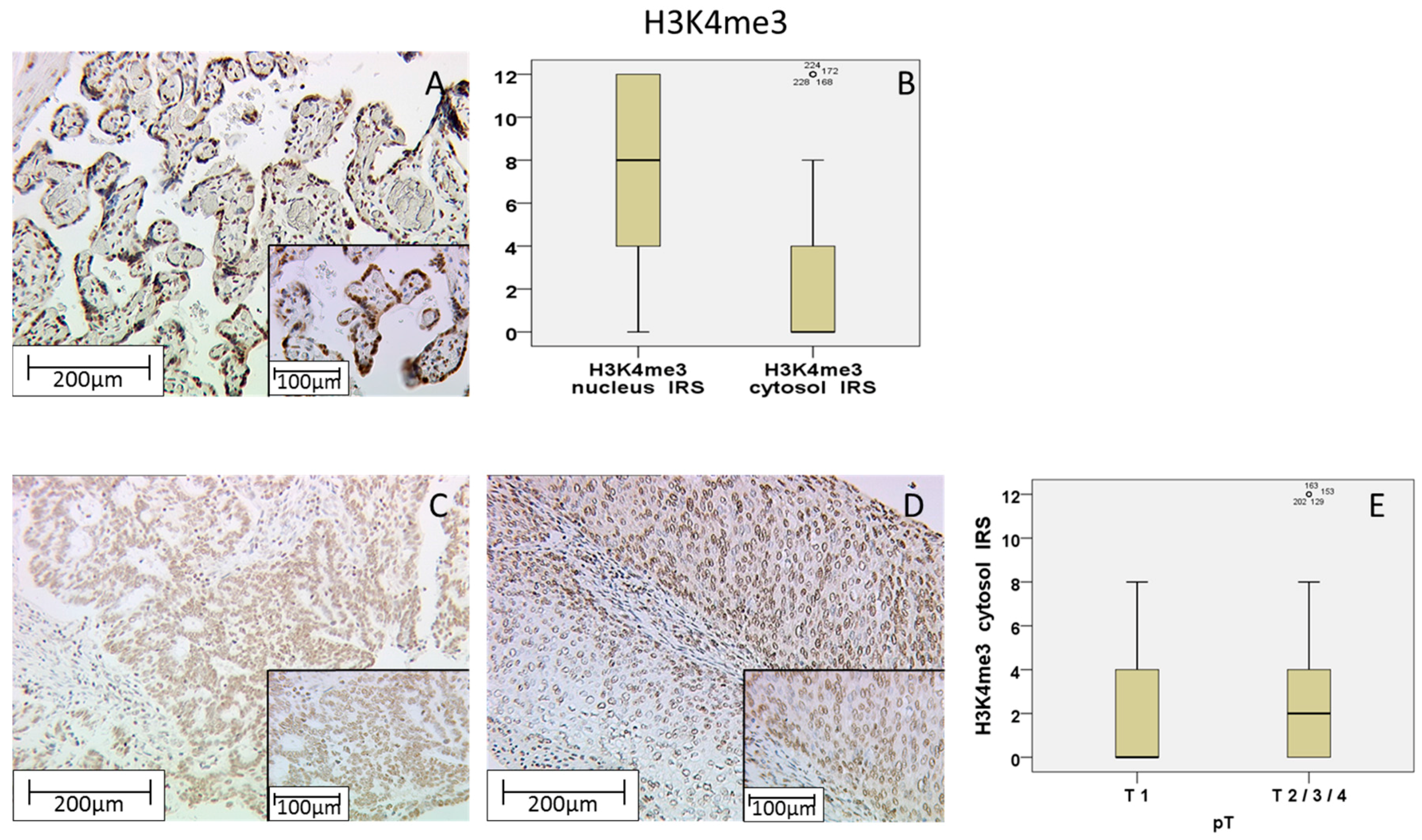
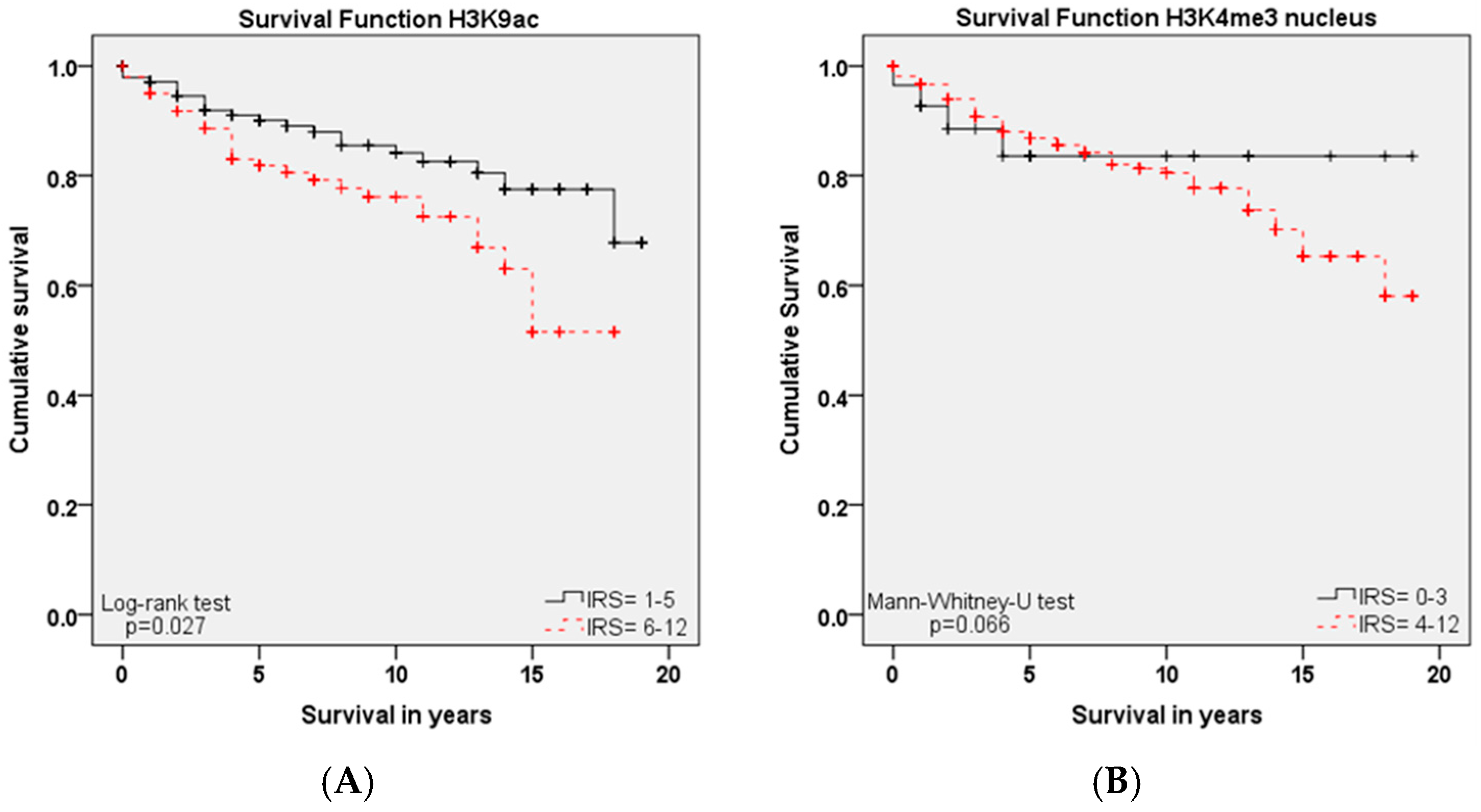
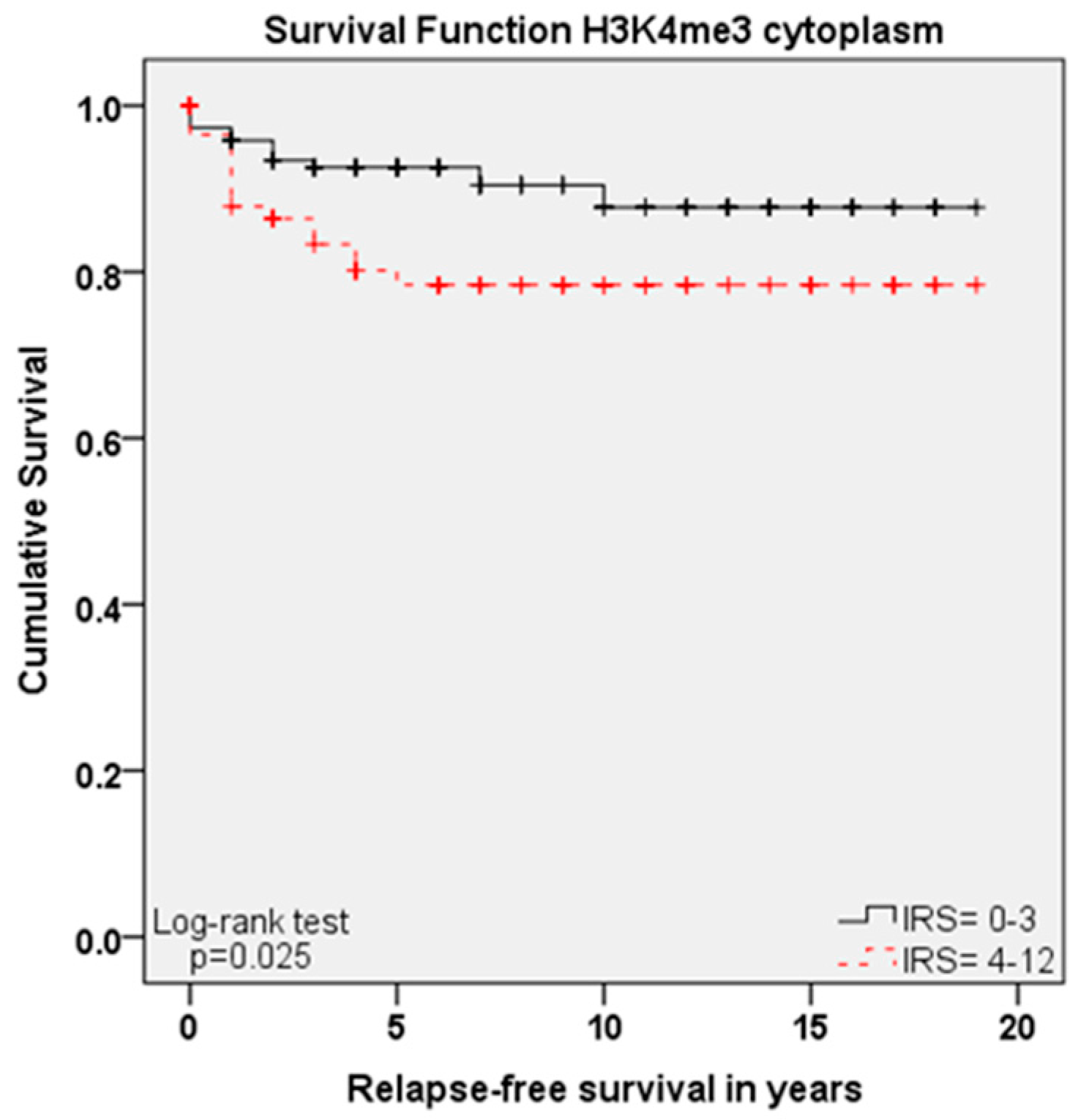
Within this study, we showed that the immunohistochemical evaluation of histone H3K9ac staining was correlated with low grading, low FIGO-classification, low T-status and negative N-status in cervical cancer. We could also find a higher expression of histone H3K9ac in adenocarcinoma compared to squamous cell carcinoma. Due to its correlation between expression and poor prognosis (overall survival), it could be used as an independent marker of prognosis. Cytoplasmic expression of histone H3K4me3 in a cervical cancer specimen was correlated with advanced T-status and poor prognosis. It seems to be a marker of relapse-free survival, while nuclear expression showed a correlation to poor prognosis without being an independent marker regarding cervical cancer.
Histone proteins give the genome the ability to pack very large amounts of DNA in a very small space, but at the same time they leave their N-terminal tails flexible [
15]. The N-terminal tail of the histone proteins can undergo post-translational modification by enzymes, adding chemical modifications such as acetylation, methylation, phosphorylation and deamination that alter the structure of the DNA package and allow or prevent gene transcription [
16]. It is already known that histone modifications at histone 3 lysine 9 acetylation (H3K9ac) denote active regions at enhancers as well as promoters, whereas the tri-methyl form, H3K4me3, preferentially identifies the gene promoters that are active [
17,
18]. In addition, it has been shown that epigenetic modulations of the genome involve histone modifications that alter the gene chromatin configuration. A decondensed (“open”) configuration allows transcription factors access to binding sites, whereas a condensed (“closed”) configuration blocks transcription binding sites, thereby regulating gene transcription [
19,
20]. Based on these findings, it has been shown that high metastatic potential had greater acetylation of histone H3 lysine 9 (H3K9ac) and tri-methylation of histone H3 lysine 4 (H3K4me3) [
19]. Therefore, these two modifications that are correlated to enhanced gene activity and in addition show high metastatic potential were used in the present study as markers for the identification of the prognostic relevance of those histone modifications for cervical cancer survival.
It is already known that E6 oncoprotein and E6-associated protein (E6-AP) form a complex which binds to p53 and causes its proteolytic degradation [
7]. P53 is a tumor suppressor, as it leads to cell cycle arrest or apoptosis in the case of DNA damage [
21]. As E6 oncoprotein induces the degradation of p53, the function of this important cell cycle protein is disturbed [
12] after HPV infection. The cell cycle regulation protein p16 is expressed at high levels in HPV-infected epithelial cells, which is why it acts as a marker for the diagnosis of a HPV associated carcinoma [
22,
23]. On the other hand, studies have shown that p16 expression is induced by an oncogene senescence-related mechanism that involves histone H3K27 demethylation by histone lysine demethylase, and that p16 expression is necessary for the survival of HPV-infected cells expressing E7 viral oncoprotein [
24,
25].
Unfortunately, p16 is not exclusively increased by E7 oncoprotein in carcinogenesis. Therefore, in a recent study, we established and published an immunohistochemical approach for the direct detection of E6 oncoprotein in uterine cervical cancer [
14]. In addition, we found a very high mutation rate of TP53 in this cancer type where p53 is initially inactivated via E6 during the development of cervical cancer. An unexpected finding is the correlation of this mutation with better survival, possibly due to better response to therapy [
13].
Because both H3K9ac and H3K4me3 are negative prognosticators for cervical cancer patients, the use of epigenetic drugs or the search for epigenetic targets could be a useful goal for cervical cancer treatment.
Recently, two main classes of epigenetic drugs—methylation inhibitors and HDAC inhibitors—are in clinical trials for the treatment of cervical cancer [
26]. One of these potential new drugs could be valproic acid (VPA). VPA was found to be an effective inhibitor of histone deacetylases and has been shown to induce anti-tumor effects by modulating cellular pathways, including cell cycle arrest, apoptosis, angiogenesis, metastasis, differentiation, and senescence [
27]. The antitumor effect of VPA in cervical cancer can be explained by either the hyper-acetylation of p53 protein, protecting it from degradation by E6 and increasing p53 activity; or via the inhibition of Akt1 and Akt2 expression, which results in apoptotic cell death [
28,
29]. Acetylation of p53 is a process that occurs in response to DNA damage and stress and is necessary for p53 transcriptional activity. Therefore, p53 was one of the first non-histone proteins that could be acetylated by histone acetyl transferases [
30].
In addition, HDAC inhibitors also interfere with cervical cancer via non-histone targets. The HDAC inhibitor suberoylanilide hydroxamic acid (SAHA) induces apoptosis in HeLa cervical cancer cells in vitro with bortezomib by activating caspase-3 and increasing the ratio of bax/bcl-2 expression [
31]. Epigenetic aberrations, such as histone protein modification have the ability to regulate the expression of oncogenes or repression of tumor suppressor genes. Therefore, these modified histone proteins are powerful candidates for the investigation of cancer pathogenesis and progression. For cervical cancer, for instance, Feng et al. [
26] highlighted a number of genes that underwent epigenetic alteration at the level of DNA methylation, histone modification, or noncoding RNA action in this type of cancer.
Further investigation of these alterations and information about them could lead to new and reliable screening methods for women at high risk of cervical cancer and can help to establish new candidates for a better treatment of this disease.



 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}