
Nimodipine but Not Nifedipine Promotes Expression of Fatty Acid 2-Hydroxylase in a Surgical Stress Model Based on Neuro2a Cells

Abstract
:1. Introduction
2. Results
2.1. Survival of Nimodipine-Treated Differentiated Neuro2a Cells
2.2. Survival of Nifedipine-Treated Neuro2a Cells
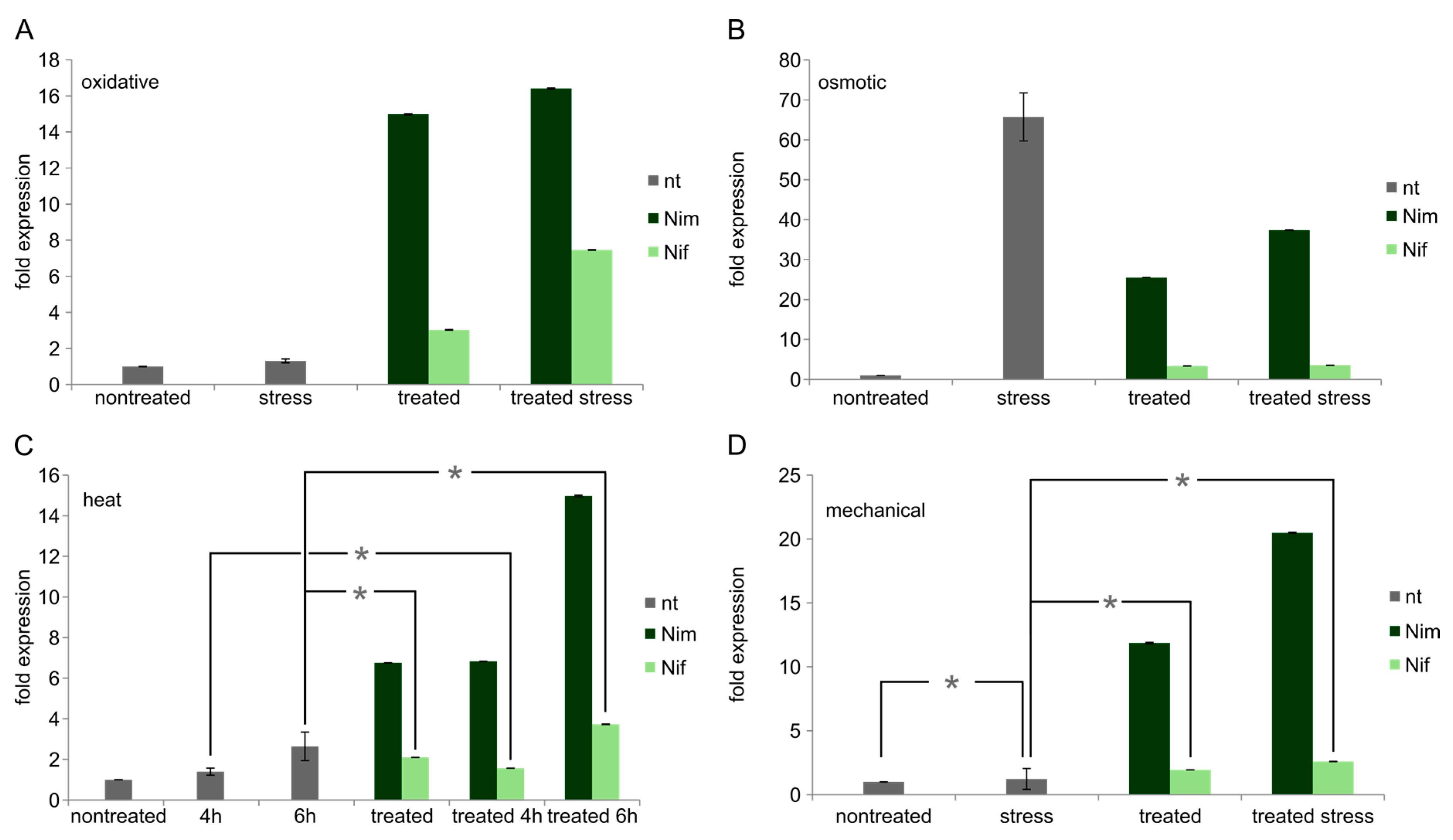
2.3. Expression Levels of FA2H mRNA in Undifferentiated Neuro2a Cells
2.4. Expression Levels of FA2H in Differentiated Neuro2a Cells
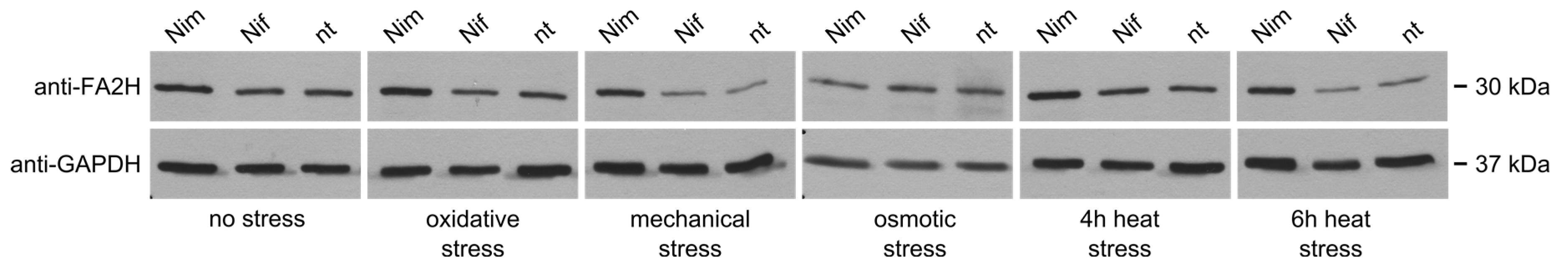
2.5. Protein Levels of FA2H in Undifferentiated Neuro2a Cells
2.6. Protein Levels of FA2H in Differentiated Neuro2a Cells
3. Discussion
4. Materials and Methods
4.1. Value of Experiments
4.2. Cell Culture
4.3. Nimodipine/Nifedipine Treatment
4.4. Stress Application
- Oxidative stress: Nimodipine or nifedipine pre-treated and control cells were treated with 2% EtOH.
- Osmotic stress: Nimodipine or nifedipine pre-treated and control cells were treated with 150 mM sodium chloride (NaCl).
- Heat: Nimodipine or nifedipine pre-treated and control cells have been incubated at 42 °C for 4 h or 6 h.
- Mechanical stress: Nimodipine or nifedipine pre-treated and control cells were shaken with two 2 mm steel beads at 500 rpm for 30 s. Afterwards steel beads were magnetically removed.
4.5. Lactate Dehydrogenase (LDH) Assay
4.6. RNA Isolation and cDNA Synthesis
4.7. qPCR
4.8. Protein Preparation
4.9. Protein Assay
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- National Institute for Health and Clinical Excellence (NICE). Hypertension in Adults: Diagnosis and Management. Clinical Guideline CG127. Available online: https://www.nice.org.uk/guidance/cg127 (accessed on 30 November 2016).
- De Lemons, J.A.; O’Rourke, R.A. Unstable angina and non-ST-segment elevation myocardial infarction. In Hurst’s the Heart; Fuster, V., Ed.; McGraw-Hill: New York, NY, USA, 2008; pp. 1371–1372. [Google Scholar]
- Thompson, A.E.; Pope, J.E. Calcium channel blockers for primary Raynaud’s phenomenon: A meta-analysis. Rheumatology 2005, 44, 145–150. [Google Scholar] [CrossRef] [PubMed]
- King, J.F.; Flenady, V.J.; Papatsonis, D.N.; Dekker, G.A.; Carbonne, B. Calcium channel blockers for inhibiting preterm labour. Cochrane Database Syst. Rev. 2003, 1, CD002255. [Google Scholar]
- Dorhout Mees, S.; Rinkel, G.J.; Feigin, V.L.; Algra, A.; van den Bergh, W.M.; Vermeulen, M.; van Gijn, J. Calcium antagonists for aneurysmal subarachnoid haemorrhage. Cochrane Database Syst. Rev. 2007, 18, CD000277. [Google Scholar]
- Rabinstein, A.A.; Lanzino, G.; Wijdicks, E.F. Multidisciplinary management and emerging therapeutic strategies in aneurysmal subarachnoid haemorrhage. Lancet Neurol. 2010, 9, 504–519. [Google Scholar] [CrossRef]
- Hydman, J.; Remahl, S.; Björck, G.; Svensson, M.; Mattsson, P. Nimodipine improves reinnervation and neuromuscular function after injury to the recurrent laryngeal nerve in the rat. Ann. Otol. Rhinol. Laryngol. 2007, 116, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Kansu, L.; Ozkarakas, H.; Efendi, H.; Okar, I. Protective effects of pentoxifylline and nimodipine on acoustic trauma in Guinea pig cochlea. Otol. Neurotol. 2011, 32, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, K.; Kumai, Y.; Sanuki, T.; Minoda, R.; Yumoto, E. The impact of nimodipine administration combined with nerve-muscle pedicle implantation on long-term denervated rat thyroarytenoid muscle. Laryngoscope 2013, 123, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.D.; Zheng, X.S.; Ying, T.T.; Yuan, Y.; Li, S.T. Nimodipine-mediated re-myelination after facial nerve crush injury in rats. J. Clin. Neurosci. 2005, 22, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, B.; Romstöck, J.; Fahlbusch, R.; Buchfelder, M.; Strauss, C. Intraoperative brainstem auditory evoked potential pattern and perioperative vasoactive treatment for hearing preservation in vestibular schwannoma surgery. J. Neurol. Neurosurg. Psychiatry 2008, 79, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.L.; Park, C. A prospective, randomized trial for use of prednisolone in patients with facial nerve paralysis after parotidectomy. Am. J. Surg. 2008, 196, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Scheller, C.; Strauss, C.; Fahlbusch, R.; Romstöck, J. Delayed facial nerve paresis following acoustic neuroma resection and postoperative vasoactive treatment. Zent. Neurochir. 2004, 65, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Scheller, C.; Richter, H.P.; Engelhardt, M.; Köenig, R.; Antoniadis, G. The influence of prophylactic vasoactive treatment on cochlear and facial nerve functions after vestibular schwannoma surgery: A prospective and open-label randomized pilot study. Neurosurgery 2007, 61, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Scheller, C.; Wienke, A.; Tatagiba, M.; Gharabaghi, A.; Ramina, K.F.; Ganslandt, O.; Bischoff, B.; Zenk, J.; Engelhorn, T.; Matthies, C.; et al. Prophylactic nimodipine treatment for cochlear and facial nerve preservation after vestibular schwannoma surgery: A randomized multicenter Phase III trial. J. Neurosurg. 2016, 124, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Scheller, K.; Scheller, C. Nimodipine promotes regeneration of peripheral facial nerve function after traumatic injury following maxillofacial surgery: An off label pilot-study. J. Craniomaxillofac. Surg. 2012, 40, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Nuglisch, J.; Karkoutly, C.; Mennel, H.D.; Roßberg, C.; Krieglstein, J. Protective effect of nimodipine against ischemic neuronal damage in rat hippocampus without changing postischemic cerebral blood flow. J. Cereb. Blood Flow Metab. 1990, 10, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Daschil, N.; Humpel, C. Nifedipine and nimodipine protect dopaminergic substantia nigra neurons against axotomy-induced cell death in rat vibrosections via modulating inflammatory responses. Brain Res. 2014, 1581, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lecht, S.; Rotfeld, E.; Arien-Zakay, H.; Tabakman, R.; Matzner, H.; Yaka, R.; Lelkes, P.I.; Lazarovici, P. Neuroprotective effects of nimodipine and nifedipine in the NGF-differentiated PC12 cells exposed to oxygen-glucose deprivation or trophic withdrawal. Int. J. Dev. Neurosci. 2012, 30, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Herzfeld, E.; Strauss, C.; Simmermacher, S.; Bork, K.; Horstkorte, R.; Dehghani, F.; Scheller, C. Investigation of the neuroprotective impact of nimodipine on Neuro2a cells by means of a surgery-like stress model. Int. J. Mol. Sci. 2014, 15, 18453–18465. [Google Scholar] [CrossRef] [PubMed]
- Boggs, J.M.; Koshy, K.M.; Rangaraj, G. Influence of structural modifications on the phase behavior of semi-synthetic cerebroside sulfate. Biochim. Biophys. Acta 1988, 938, 361–372. [Google Scholar] [CrossRef]
- Löfgren, H.; Pascher, I. Molecular arrangements of sphingolipids. The monolayer behaviour of ceramides. Chem. Phys. Lipids 1977, 20, 273–284. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Alderson, N.L.; Monje, P.V.; Wood, P.M.; Hama, H. FA2H is responsible for the formation of 2-hydroxy galactolipids in peripheral nervous system myelin. J. Lipid Res. 2008, 49, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Pascher, I. Molecular arrangements in sphingolipids. Conformation and hydrogen bonding of ceramide and their implication on membrane stability and permeability. Biochim. Biophys. Acta 1976, 455, 433–451. [Google Scholar] [CrossRef]
- Stewart, R.J.; Boggs, J.M. A carbohydrate-carbohydrate interaction between galactosylceramide-containing liposomes and cerebroside sulfate-containing liposomes: Dependence on the glycolipid ceramide composition. Biochemistry 1993, 32, 10666–10674. [Google Scholar] [CrossRef] [PubMed]
- Ljunggren, B.; Brandt, L.; Säveland, H.; Nilsson, P.E.; Cronqvist, S.; Andersson, K.E.; Vinge, E. Outcome in 60 consecutive patients treated with early aneurysm operation and intravenous nimodipine. J. Neurosurg. 1984, 61, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Sittel, C.; Sittel, A.; Guntinas-Lichius, O.; Eckel, H.E.; Stennert, E. Bell’s palsy: A 10-year experience with antiphlogistic-rheologic infusion therapy. Am. J. Otol. 2000, 21, 425–432. [Google Scholar] [CrossRef]
- Garrido, C.; Gurbuxani, S.; Ravagnan, L.; Kroemer, G. Heat shock proteins: Endogenous modulators of apoptotic cell death. Biochem. Biophys. Res. Commun. 2011, 286, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Kasza, Á.; Hunya, Á.; Frank, Z.; Fülöp, F.; Török, Z.; Balogh, G.; Sántha, M.; Bálind, Á.; Bernáth, S.; Blundell, K.L.; et al. Dihydropyridine Derivatives Modulate Heat Shock Responses and have a Neuroprotective Effect in a Transgenic Mouse Model of Alzheimer’s Disease. J. Alzheimers Dis. 2016, 53, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, P.J.; Van Assouw, H.P.; Peeters, L.; Leysen, J.E. Neurotoxic action of veratridine in rat brain neuronal cultures: Mechanism of neuroprotection by Ca2+ antagonists nonselective for slow Ca2+ channels. J. Pharmacol. Exp. Ther. 1990, 255, 1117–1122. [Google Scholar] [CrossRef]
- Hoshi, M.; Williams, M.; Kishimoto, Y. Characterization of brain cerebrosides at early stages of development in the rat. J. Neurochem. 1973, 21, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, Y.; Radin, N.S. Isolation and determination methods for brain cerebrosides, hydroxy fatty acids, and unsaturated and saturated fatty acids. J. Lipid Res. 1959, 1, 72–78. [Google Scholar]
- Kota, V.; Hama, H. 2′-Hydroxy ceramide in membrane homeostasis and cell signaling. Adv. Biol. Regul. 2014, 54, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Alderson, N.L.; Maldonado, E.N.; Kern, M.J.; Bhat, N.R.; Hama, H. FA2H-dependent fatty acid 2-hydroxylation in postnatal mouse brain. J. Lipid Res. 2006, 47, 2772–2780. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, M.; Yaghootfam, A.; Fewou, S.N.; Zöller, I.; Gieselmann, V. A mammalian fatty acid hydroxylase responsible for the formation of alpha-hydroxylated galactosylceramide in myelin. Biochem. J. 2005, 388, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Potter, K.A.; Kern, M.J.; Fullbright, G.; Bielawski, J.; Scherer, S.S.; Yum, S.W.; Li, J.J.; Cheng, H.; Han, X.; Venkata, J.K.; et al. Central nervous system dysfunction in a mouse model of FA2H deficiency. Glia 2011, 59, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Zöller, I.; Meixner, M.; Hartmann, D.; Büssow, H.; Meyer, R.; Gieselmann, V.; Eckhardt, M. Absence of 2-hydroxylated sphingolipids is compatible with normal neural development but causes late-onset axon and myelin sheath degeneration. J. Neurosci. 2008, 28, 9741–9754. [Google Scholar] [CrossRef] [PubMed]
- Edvardson, S.; Hama, H.; Shaag, A.; Gomori, J.M.; Berger, I.; Soffer, D.; Korman, S.H.; Taustein, I.; Saada, A.; Elpeleg, O. Mutations in the fatty acid 2-hydroxylase gene are associated with leukodystrophy with spastic paraparesis and dystonia. Am. J. Hum. Genet. 2008, 83, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; David, G. Stimulation-induced Ca2+ influx at nodes of Ranvier in mouse peripheral motor axons. J. Physiol. 2016, 594, 39–57. [Google Scholar] [CrossRef] [PubMed]
- McMurran, C.E.; Jones, C.A.; Fitzgerald, D.C.; Franklin, R.J. CNS Remyelination and the Innate Immune System. Front. Cell Dev. Biol. 2016, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.J.; Goldman, S.A. Glia Disease and Repair-Remyelination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020594. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.M.; Robinson, A.P.; Miller, S.D. Strategies for protecting oligodendrocytes and enhancing remyelination in multiple sclerosis. Discov. Med. 2013, 16, 53–63. [Google Scholar] [PubMed]
- Harlow, D.E.; Honce, J.M.; Miravalle, A.A. Remyelination Therapy in Multiple Sclerosis. Front. Neurol. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, R.G.; Sikorska, M.; Sandhu, J.K.; Lanthier, P.; Ribecco-Lutkiewicz, M.; Bani-Yaghoub, M. Differentiation of mouse Neuro 2A cells into dopamine neurons. J. Neurosci. Methods 2010, 186, 60–67. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide | Sequence (5′→3′) | Tm (°C) | Fragment Size |
|---|---|---|---|
| qFA2H_fwd | GGAGAAGTATGATGAGTGGGTTC | 62 | 295 bp |
| qFA2H_rev | GAAGCGGTGGATGACGTATT | 62 | |
| qGAPDH_fwd | GGAGAAACCTGCCAAGTATGA | 62 | 223 bp |
| qGAPDH_rev | CCTGTTGCTGTAGCCGTATT | 62 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herzfeld, E.; Speh, L.; Strauss, C.; Scheller, C. Nimodipine but Not Nifedipine Promotes Expression of Fatty Acid 2-Hydroxylase in a Surgical Stress Model Based on Neuro2a Cells. Int. J. Mol. Sci. 2017, 18, 964. https://doi.org/10.3390/ijms18050964
Herzfeld E, Speh L, Strauss C, Scheller C. Nimodipine but Not Nifedipine Promotes Expression of Fatty Acid 2-Hydroxylase in a Surgical Stress Model Based on Neuro2a Cells. International Journal of Molecular Sciences. 2017; 18(5):964. https://doi.org/10.3390/ijms18050964
Chicago/Turabian StyleHerzfeld, Eva, Lea Speh, Christian Strauss, and Christian Scheller. 2017. "Nimodipine but Not Nifedipine Promotes Expression of Fatty Acid 2-Hydroxylase in a Surgical Stress Model Based on Neuro2a Cells" International Journal of Molecular Sciences 18, no. 5: 964. https://doi.org/10.3390/ijms18050964
APA StyleHerzfeld, E., Speh, L., Strauss, C., & Scheller, C. (2017). Nimodipine but Not Nifedipine Promotes Expression of Fatty Acid 2-Hydroxylase in a Surgical Stress Model Based on Neuro2a Cells. International Journal of Molecular Sciences, 18(5), 964. https://doi.org/10.3390/ijms18050964