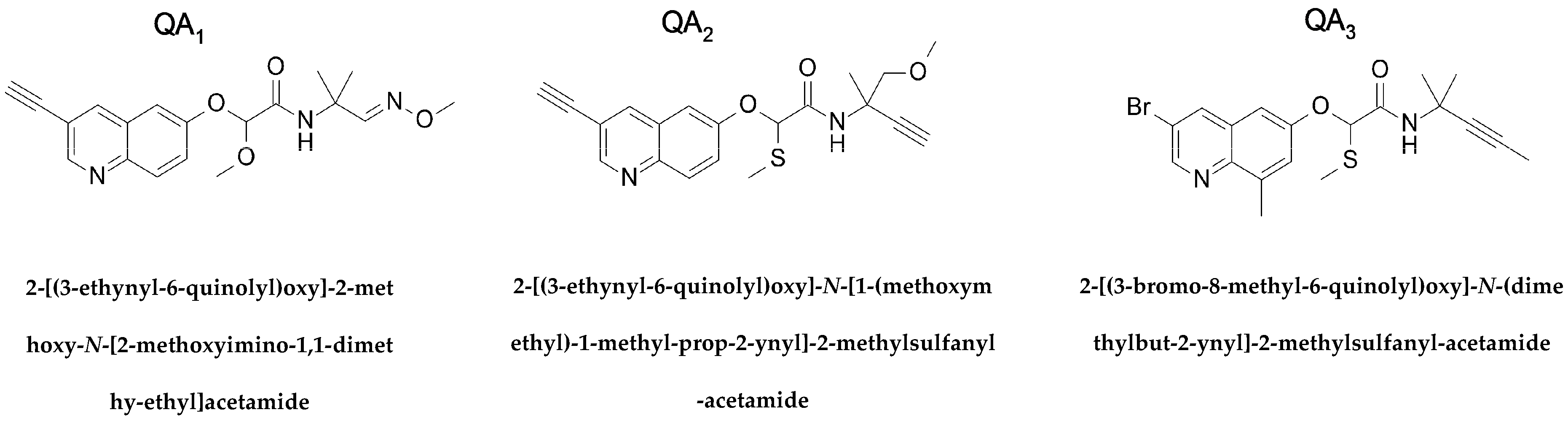
Quinolin-6-Yloxyacetamides Are Microtubule Destabilizing Agents That Bind to the Colchicine Site of Tubulin

 and
and
Abstract
:1. Introduction
2. Results
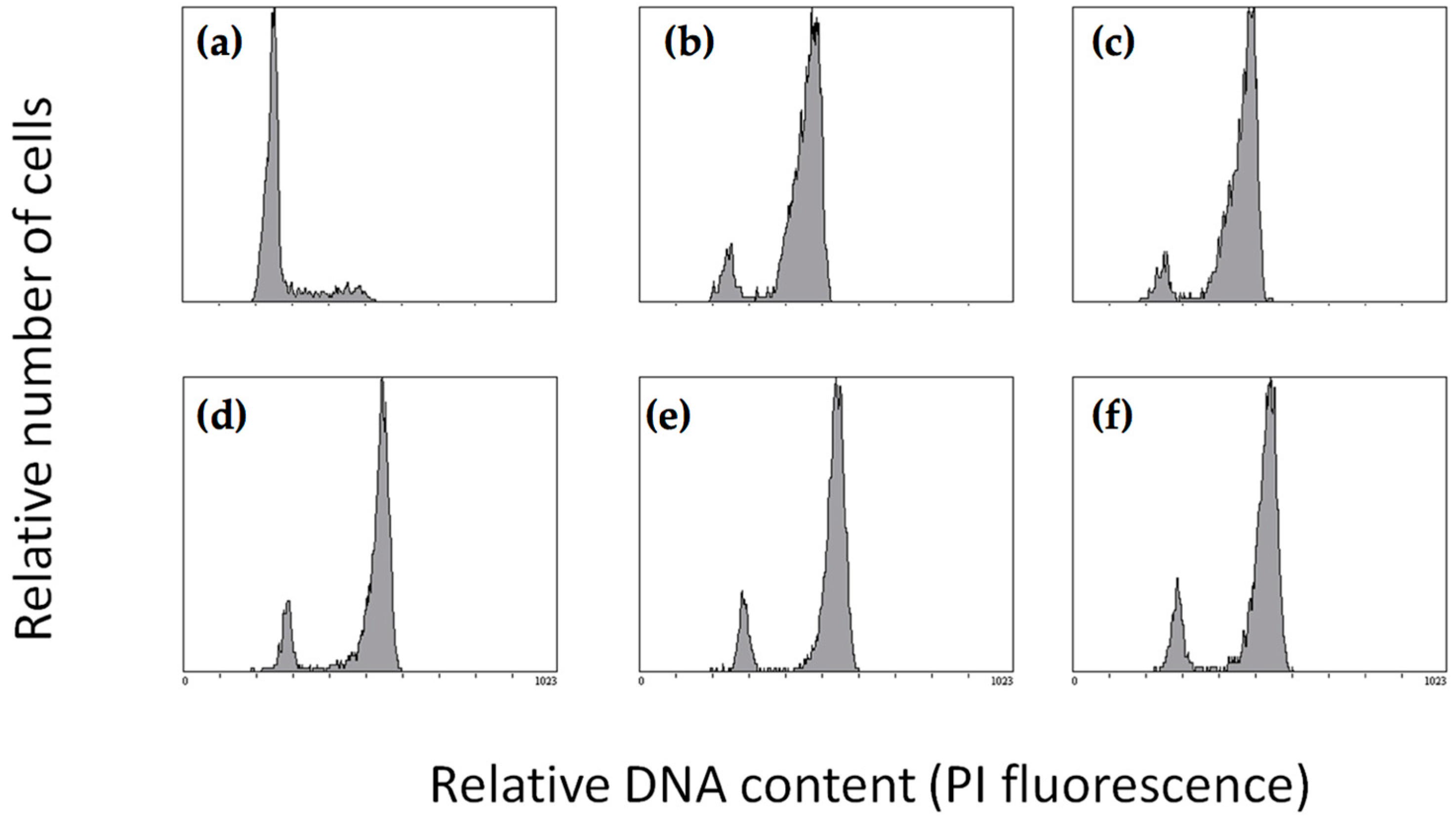
2.1. Effect of Quinolin-6-Yloxyacetamides on Cells
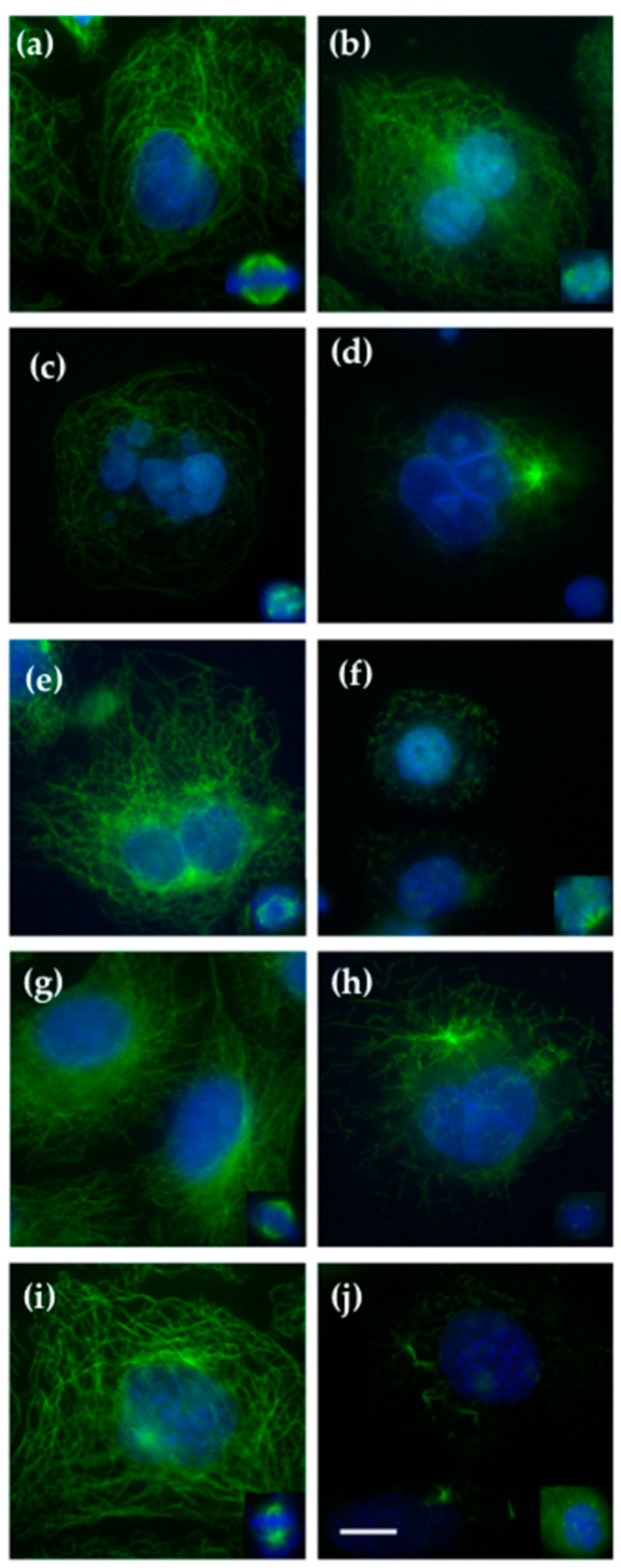
2.2. Effect on Cellular Microtubules
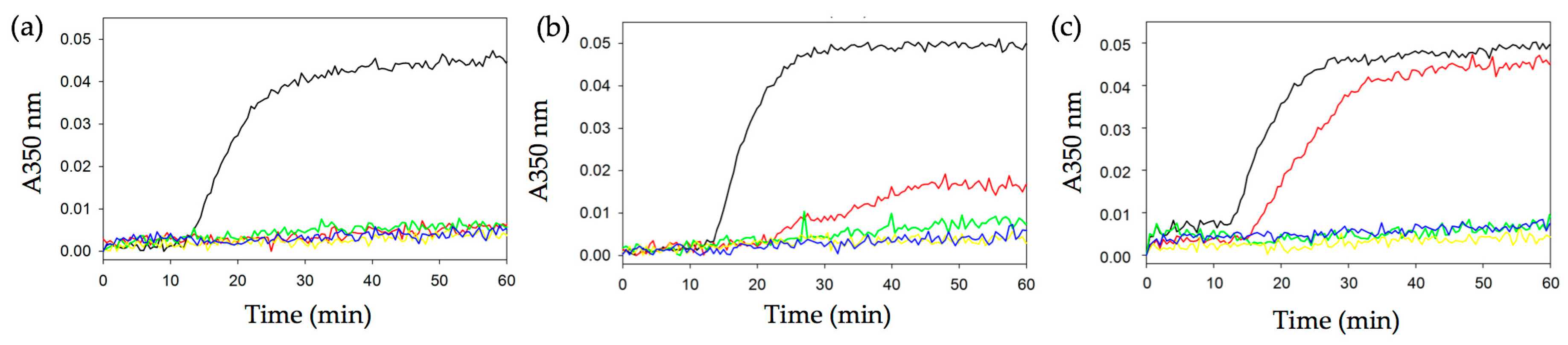
2.3. Effect of Quinolin-6-Yloxyacetamides on Microtubule Formation In Vitro
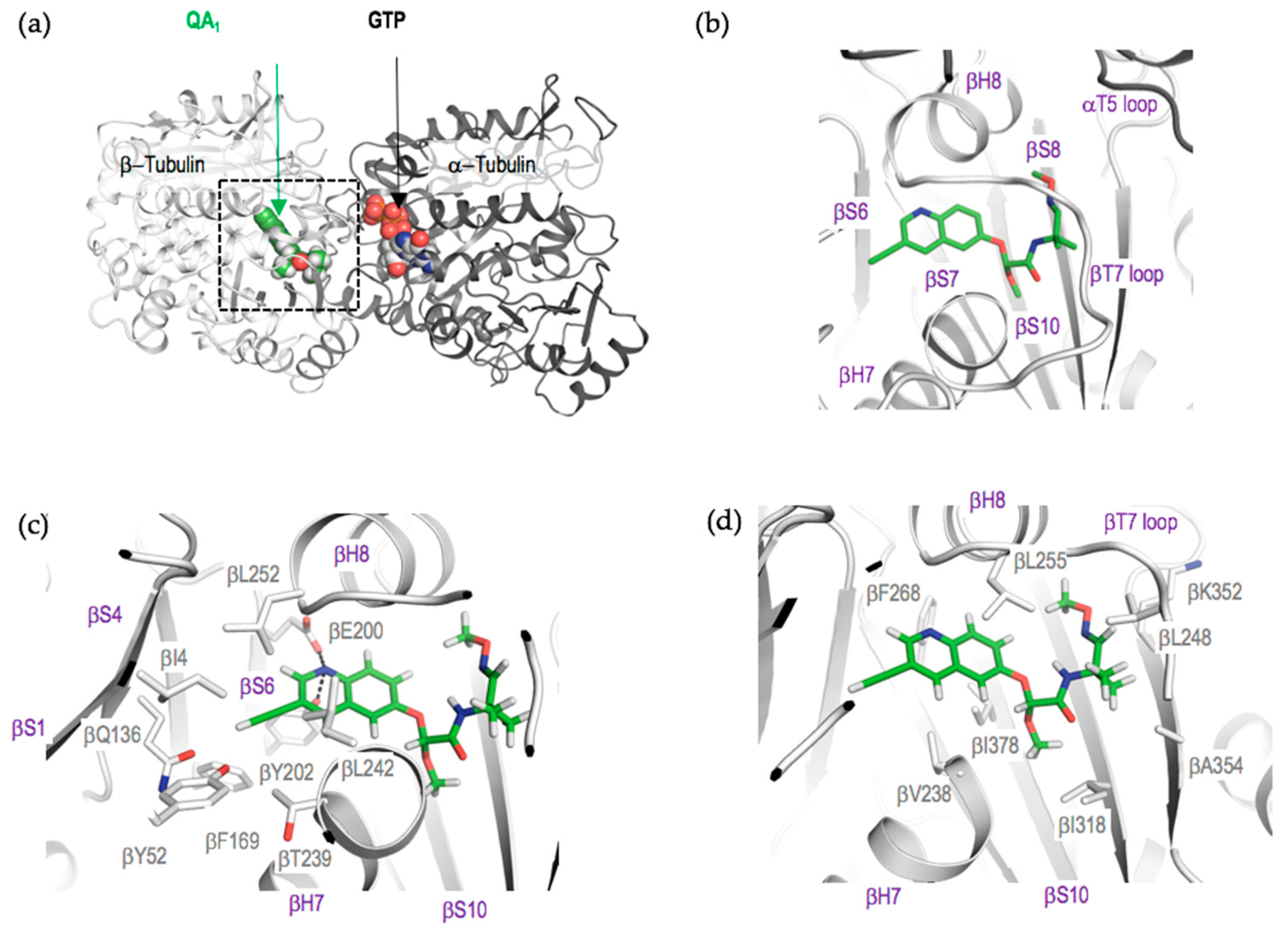
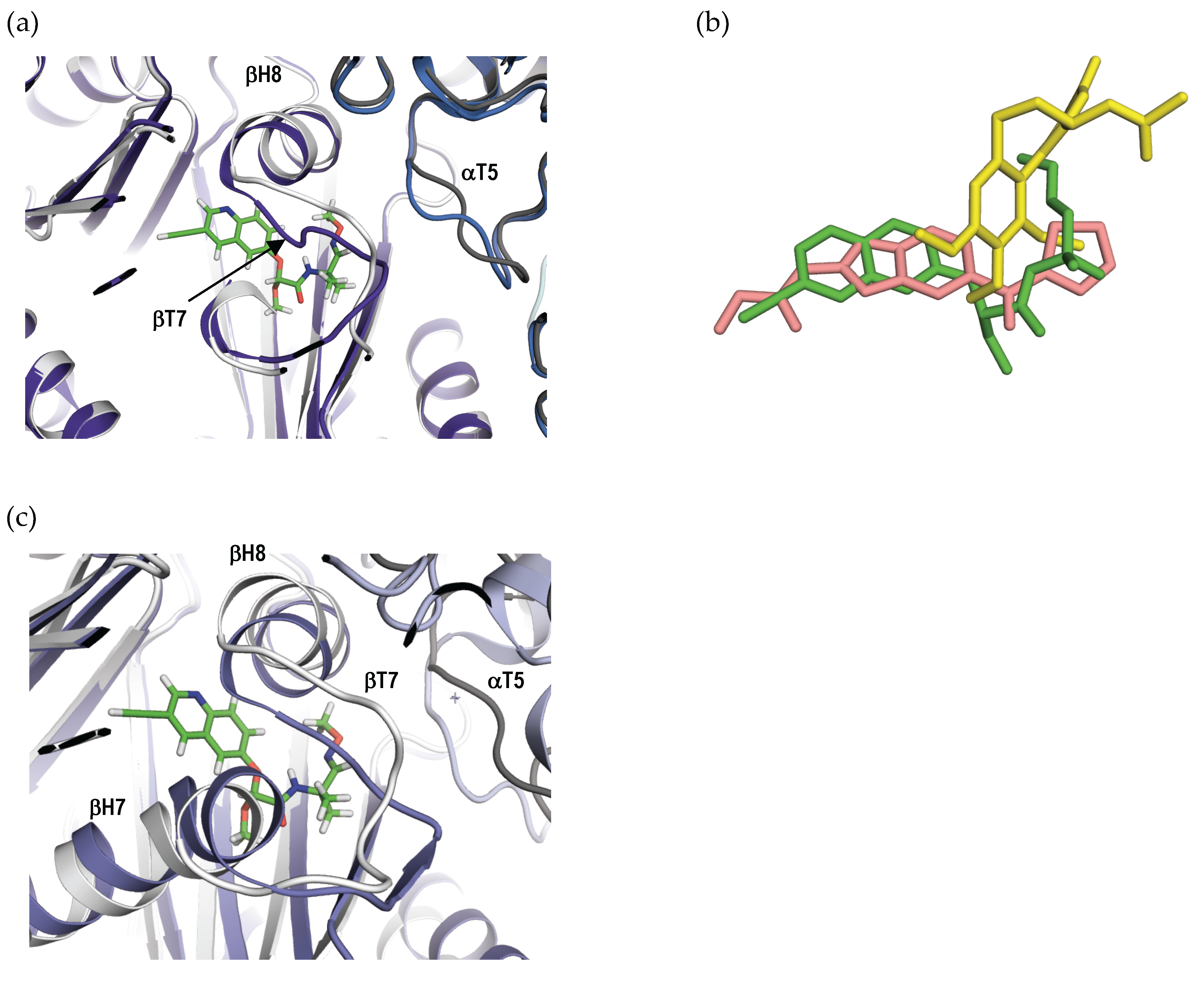
2.4. Crystal Structure of the Tubulin-QA1 Complex
3. Discussion
4. Materials and Methods
4.1. Proteins and Ligands
4.2. Tubulin Polymerization Assay
4.3. Cell Culture and Cell Biology Assays
4.4. Complex Reconstitution, Crystallization and Soaking of T2R-TTL (Tubulin-RB3-Tubulin Tyrosine Ligase Complex) Crystals with QA1
4.5. Data Collection and Structure Solution
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| QAs | Quinolin-6-yloxyacetamides |
| MT | Microtubules |
| MSA | Microtubule stabilizing agent |
| MDA | Microtubule destabilizing agent |
References
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of microtubule assembly in vitro by taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Akhmanova, A.; Steinmetz, M.O. Control of microtubule organization and dynamics: Two ends in the limelight. Nat. Rev. Mol. Cell Biol. 2015, 16, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Dorleans, A.; Gigant, B.; Ravelli, R.B.G.; Mailliet, P.; Mikol, V.; Knossow, M. Variations in the colchicine-binding domain provide insight into the structural switch of tubulin. Proc. Natl. Acad. Sci. USA 2009, 106, 13775–13779. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Bargsten, K.; Zurwerra, D.; Field, J.J.; Diaz, J.F.; Altmann, K.-H.; Steinmetz, M.O. Molecular mechanism of action of microtubule-stabilizing anticancer agents. Science 2013, 339, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Alushin, G.M.; Lander, G.C.; Kellogg, E.H.; Zhang, R.; Baker, D.; Nogales, E. High-resolution microtubule structures reveal the structural transitions in αβ-tubulin upon GTP hydrolysis. Cell 2014, 157, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Bargsten, K.; Northcote, P.T.; Marsh, M.; Altmann, K.-H.; Miller, J.H.; Diaz, J.F.; Steinmetz, M.O. Structural basis of microtubule stabilization by laulimalide and peloruside A. Angew. Chem. Int. Ed. Engl. 2014, 53, 1621–1625. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Bargsten, K.; Diaz, J.F.; Marsh, M.; Cuevas, C.; Liniger, M.; Neuhaus, C.; Andreu, J.M.; Altmann, K.-H.; Steinmetz, M.O. A new tubulin-binding site and pharmacophore for microtubule-destabilizing anticancer drugs. Proc. Natl. Acad. Sci. USA 2014, 111, 13817–13821. [Google Scholar] [CrossRef] [PubMed]
- Doodhi, H.; Prota, A.E.; Rodriguez-Garcia, R.; Xiao, H.; Custar, D.W.; Bargsten, K.; Katrukha, E.A.; Hilbert, M.; Hua, S.; Jiang, K.; et al. Termination of protofilament elongation by eribulin induces lattice defects that promote microtubule catastrophes. Curr. Biol. 2016, 26, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Danel, F.; Bachmann, F.; Bargsten, K.; Buey, R.M.; Pohlmann, J.; Reinelt, S.; Lane, H.; Steinmetz, M.O. The novel microtubule-destabilizing drug BAL27862 binds to the colchicine site of tubulin with distinct effects on microtubule organization. J. Mol. Biol. 2014, 426, 1848–1860. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B. Antibody-drug conjugates for cancer: Poised to deliver? Nat. Publ. Group 2010, 9, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, H.; Viskov, C.; Garcia-Echeverria, C. Antibody-drug conjugates-a new wave of cancer drugs. Bioorg. Med. Chem. Lett. 2014, 24, 5357–5363. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.M. Antibody-drug conjugates: Targeted delivery and future prospects. Ther. Deliv. 2016, 7, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Kizaki, M.; Hashimoto, Y. New tubulin polymerization inhibitor derived from thalidomide: Implications for anti-myeloma therapy. Curr. Med. Chem. 2008, 15, 754–765. [Google Scholar] [CrossRef] [PubMed]
- Rozic, G.; Paukov, L.; Jakubikova, J.; Ben-Shushan, D.; Duek, A.; Leiba, A.; Avigdor, A.; Nagler, A.; Leiba, M. The novel compound STK405759 is a microtubule-targeting agent with potent and selective cytotoxicity against multiple myeloma in vitro and in vivo. Oncotarget 2016, 7, 62572–62584. [Google Scholar] [CrossRef] [PubMed]
- Fryknas, M.; Gullbo, J.; Wang, X.; Rickardson, L.; Jarvius, M.; Wickstrom, M.; Hassan, S.; Andersson, C.; Gustafsson, M.; Westman, G.; et al. Screening for phenotype selective activity in multidrug resistant cells identifies a novel tubulin active agent insensitive to common forms of cancer drug resistance. BMC Cancer 2013, 13, 374. [Google Scholar] [CrossRef] [PubMed]
- Lamberth, C.; Kessabi, F.M.; Beaudegnies, R.; Quaranta, L.; Trah, S.; Berthon, G.; Cederbaum, F.; Knauf-Beiter, G.; Grasso, V.; Bieri, S.; et al. Synthesis and fungicidal activity of quinolin-6-yloxyacetamides, a novel class of tubulin polymerization inhibitors. Bioorg. Med. Chem. 2014, 22, 3922–3930. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.A.; Thrower, D.; Wilson, L. Effects of vinblastine, podophyllotoxin and nocodazole on mitotic spindles. Implications for the role of microtubule dynamics in mitosis. J. Cell Sci. 1992, 102, 401–416. [Google Scholar] [PubMed]
- Ravelli, R.B.G.; Gigant, B.; Curmi, P.A.; Jourdain, I.; Lachkar, S.; Sobel, A.; Knossow, M. Insight into tubulin regulation from a complex with colchicine and a stathmin-like domain. Nature 2004, 428, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Gigant, B.; Yu, Y.; Wu, Y.; Chen, X.; Lai, Q.; Yang, Z.; Chen, Q.; Yang, J. Structures of a diverse set of colchicine binding site inhibitors in complex with tubulin provide a rationale for drug discovery. FEBS J. 2016, 283, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Nogales, E.; Wolf, S.G.; Downing, K.H. Structure of the αβ tubulin dimer by electron crystallography. Nature 1998, 391, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Gigant, B.; Wang, C.; Ravelli, R.B.G.; Roussi, F.; Steinmetz, M.O.; Curmi, P.A.; Sobel, A.; Knossow, M. Structural basis for the regulation of tubulin by vinblastine. Nature 2005, 435, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Setter, J.; Waight, A.B.; Bargsten, K.; Murga, J.; Diaz, J.F.; Steinmetz, M.O. Pironetin binds covalently to αCys316 and perturbs a major loop and helix of α-tubulin to inhibit microtubule formation. J. Mol. Biol. 2016, 428, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, Y.; Wang, T.; Jiang, J.; Botting, C.H.; Liu, H.; Chen, Q.; Yang, J.; Naismith, J.H.; Zhu, X.; Chen, L. Pironetin reacts covalently with cysteine-316 of α-tubulin to destabilize microtubule. Nat. Commun. 2016, 7, 12103. [Google Scholar] [CrossRef] [PubMed]
- Skoufias, D.A.; Wilson, L. Mechanism of inhibition of microtubule polymerization by colchicine: Inhibitory potencies of unliganded colchicine and tubulin-colchicine complexes. Biochemistry 1992, 31, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Andreu, J.M. Large scale purification of brain tubulin with the modified Weisenberg procedure. Methods Mol. Med. 2007, 137, 17–28. [Google Scholar] [PubMed]
- Murphy Kessabi, F.; Brunner, H.-G.; Beaudegnies, R.; Quaranta, L.; Cederbaum, F. Novel Fungicides. WO 2009/030467, 12 March 2009. [Google Scholar]
- Beaudegnies, R.; Murphy Kessabi, F.; Quaranta, L.; Brunner, H.-G.; Cederbaum, F. Fungicidal 2-Alkylthio-2-quinolinyloxy-acetamide Derivatives. WO 2009/030469, 12 March 2009. [Google Scholar]
- Murphy Kessabi, F.; Quaranta, L.; Beaudegnies, R.; Brunner, H.-G.; Cederbaum, F. Fungicides. WO 2009/087098, 16 July 2009. [Google Scholar]
- Buey, R.M.; Calvo, E.; Barasoain, I.; Pineda, O.; Edler, M.C.; Matesanz, R.; Cerezo, G.; Vanderwal, C.D.; Day, B.W.; Sorensen, E.J.; et al. Cyclostreptin binds covalently to microtubule pores and lumenal taxoid binding sites. Nat. Chem. Biol. 2007, 3, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Buey, R.M.; Barasoain, I.; Jackson, E.; Meyer, A.; Giannakakou, P.; Paterson, I.; Mooberry, S.; Andreu, J.M.; Diaz, J.F. Microtubule interactions with chemically diverse stabilizing agents: Thermodynamics of binding to the paclitaxel site predicts cytotoxicity. Chem. Biol. 2005, 12, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Magiera, M.M.; Kuijpers, M.; Bargsten, K.; Frey, D.; Wieser, M.; Jaussi, R.; Hoogenraad, C.C.; Kammerer, R.A.; Janke, C.; et al. Structural basis of tubulin tyrosination by tubulin tyrosine ligase. J. Cell Biol. 2013, 200, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. Xds. Acta Crystallogr. Sect. Biol. Ldots 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Echols, N.; Headd, J.J.; Hung, L.-W.; Jain, S.; Kapral, G.J.; Grosse Kunstleve, R.W.; et al. The Phenix software for automated determination of macromolecular structures. Methods 2011, 55, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B., III; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | A549 | A2780 | A2780AD | R/S | 3T3 |
|---|---|---|---|---|---|
| QA1 | 60 ± 2 | 71 ± 14 | 141 ± 42 | 1.9 | 79 ± 5 |
| QA2 | 44.3 ± 11 | 104 ± 8 | 262 ± 85 | 2.5 | 125 ± 9 |
| QA3 | 707 ± 23 | 900 ± 100 | 800 ± 80 | 0.9 | 2400 ± 600 |
| Paclitaxel | 3.2 ± 1 | 1.2 ± 0.1 | 1100 ± 300 | 916 | ND |
| Colchicine | 55 ± 4 | 13.6 ± 2 | 663 ± 23 | 48 | 62.2 ± 4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Sáez-Calvo, G.; Olieric, N.; De Asís Balaguer, F.; Barasoain, I.; Lamberth, C.; Díaz, J.F.; Steinmetz, M.O. Quinolin-6-Yloxyacetamides Are Microtubule Destabilizing Agents That Bind to the Colchicine Site of Tubulin. Int. J. Mol. Sci. 2017, 18, 1336. https://doi.org/10.3390/ijms18071336
Sharma A, Sáez-Calvo G, Olieric N, De Asís Balaguer F, Barasoain I, Lamberth C, Díaz JF, Steinmetz MO. Quinolin-6-Yloxyacetamides Are Microtubule Destabilizing Agents That Bind to the Colchicine Site of Tubulin. International Journal of Molecular Sciences. 2017; 18(7):1336. https://doi.org/10.3390/ijms18071336
Chicago/Turabian StyleSharma, Ashwani, Gonzalo Sáez-Calvo, Natacha Olieric, Francisco De Asís Balaguer, Isabel Barasoain, Clemens Lamberth, J. Fernando Díaz, and Michel O. Steinmetz. 2017. "Quinolin-6-Yloxyacetamides Are Microtubule Destabilizing Agents That Bind to the Colchicine Site of Tubulin" International Journal of Molecular Sciences 18, no. 7: 1336. https://doi.org/10.3390/ijms18071336
APA StyleSharma, A., Sáez-Calvo, G., Olieric, N., De Asís Balaguer, F., Barasoain, I., Lamberth, C., Díaz, J. F., & Steinmetz, M. O. (2017). Quinolin-6-Yloxyacetamides Are Microtubule Destabilizing Agents That Bind to the Colchicine Site of Tubulin. International Journal of Molecular Sciences, 18(7), 1336. https://doi.org/10.3390/ijms18071336