Cellular Reparative Mechanisms of Mesenchymal Stem Cells for Retinal Diseases


Abstract
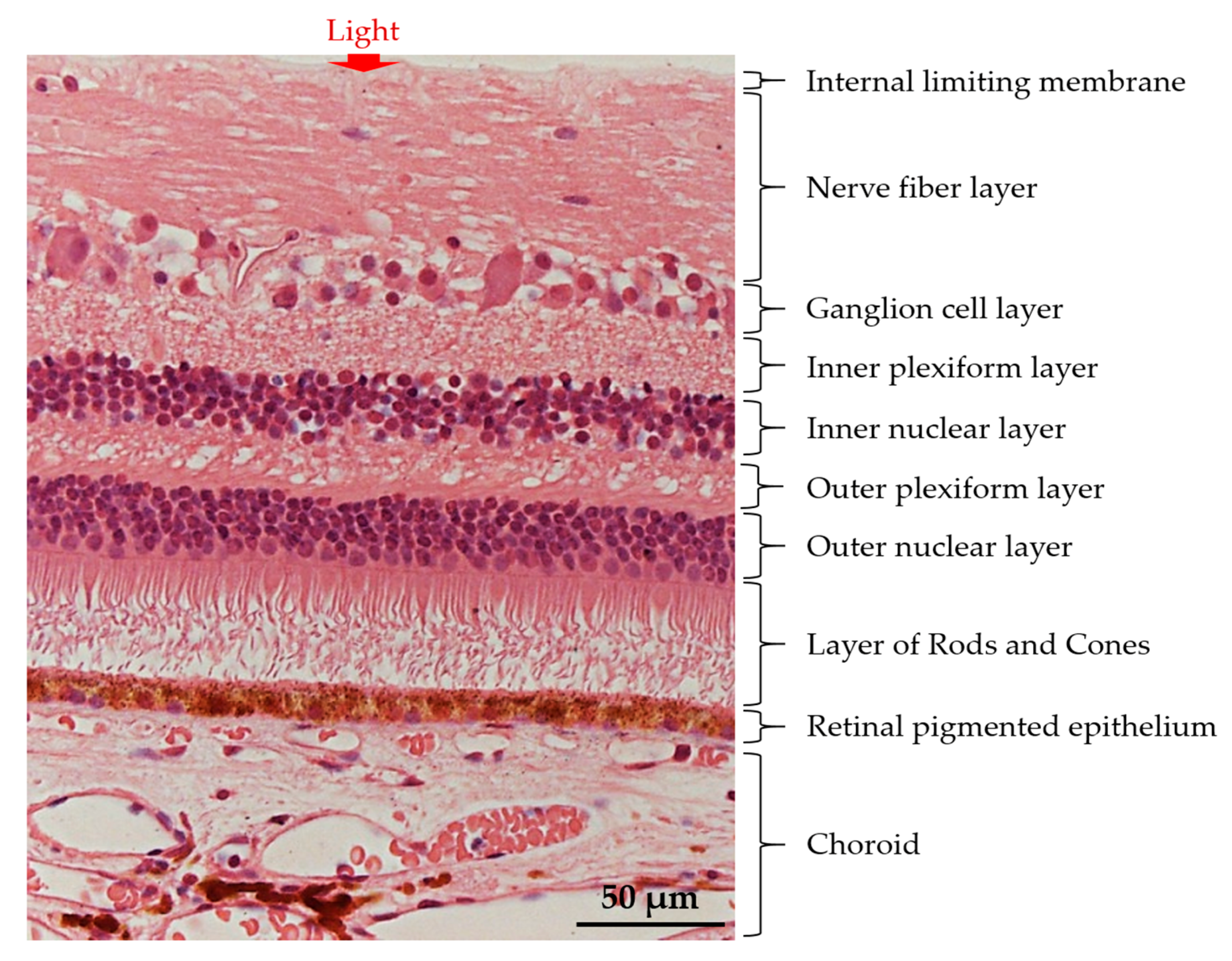
:1. Introduction
2. Current Therapeutic Approach for Retinal Diseases and Its Limitations
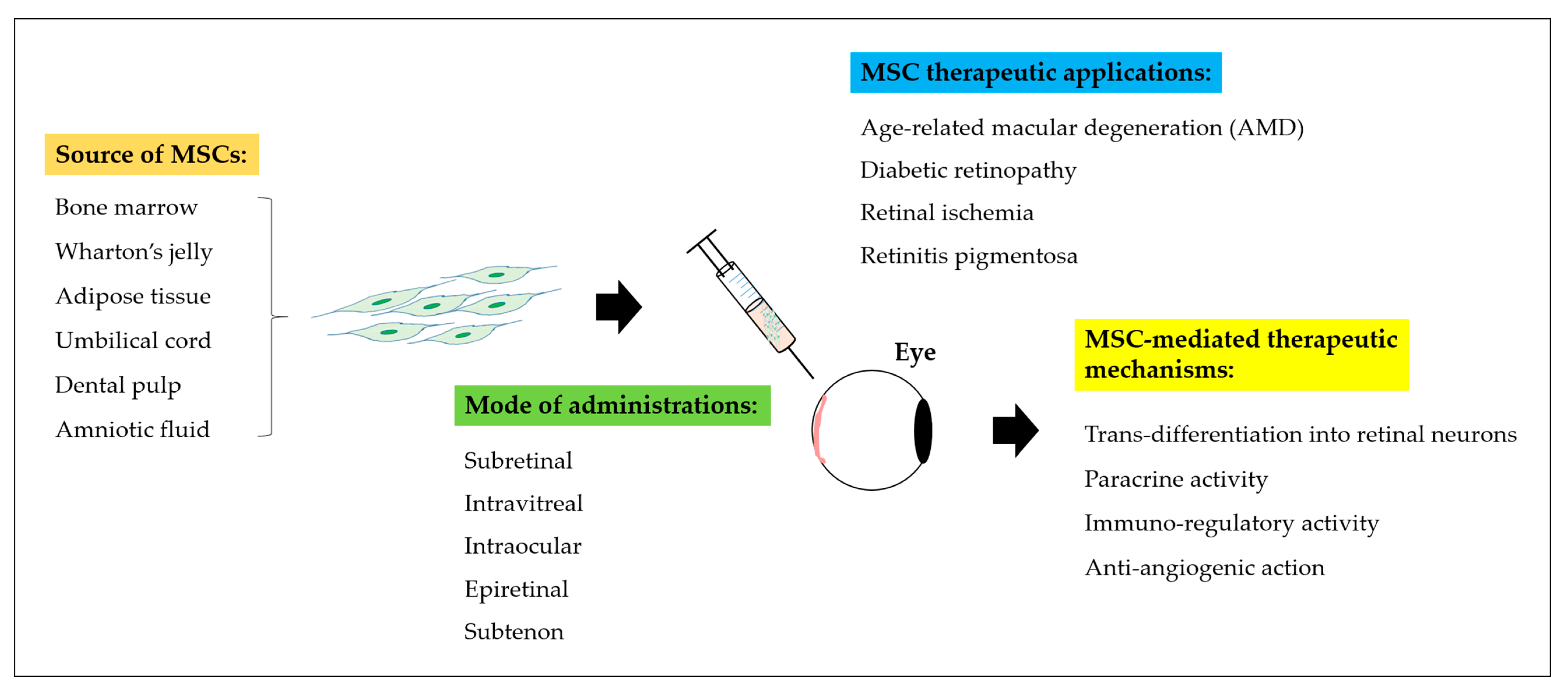
3. Alternative Therapeutic Strategies for Retinal Repair Using Stem Cell-Based Approach
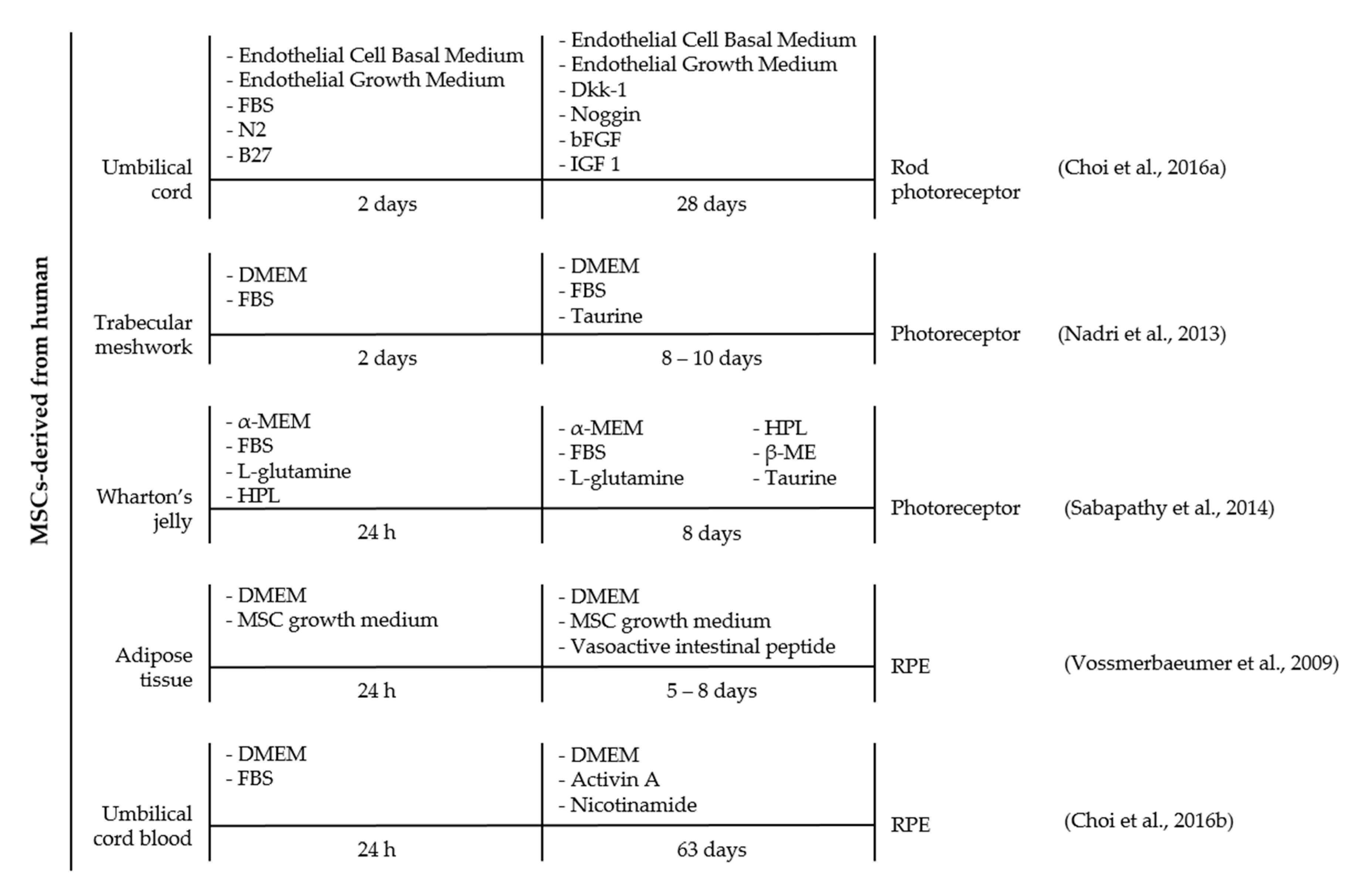
4. MSCs and Its Differentiation for the Treatment of Retinal Diseases
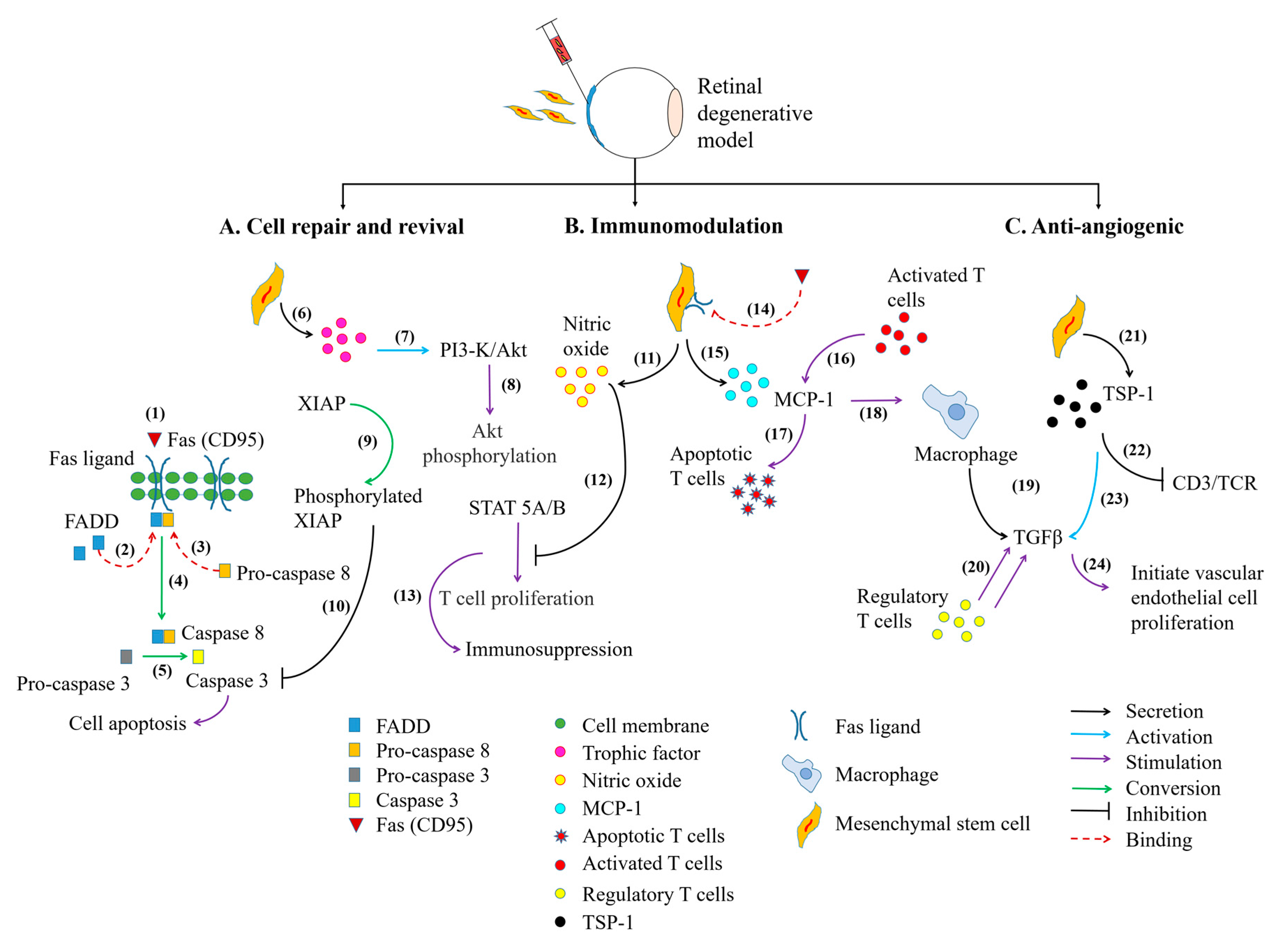
5. Paracrine Activity of MSCs Aids Retinal Cell Repair and Revival
6. MSCs Alleviates Inflammation in Retinal Diseases
7. MSCs Modulates Angiogenic Activity in Retinal Diseases
8. Conclusions
Acknowledgments
Author contributions
Conflicts of Interest
References
- Athanasiou, D.; Aguilà, M.; Bevilacqua, D.; Novoselov, S.S.; Parfitt, D.A.; Cheetham, M.E. The cell stress machinery and retinal degeneration. FEBS Lett. 2013, 587, 2008–2017. [Google Scholar] [CrossRef] [PubMed]
- Mannu, G.S. Retinal phototransduction. Neurosciences 2014, 19, 275–280. [Google Scholar] [PubMed]
- Reed, B.T.; Behar-Cohen, F.; Krantic, S. Seeing early signs of Alzheimer’s Disease through the lens of the eye. Curr. Alzheimer Res. 2016, 14, 6–17. [Google Scholar] [CrossRef]
- Palczewski, K. Chemistry and biology of vision. J. Biol. Chem. 2012, 287, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Triviño, A.; de Hoz, R.; Rojas, B.; Gallego, B.I.; Ramírez, A.I.; Salazar, J.J.; Ramírez, J.M. Effects of Hypercholesterolaemia in the Retina. In Ocular Diseases; InTech: Rijeka, Croatia, 2012. [Google Scholar] [Green Version]
- WHO (World Health Organization). WHO 2017. Available online: http://www.who.int/blindness/en/ (accessed on 8 June 2017).
- Weng, Y.; Liu, J.; Jin, S.; Guo, W.; Liang, X.; Hu, Z. Nanotechnology-based strategies for treatment of ocular disease. Acta Pharm. Sin. B 2017, 7, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Hernández, C.; Dal Monte, M.; Simó, R.; Casini, G. Neuroprotection as a therapeutic target for diabetic retinopathy. J. Diabetes Res. 2016, 2016, 9508541. [Google Scholar] [CrossRef] [PubMed]
- Narayan, D.S.; Wood, J.P.M.; Chidlow, G.; Casson, R.J. A review of the mechanisms of cone degeneration in retinitis pigmentosa. Acta Ophthalmol. 2016, 94, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H.; Xiao, X. Oxidative stress, innate immunity, and age-related macular degeneration. AIMS Mol. Sci. 2016, 3, 196–221. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Van Norren, D.; Vos, J.J. Light damage to the retina: An historical approach. Eye 2016, 30, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Cejkova, J.; Trosan, P.; Cejka, C.; Lencova, A.; Zajicova, A.; Javorkova, E.; Kubinova, S.; Sykova, E.; Holan, V. Suppression of alkali-induced oxidative injury in the cornea by mesenchymal stem cells growing on nanofiber scaffolds and transferred onto the damaged corneal surface. Exp. Eye Res. 2013, 116, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, F.; Cancarini, A.; dell’Omo, R.; Rezzola, S.; Romano, M.R.; Costagliola, C. Diabetic retinopathy: Vascular and inflammatory disease. J. Diabetes Res. 2015, 2015, 582060. [Google Scholar] [CrossRef] [PubMed]
- Chader, G.J.; Young, M. Preface: Sight restoration through stem cell therapy. Investig. Ophthalmol. Vis. Sci. 2016, 57, ORSFa1–ORSFa5. [Google Scholar] [CrossRef] [PubMed]
- Shirley Ding, S.L.; Leow, S.N.; Munisvaradass, R.; Koh, E.H.; Bastion, M.L.C.; Then, K.Y.; Kumar, S.; Mok, P.L. Revisiting the role of erythropoietin for treatment of ocular disorders. Eye 2016, 30, 1293–1309. [Google Scholar] [CrossRef] [PubMed]
- Klassen, H. Stem cells in clinical trials for treatment of retinal degeneration. Expert Opin. Biol. Ther. 2015, 16, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Villegas, V.M.; Aranguren, L.A.; Kovach, J.L.; Schwartz, S.G.; Flynn, H.W., Jr. Current advances in the treatment of neovascular age-related macular degeneration. Expert Opin. Drug Deliv. 2016, 14, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Adamis, A.P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Greenemeier, L. FDA approves first retinal implant. Nature 2013, 15, 26–28. [Google Scholar] [CrossRef]
- Da Cruz, L.; Dorn, J.D.; Humayun, M.S.; Dagnelie, G.; Handa, J.; Barale, P.O.; Sahel, J.A.; Stanga, P.E.; Hafezi, F.; Safran, A.B.; et al. Five-year safety and performance results from the Argus II Retinal Prosthesis System Clinical Trial. Ophthalmology 2016, 123, 2248–2254. [Google Scholar] [CrossRef] [PubMed]
- Rayess, N.; Houston, S.K.S.; Gupta, O.P.; Ho, A.C.; Regillo, C.D. Treatment outcomes after 3 years in neovascular age-related macular degeneration using a treat-and-extend regimen. Am. J. Ophthalmol. 2015, 159, 3–8.e1. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.A.; Glassman, A.R.; Ayala, A.R.; Jampol, L.M.; Bressler, N.M.; Bressler, S.B.; Brucker, A.J.; Ferris, F.L.; Hampton, G.R.; Jhaveri, C.; et al. Aflibercept, Bevacizumab, or Ranibizumab for Diabetic Macular Edema two-year results from a comparative effectiveness randomized clinical trial. Ophthalmology 2016, 123, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Boyer, D.S.; Hopkins, J.J.; Sorof, J.; Ehrlich, J.S. Anti-vascular endothelial growth factor therapy for diabetic macular edema. Ther. Adv. Endocrinol. Metab. 2013, 4, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. VEGF and intraocular neovascularization: From discovery to therapy. Transl. Vis. Sci. Technol. 2016, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.D.; Lindsley, K.; Vedula, S.S.; Krzystolik, M.G.; Hawkins, B.S. Anti-vascular endothelial growth factor for neovascular age-related macular degeneration. Cochrane Database Syst. Rev. 2014, 8, CD005139. [Google Scholar]
- Eguizabal, C.; Montserrat, N.; Veiga, A.; Belmonte, J.I. Dedifferentiation, transdifferentiation, and reprogramming: Future directions in regenerative medicine. Semin. Reprod. Med. 2013, 31, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, I.; Kolisis, F.N.; Papaevangeliou, D.; Zoumpourlis, V. Comparative evaluation of human mesenchymal stem cells of fetal (Wharton’s Jelly) and adult (adipose tissue) origin during prolonged in vitro expansion: Considerations for cytotherapy. Stem Cells Int. 2013, 2013, 246134. [Google Scholar] [CrossRef] [PubMed]
- Assawachananont, J.; Mandai, M.; Okamoto, S.; Yamada, C.; Eiraku, M.; Yonemura, S.; Sasai, Y.; Takahashi, M. Transplantation of embryonic and induced pluripotent stem cell-derived 3D retinal sheets into retinal degenerative mice. Stem Cell Rep. 2014, 2, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Rezanejad, H.; Soheili, Z.S.; Haddad, F.; Matin, M.M.; Samiei, S.; Manafi, A.; Ahmadieh, H. In vitro differentiation of adipose-tissue-derived mesenchymal stem cells into neural retinal cells through expression of human PAX6 (5a) gene. Cell Tissue Res. 2014, 356, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.K.; Yung, J.S.Y.; Choy, K.W.; Cao, D.; Leung, C.K.S.; Cheung, H.S.; Pang, C.P. Transdifferentiation of periodontal ligament-derived stem cells into retinal ganglion-like cells and its microRNA signature. Sci. Rep. 2015, 5, 16429. [Google Scholar] [CrossRef] [PubMed]
- Worthington, K.S.; Green, B.J.; Rethwisch, M.; Wiley, L.A.; Tucker, B.A.; Guymon, C.A.; Salem, A.K. Neuronal differentiation of induced pluripotent stem cells on surfactant templated chitosan hydrogels. Biomacromolecules 2016, 17, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- Nicoară, S.D.; Șușman, S.; Tudoran, O.; Bărbos, O.; Cherecheș, G.; Aștilean, S.; Potara, M.; Sorițău, O. Novel strategies for the improvement of stem cells’ transplantation in degenerative retinal diseases. Stem Cells Int. 2016, 2016, 1236721. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.D.; Regillo, C.D.; Lam, B.L.; Eliott, D.; Rosenfeld, P.J.; Gregori, N.Z.; Hubschman, J.P.; Davis, J.L.; Heilwell, G.; Spirn, M.; et al. Human embryonic stem cell-derived retinal pigment epithelium in patients with age-related macular degeneration and Stargardt’s macular dystrophy: Follow-up of two open-label phase 1/2 studies. Lancet 2015, 385, 509–516. [Google Scholar] [CrossRef]
- Schwartz, S.D.; Tan, G.; Hosseini, H.; Nagiel, A. Subretinal transplantation of embryonic stem cell-derived retinal pigment epithelium for the treatment of macular degeneration: An assessment at 4 years. Investig. Ophthalmol. Vis. Sci. 2016, 57, ORSFc1–ORSFc9. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Fukuda, K. Methods of induced pluripotent stem cells for clinical application. World J. Stem Cells 2015, 7, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induced pluripotent stem cells in medicine and biology. Development 2013, 140, 2457–2461. [Google Scholar] [CrossRef] [PubMed]
- Osakada, F.; Ikeda, H.; Mandai, M.; Wataya, T.; Watanabe, K.; Yoshimura, N.; Akaike, A.; Sasai, Y.; Takahashi, M. Toward the generation of rod and cone photoreceptors from mouse, monkey and human embryonic stem cells. Nat. Biotechnol. 2008, 26, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Alexander, P.; Thomson, H.A.J.; Luff, A.J.; Lotery, A.J. Retinal pigment epithelium transplantation: Concepts, challenges, and future prospects. Eye 2015, 29, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Zhao, H.; Feng, R.; Zhang, X.; Tai, D.; An, G.; Wen, J.; Tan, J. Efficient induction of pluripotent stem cells from menstrual blood. Stem Cells Dev. 2013, 22, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Nakano-Okuno, M.; Borah, B.R.; Nakano, I. Ethics of iPSC-Based clinical research for age-related macular degeneration: Patient-centered risk-benefit analysis. Stem Cell Rev. Rep. 2014, 10, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, A.; Miyagawa, S.; Fukushima, S.; Kawamura, T.; Kashiyama, N.; Ito, E.; Masuda, S.; Toda, K.; Hatazawa, J.; Morii, E. Teratocarcinomas arising from allogeneic induced pluripotent stem cell-derived cardiac tissue constructs provoked host immune rejection in mice. Sci. Rep. 2016, 6, 19464. [Google Scholar] [CrossRef] [PubMed]
- Itakura, G.; Kobayashi, Y.; Nishimura, S.; Iwai, H.; Takano, M.; Iwanami, A.; Toyama, Y.; Okano, H.; Nakamura, M. Controlling immune rejection is a fail-safe system against potential tumorigenicity after human iPSC-derived neural stem cell transplantation. PLoS ONE 2015, 10, e0116413. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-T.; Luo, Y.-H.; Chien, C.-S.; Wu, C.-H.; Tseng, P.-C.; Chiou, S.-H.; Lee, Y.C.; Whang-Peng, J.; Chen, Y.M. Induced pluripotent stem cell–conditioned medium suppressed melanoma tumorigenicity through the enhancement of natural-killer cellular immunity. J. Immunother. 2016, 39, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Scheiner, Z.S.; Talib, S.; Feigal, E.G. The potential for immunogenicity of autologous induced pluripotent stem cell-derived therapies. J. Biol. Chem. 2014, 289, 4571–4577. [Google Scholar] [CrossRef] [PubMed]
- Zarbin, M. Cell-based therapy for degenerative retinal disease. Trends Mol. Med. 2016, 22, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Song, W.K.; Park, K.M.; Kim, H.J.; Lee, J.H.; Choi, J.; Chong, S.Y.; Shim, S.H.; del Priore, L.V.; Lanza, R. Treatment of macular degeneration using embryonic stem cell-derived retinal pigment epithelium: Preliminary results in Asian patients. Stem Cell Rep. 2015, 4, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Mok, P.L.; Leong, C.F.; Cheong, S.K. Cellular mechanisms of emerging applications of mesenchymal stem cells. Malays. J. Pathol. 2013, 35, 17–32. [Google Scholar] [PubMed]
- Chung, S.; Rho, S.; Kim, G.; Kim, S.-R.; Baek, K.-H.; Kang, M.; Lew, H. Human umbilical cord blood mononuclear cells and chorionic plate-derived mesenchymal stem cells promote axon survival in a rat model of optic nerve crush injury. Int. J. Mol. Med. 2016, 37, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.; Dreixler, J.C.; Mathew, B.; Balyasnikova, I.; Mann, J.R.; Boddapoti, V.; Xue, L.; Lesniak, M.S. Hypoxic-Preconditioned bone marrow stem cell medium significantly improves outcome after retinal ischemia in rats. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3522–3532. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, M.; Urzua, C.A.; Montecino, S.; Leal, K.; Conget, P.; Ezquer, F. Intravitreal administration of multipotent mesenchymal stromal cells triggers a cytoprotective microenvironment in the retina of diabetic mice. Stem Cell Res. Ther. 2016, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Leow, S.N.; Luu, C.D.; Nizam, M.H.H.; Mok, P.L.; Ruhaslizan, R.; Wong, H.S.; Halim, W.H.W.A.; Ng, M.H.; Ruszymah, B.H.I.; Chowdhury, S.R.; et al. Safety and efficacy of human Wharton’s Jelly-derived mesenchymal stem cells therapy for retinal degeneration. PLoS ONE 2015, 10, e0128973. [Google Scholar] [CrossRef] [PubMed]
- Mead, B.; Hill, L.J.; Blanch, R.J.; Ward, K.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Mesenchymal stromal cell-mediated neuroprotection and functional preservation of retinal ganglion cells in a rodent model of glaucoma. Cytotherapy 2016, 18, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Park, J.M.; Kong, T.H.; Kim, C.; Bae, S.H.; Kim, H.W.; Moon, J. Retinal angiogenesis effects of TGF-β1 and paracrine factors secreted from human placental stem cells in response to a pathological environment. Cell Transplant. 2016, 25, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Ren, H.; Han, Z. Mesenchymal stem cells: Immunomodulatory capability and clinical potential in immune diseases. J. Cell. Immunother. 2016, 2, 3–20. [Google Scholar] [CrossRef]
- Cejka, C.; Holan, V.; Trosan, P.; Zajicova, A.; Javorkova, E.; Cejkova, J. The favorable effect of mesenchymal stem cell treatment on the antioxidant protective mechanism in the corneal epithelium and renewal of corneal optical properties changed after alkali burns. Oxid. Med. Cell. Longev. 2016, 2016, 5843809. [Google Scholar] [CrossRef] [PubMed]
- Schafer, R.; Spohn, G.; Baer, P.C. Mesenchymal stem/stromal cells in regenerative medicine: Can preconditioning strategies improve therapeutic efficacy? Transfus. Med. Hemother. 2016, 43, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Ma, Y.H.; Chen, Y.F.; Qiu, X.C.; Wu, J.L.; Ling, E.A.; Zeng, Y.S. Autocrine fibronectin from differentiating mesenchymal stem cells induces the neurite elongation in vitro and promotes nerve fiber regeneration in transected spinal cord injury. J. Biomed. Mater. Res. A 2016, 104, 1902–1911. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Nagaishi, K.; Konari, N.; Saito, Y.; Chikenji, T.; Mizue, Y.; Fujimiya, M. Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by exosome transfer into damaged neurons and astrocytes. Sci. Rep. 2016, 6, 24805. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.-T.; Zhang, L.-J.; Shao, H.; Bai, L.-L.; Yu, B.; Su, C.; Dong, L.J.; Liu, X.; Li, X.R.; Zhang, X.M. Therapeutic effects of mesenchymal stem cells administered at later phase of recurrent experimental autoimmune uveitis. Int. J. Ophthalmol. 2016, 9, 1381–1389. [Google Scholar] [PubMed]
- Guan, Y.; Cui, L.; Qu, Z.; Lu, L.; Wang, F.; Wu, Y.; Zhang, J.; Gao, F.; Tian, H.; Xu, L.; et al. Subretinal transplantation of rat MSCs and erythropoietin gene modified rat MSCs for protecting and rescuing degenerative retina in rats. Curr. Mol. Med. 2013, 13, 1419–1431. [Google Scholar] [CrossRef] [PubMed]
- Çerman, E.; Akkoç, T.; Eraslan, M.; Şahin, Ö.; Özkara, S.; Aker, F.V.; Subaşı, C.; Karaöz, E.; Akkoç, T. Retinal electrophysiological effects of intravitreal bone marrow derived mesenchymal stem cells in streptozotocin induced diabetic rats. PLoS ONE 2016, 11, e0156495. [Google Scholar]
- Gramlich, O.W.; Burand, A.J.; Brown, A.J.; Deutsch, R.J.; Kuehn, M.H.; Ankrum, J.A. Cryopreserved mesenchymal stromal cells maintain potency in a retinal ischemia/reperfusion injury model: Toward an off-the-shelf therapy. Sci. Rep. 2016, 6, 26463. [Google Scholar] [CrossRef] [PubMed]
- Tzameret, A.; Sher, I.; Belkin, M.; Treves, A.J.; Meir, A.; Nagler, A.; Levkovitch-Verbin, H.; Barshack, I.; Rosner, M.; Rotenstreich, Y. Transplantation of human bone marrow mesenchymal stem cells as a thin subretinal layer ameliorates retinal degeneration in a rat model of retinal dystrophy. Exp. Eye Res. 2014, 118, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Tzameret, A.; Sher, I.; Belkin, M.; Treves, A.J.; Meir, A.; Nagler, A.; Levkovitch-Verbin, H.; Rotenstreich, Y.; Solomon, A.S. Epiretinal transplantation of human bone marrow mesenchymal stem cells rescues retinal and vision function in a rat model of retinal degeneration. Stem Cell Res. 2015, 15, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Wang, S.; Girman, S.; McGill, T.; Ragaglia, V.; Lund, R. Human adult bone marrow-derived somatic cells rescue vision in a rodent model of retinal degeneration. Exp. Eye Res. 2010, 91, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Lund, R.D.; Wang, S.; Lu, B.; Girman, S.; Holmes, T.; Sauvé, Y.; Messina, D.J.; Harris, I.R.; Kihm, A.J.; Harmon, A.M.; et al. Cells isolated from umbilical cord tissue rescue photoreceptors and visual functions in a rodent model of retinal disease. Stem Cells 2009, 25, 602–611. [Google Scholar] [CrossRef]
- Yuan, J.; Yu, J.X. Gender difference in the neuroprotective effect of rat bone marrow mesenchymal cells against hypoxia-induced apoptosis of retinal ganglion cells. Neural Regen. Res. 2016, 11, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.; Boue, S.; Izpisua Belmonte, J.C. Dedifferentiation, transdifferentiation and reprogramming: Three routes to regeneration. Nat. Rev. Mol. Cell Biol. 2011, 12, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, H.; Kushida, Y.; Nojima, M.; Kuroda, Y.; Wakao, S.; Ishida, K.; Endo, F.; Kume, K.; Takahara, T.; Nitta, H.; et al. A distinct subpopulation of bone marrow mesenchymal stem cells, muse cells, directly commit to the replacement of liver components. Am. J. Transplant. 2016, 16, 468–483. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Shin, J.; Kim, J.; Shin, T.-H.; Seo, Y. Direct cell fate conversion of human somatic stem cells into cone and rod photoreceptor-like cells by inhibition of microRNA-203. Oncotarget 2016, 7, 42139–42149. [Google Scholar] [CrossRef] [PubMed]
- Nadri, S.; Yazdani, S.; Arefian, E.; Gohari, Z.; Eslaminejad, M.B.; Kazemi, B.; Soleimani, M. Mesenchymal stem cells from trabecular meshwork become photoreceptor-like cells on amniotic membrane. Neurosci. Lett. 2013, 541, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Vossmerbaeumer, U.; Ohnesorge, S.; Kuehl, S.; Haapalahti, M.; Kluter, H.; Jonas, J.B.; Thierse, H.J.; Bieback, K. Retinal pigment epithelial phenotype induced in human adipose tissue-derived mesenchymal stromal cells. Cytotherapy 2009, 11, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Sabapathy, V.; Sundaram, B.; Vm, S.; Mankuzhy, P.; Kumar, S. Human wharton’s jelly mesenchymal stem cells plasticity augments scar-free skin wound healing with hair growth. PLoS ONE 2014, 9, e93726. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Kim, J.-J.; Seo, M.-S.; Park, S.-B.; Shin, T.-H.; Shin, J.-H.; Seo, Y.; Kim, H.S.; Kang, K.S. Inhibition by miR-410 facilitates direct retinal pigment epithelium differentiation of umbilical cord blood-derived mesenchymal stem cells. J. Vet. Sci. 2016, 18, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.S.; Park, Y.-B.; Ha, C.-W.; Kim, J.A.; Heo, J.-C.; Han, W.-J.; Oh, S.Y.; Choi, S.J. Different characteristics of mesenchymal stem cells isolated from different layers of full term placenta. PLoS ONE 2017, 12, e0172642. [Google Scholar] [CrossRef] [PubMed]
- Divya, M.S.; Roshin, G.E.; Divya, T.S.; Rasheed, V.A.; Santhoshkumar, T.R.; Elizabeth, K.E.; James, J.; Pillai, R.M. Umbilical cord blood-derived mesenchymal stem cells consist of a unique population of progenitors co-expressing mesenchymal stem cell and neuronal markers capable of instantaneous neuronal differentiation. Stem Cell Res. Ther. 2012, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Trivanović, D.; Jauković, A.; Popović, B.; Krstić, J.; Mojsilović, S.; Okić-Djordjević, I.; Kukolj, T.; Obradović, H.; Santibanez, J.F.; Bugarski, D. Mesenchymal stem cells of different origin: Comparative evaluation of proliferative capacity, telomere length and pluripotency marker expression. Life Sci. 2015, 141, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.N.; Moschidou, D.; Puga-Iglesias, T.-I.; Kuleszewicz, K.; Vanleene, M.; Shefelbine, S.J.; Bou-Gharios, G.; Fisk, N.M.; David, A.L.; de Coppi, P.; et al. Ontological differences in first compared to third trimester human fetal placental chorionic stem cells. PLoS ONE 2012, 7, e43395. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Hu, J.; Zhao, J.; Liu, J.; Ouyang, W.; Yang, C.; Gong, N.; Du, L.; Khanal, A.; Chen, L. Side-by-side comparison of the biological characteristics of human umbilical cord and adipose tissue-derived mesenchymal stem cells. Biomed. Res. Int. 2013, 2013, 438243. [Google Scholar] [CrossRef] [PubMed]
- Dezawa, M. Muse cells provide the pluripotency of mesenchymal stem cells: Direct contribution of muse cells to tissue regeneration. Cell Transplant. 2016, 25, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Simerman, A.A.; Phan, J.D.; Dumesic, D.A.; Chazenbalk, G.D. Muse cells: Nontumorigenic pluripotent stem cells present in adult tissues—A paradigm shift in tissue regeneration and evolution. Stem Cells Int. 2016, 2016, 1463258. [Google Scholar] [CrossRef] [PubMed]
- Kanno, H. Regenerative therapy for neuronal diseases with transplantation of somatic stem cells. World J. Stem Cells 2013, 5, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Bakondi, B.; Girman, S.; Lu, B.; Wang, S. Multimodal delivery of isogenic mesenchymal stem cells yields synergistic protection from retinal degeneration and vision loss. Stem Cells Transl. Med. 2017, 6, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Xu, N.; Xu, W.; Xu, G. Mesenchymal stem cells attenuate hydrogen peroxide-induced oxidative stress and enhance neuroprotective effects in retinal ganglion cells. Vitr. Cell. Dev. Biol. Anim. 2016, 53, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Mead, B.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Paracrine-mediated neuroprotection and neuritogenesis of axotomised retinal ganglion cells by human dental pulp stem cells: Comparison with human bone marrow and adipose-derived mesenchymal stem cells. PLoS ONE 2014, 9, e109305. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.C.; Yeo, R.W.Y.; Lim, S.K. Mesenchymal stem cell exosomes. Semin. Cell Dev. Biol. 2015, 40, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Burrello, J.; Monticone, S.; Gai, C.; Gomez, Y.; Kholia, S.; Camussi, G. Stem cell-derived extracellular vesicles and immune-modulation. Front. Cell Dev. Biol. 2016, 4, 83. [Google Scholar] [CrossRef] [PubMed]
- Wyse, R.D.; Dunbar, G.L.; Rossignol, J. Use of genetically modified mesenchymal stem cells to treat neurodegenerative diseases. Int. J. Mol. Sci. 2014, 15, 1719–1745. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Shao, H.; Su, C.; Jiang, Y.; Chen, X.; Bai, L.; Zhang, Y.; Li, Q.; Zhang, X.; Li, X. Exosomes derived from MSCs ameliorate retinal laser injury partially by inhibition of MCP-1. Sci. Rep. 2016, 6, 34562. [Google Scholar] [CrossRef] [PubMed]
- Nussenblatt, R.B.; Lee, R.W.J.; Chew, E.; Wei, L.; Liu, B.; Sen, H.N.; Dick, A.D.; Ferris, F.L. Immune responses in age-related macular degeneration and a possible long-term therapeutic strategy for prevention. Am. J. Ophthalmol. 2014, 158, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.V.; Xu, H. Good news-bad news: The yin and yang of immune privilege in the eye. Front. Immunol. 2012, 3, 338. [Google Scholar] [CrossRef] [PubMed]
- Perez, V.L.; Caspi, R.R. Immune mechanisms in inflammatory and degenerative eye disease. Trends Immunol. 2015, 36, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, I.; Van Noorden, C.J.F.; Schlingemann, R.O. Molecular basis of the inner blood-retinal barrier and its breakdown in diabetic macular edema and other pathological conditions. Prog. Retin. Eye Res. 2013, 34, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.R. A look at autoimmunity and inflammation in the eye. J. Clin. Investig. 2010, 120, 3073–3083. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, S.M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.-L.; Tse, H.F.; Fu, Q.L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, G.K.; Kondziolka, D.; Wechsler, L.R.; Lunsford, L.D.; Coburn, M.L.; Billigen, J.B.; Kim, A.S.; Johnson, J.N.; Bates, D.; King, B.; et al. Clinical outcomes of transplanted modified bone marrow-derived mesenchymal stem cells in stroke: A phase 1/2a study. Stroke 2016, 47, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Ko, A.Y.; Ko, J.H.; Lee, H.J.; Kim, M.K.; Wee, W.R.; Khwarg, S.I.; Oh, J.Y. Mesenchymal stem/stromal cells protect the ocular surface by suppressing inflammation in an experimental dry eye. Mol. Ther. 2015, 23, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Tasso, R.; Ilengo, C.; Quarto, R.; Cancedda, R.; Caspi, R.R.; Pennesi, G. Mesenchymal stem cells induce functionally active T-regulatory lymphocytes in a paracrine fashion and ameliorate experimental autoimmune uveitis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, H.; Shao, H.; Nian, H.; Zhang, Y.; Bai, L.; Su, C.; Liu, X.; Dong, L.; Li, X.; et al. Long-term therapeutic effects of mesenchymal stem cells compared to dexamethasone on recurrent experimental autoimmune uveitis of rats. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5561–5571. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Jiao, C.; Zhao, S.; Li, X.; Ren, X.; Zhang, L.; Han, Z.C.; Zhang, X. Immunomodulatory effects of mesenchymal stem cells in a rat corneal allograft rejection model. Exp. Eye Res. 2012, 102, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.M.; Crawley, L.; Pahlitzsch, M.; Javaid, F.; Cordeiro, M.F. Glaucoma: The retina and beyond. Acta Neuropathol. 2016, 132, 807–826. [Google Scholar] [CrossRef] [PubMed]
- Munemasa, Y.; Kitaoka, Y. Molecular mechanisms of retinal ganglion cell degeneration in glaucoma and future prospects for cell body and axonal protection. Front. Cell. Neurosci. 2012, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Emre, E.; Yüksel, N.; Duruksu, G.; Pirhan, D.; Subaşi, C.; Erman, G.; Karaöz, E. Neuroprotective effects of intravitreally transplanted adipose tissue and bone marrow-derived mesenchymal stem cells in an experimental ocular hypertension model. Cytotherapy 2015, 17, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Hou, H.; Liang, H.; Weinreb, R.N.; Wang, H.; Wang, Y. Bone marrow-derived cells in ocular neovascularization: Contribution and mechanisms. Angiogenesis 2016, 19, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Diabetic macular edema, retinopathy and age-related macular degeneration as inflammatory conditions. Arch. Med. Sci. 2016, 12, 1142–1157. [Google Scholar] [CrossRef] [PubMed]
- Ghazaryan, E.; Zhang, Y.; He, Y.; Liu, X.; Li, Y.; Xie, J.; Su, G. Mesenchymal stem cells in corneal neovascularization: Comparison of different application routes. Mol. Med. Rep. 2016, 14, 3104–3112. [Google Scholar] [CrossRef] [PubMed]
- Yafai, Y.; Eichler, W.; Iandiev, I.; Unterlauft, J.-D.; Jochmann, C.; Wiedemann, P.; Bringmann, A. Thrombospondin-1 is produced by retinal glial cells and inhibits the growth of vascular endothelial cells. Ophthalmic Res. 2014, 52, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Farnoodian, M.; Kinter, J.B.; Yadranji Aghdam, S.; Zaitoun, I.; Sorenson, C.M.; Sheibani, N. Expression of pigment epithelium-derived factor and thrombospondin-1 regulate proliferation and migration of retinal pigment epithelial cells. Physiol. Rep. 2015, 3, e12266. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Ruiz, L.; Regenfuss, B.; Mir, F.A.; Kearns, J.; Masli, S. Conjunctival inflammation in thrombospondin-1 deficient mouse model of Sjögren’s syndrome. PLoS ONE 2013, 8, e75937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, H.-Y.; Liang, H.-L.; Wang, Y.-S.; Zhang, Z.-X.; Wang, B.-R.; Shi, Y.-Y.; Dong, X.; Cai, Y. A therapeutic strategy for choroidal neovascularization based on recruitment of mesenchymal stem cells to the sites of lesions. Mol. Ther. 2010, 18, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Ribot, J.; Caliaperoumal, G.; Paquet, J.; Boisson-Vidal, C.; Petite, H.; Anagnostou, F. Type 2 diabetes alters mesenchymal stem cell secretome composition and angiogenic properties. J. Cell. Mol. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.Y.; Ramakrishnan, D.P.; Silverstein, R.L. Thrombospondin-1 modulates VEGF signaling via CD36 by recruiting SHP-1 to VEGFR2 complex in microvascular endothelial cells. Blood 2013, 122, 1822–1832. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Target | Source of MSCs | Experimental Design (Route of Delivery; Cell Concentration) | Research Outcomes | References |
|---|---|---|---|---|
| AMD | Rat bone marrow | Subretinal; 1.0 × 106 cells/eye | Integrated MSCs enhanced retinal cell survivability, architecture, and functionality of induced retinal degeneration rat model through MSC differentiation and replacement of loss RPE. | [61] |
| Diabetic retinopathy | Rat bone marrow | Intravitreal; 1.0 × 105 cells/µL | MSCs were found to integrate mostly in the diabetic eyes with reduction in retinal gliosis followed by an increased in retinal function. | [62] |
| Mouse adipose tissue | Intravitreal; 1.0 × 105 cells/µL | MSCs regenerated into retinal astrocytes and RGC, and protected RGC from oxidative damage through secretion of MSC factors comprising of NGF, bFGF, and GDNF. | [51] | |
| Retinal ischemia | Human bone marrow | Intravitreal; Not available | Administration of hypoxic-conditioned medium from MSCs in rat model of retinal ischemia promoted RGC survivability and restored retinal function through MSC paracrine effect. | [50] |
| Not available | Intraocular; 1.0 × 104 cells/µL | Engrafted MSCs improved RGC survivability after retinal ischemic injury in a mouse model. | [63] | |
| Retinal degeneration | Human bone marrow | Subretinal and intravitreal; 5.0 × 105 cells/µL | MSCs increased photoreceptor cell survivability from degeneration and sustained retinal function in retinal degenerating rat model. | [64] |
| Epiretinally; 5 × 104 cells/µL | [65] | |||
| Subretinal; 2.5 × 104 cells/µL | [66] | |||
| Human umbilical cord blood | Subretinal; Not available | A significant preservation of degenerating rat retinal photoreceptors, function, and architecture through secretion of MSC neurotrophic factors, such as IL-6, FGF2, and BDNF. | [67] | |
| Glaucoma | Rat bone marrow | In vitro co-culture system; Not available | In vitro co-cultured of MSCs with hypoxic-induced rat RGC exerted anti-apoptotic effect on RGC via reduction in caspase-3 activity. | [68] |
| Human dental pulp, bone marrow, and adipose tissue | Intravitreal; 3.0 × 104 cells/µL | MSCs derived from human dental pulp and bone marrow increased RGC survivability and restored retinal function in ocular-induced hypertensive rat model. | [53] |
| Application | Source of MSCs | Experimental Design | Clinical Phases | Clinical Trials Identifier (ClinicalTrials.gov) | |
|---|---|---|---|---|---|
| Route of Delivery | Concentration of Stem cells | ||||
| Umbilical tissue | Subretinal | 6.0 × 104 cells–3.0 × 105 cells | Phase 1/2a | NCT01226628 |
| Bone marrow | Intravitreal | Not available | Phase 1/2 | NCT02016508 | |
| Bone marrow | Intravitreal | 3.4 × 106 cells/0.1 mL | Phase 1 | NCT01736059 | |
| Bone marrow | Intravitreal | 10.0 × 106 cells/0.1 mL | Phase 1/2 | NCT01518127 | |
| Bone marrow | Intravitreal | 3.4 × 106 cells/0.1 mL | Phase 1 | NCT01736059 |
| Bone marrow | Intravitreal | 1.0 × 106 cells/0.1 mL | Phase 1 | NCT01531348 | |
| Bone marrow | Intravitreal | 10.0 × 106 cells/0.1 mL | Phase 2 | NCT01560715 | |
| Bone marrow | Intravitreal | 3.4 × 106 cells/0.1 mL | Phase 1 | NCT01736059 | |
| Bone marrow | Intravitreal | 10.0 × 106 cells/0.1 mL | Phase 1 | NCT01068561 | |
| Bone marrow | Intravitreal | 3.4 × 106 cells/0.1 mL | Phase 1 | NCT01736059 |
| Bone marrow | Intravitreal | 10.0 × 106 cells/0.1 mL | Phase 1/2 | NCT01518842 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, S.L.S.; Kumar, S.; Mok, P.L. Cellular Reparative Mechanisms of Mesenchymal Stem Cells for Retinal Diseases. Int. J. Mol. Sci. 2017, 18, 1406. https://doi.org/10.3390/ijms18081406
Ding SLS, Kumar S, Mok PL. Cellular Reparative Mechanisms of Mesenchymal Stem Cells for Retinal Diseases. International Journal of Molecular Sciences. 2017; 18(8):1406. https://doi.org/10.3390/ijms18081406
Chicago/Turabian StyleDing, Suet Lee Shirley, Suresh Kumar, and Pooi Ling Mok. 2017. "Cellular Reparative Mechanisms of Mesenchymal Stem Cells for Retinal Diseases" International Journal of Molecular Sciences 18, no. 8: 1406. https://doi.org/10.3390/ijms18081406
APA StyleDing, S. L. S., Kumar, S., & Mok, P. L. (2017). Cellular Reparative Mechanisms of Mesenchymal Stem Cells for Retinal Diseases. International Journal of Molecular Sciences, 18(8), 1406. https://doi.org/10.3390/ijms18081406