TGFβ Controls Ovarian Cancer Cell Proliferation

 , , ,
, , , 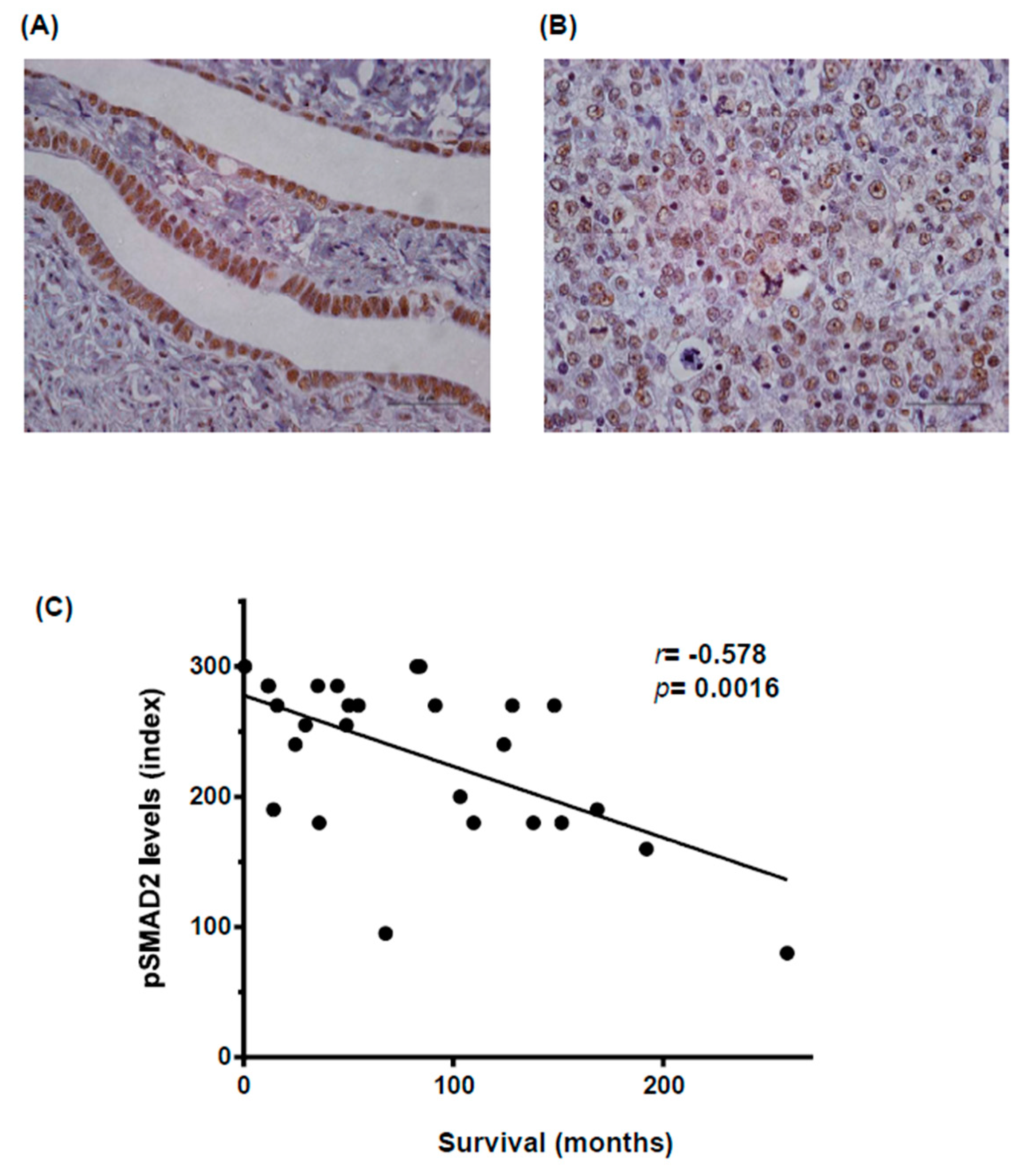{kind=link}
Abstract
:1. Ovarian Cancer
2. The Highly Discrepant Literature about the Origin of Ovarian Cancer
3. TGFβ Transforming Growth Factor) Member Signaling Occurs in Normal Ovary
4. TGF-β and Ovarian Cancer
5. TGFβ Controls Proliferation of Ovarian Cancer Cells
6. Therapeutic Approaches
7. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berrino, F.; De Angelis, R.; Sant, M.; Rosso, S.; Bielska-Lasota, M.; Coebergh, J.W.; Santaquilani, M. Survival for eight major cancers and all cancers combined for European adults diagnosed in 1995–99: Results of the eurocare-4 study. Lancet Oncol. 2007, 8, 773–783. [Google Scholar] [CrossRef]
- Ferlay, J.; Parkin, D.M.; Steliarova-Foucher, E. Estimates of cancer incidence and mortality in Europe in 2008. Eur. J. Cancer 2010, 46, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Oberaigner, W.; Minicozzi, P.; Bielska-Lasota, M.; Allemani, C.; de Angelis, R.; Mangone, L.; Sant, M. Survival for ovarian cancer in Europe: The across-country variation did not shrink in the past decade. Acta. Oncol. 2012, 51, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, E.; Meeker, A.K.; Visvanathan, K.; Gross, A.L.; Wang, T.L.; Kurman, R.J.; Shih Ie, M. Telomere length in different histologic types of ovarian carcinoma with emphasis on clear cell carcinoma. Mod. Pathol. 2011, 24, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.W.; Kleihues, P. World Cancer Report; IARC Press: Lyon, France, 2003. [Google Scholar]
- Dubeau, L. The cell of origin of ovarian epithelial tumours. Lancet. Oncol. 2008, 9, 1191–1197. [Google Scholar] [CrossRef]
- Prat, J. Ovarian carcinomas: Five distinct diseases with different origins, genetic alterations, and clinicopathological features. Virchows. Arch. 2012, 460, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Shih Ie, M. The origin and pathogenesis of epithelial ovarian cancer: A proposed unifying theory. Am. J. Surg. Pathol. 2010, 34, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Karnezis, A.N.; Cho, K.R.; Gilks, C.B.; Pearce, C.L.; Huntsman, D.G. The disparate origins of ovarian cancers: Pathogenesis and prevention strategies. Nat. Rev. Cancer 2017, 17, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, F.; Muto, M.G.; Lee, Y.; Elvin, J.A.; Callahan, M.J.; Feltmate, C.; Garber, J.E.; Cramer, D.W.; Crum, C.P. The tubal fimbria is a preferred site for early adenocarcinoma in women with familial ovarian cancer syndrome. Am. J. Surg. Pathol. 2006, 30, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Miron, A.; Drapkin, R.; Nucci, M.R.; Medeiros, F.; Saleemuddin, A.; Garber, J.; Birch, C.; Mou, H.; Gordon, R.W.; et al. A candidate precursor to serous carcinoma that originates in the distal fallopian tube. J. Pathol. 2007, 211, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Karst, A.M.; Levanon, K.; Drapkin, R. Modeling high-grade serous ovarian carcinogenesis from the fallopian tube. Proc. Natl. Acad. Sci. USA 2011, 108, 7547–7552. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Shih, Ie.M. Molecular pathogenesis and extraovarian origin of epithelial ovarian cancer–shifting the paradigm. Hum. Pathol. 2011, 42, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Terada, T. Endometrioid adenocarcinoma of the ovary arising in atypical endometriosis. Int. J. Clin. Exp. Pathol. 2012, 5, 924–927. [Google Scholar] [PubMed]
- Yuan Z, W.Y.; Cragun, J.M.; Chambers, S.K.; Zheng, W. Cell origin of endometriosis: Contribution by the fallopian tube epithelium. Am. J. Clin. Exp. Obstet. Gynecol. 2013, 1, 37–42. [Google Scholar]
- Eckert, M.A.; Pan, S.; Hernandez, K.M.; Loth, R.M.; Andrade, J.; Volchenboum, S.L.; Faber, P.; Montag, A.; Lastra, R.; Peter, M.E.; et al. Genomics of ovarian cancer progression reveals diverse metastatic trajectories including intraepithelial metastasis to the fallopian tube. Cancer Discov. 2016, 6, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Bast, R.C., Jr.; Hennessy, B.; Mills, G.B. The biology of ovarian cancer: New opportunities for translation. Nat. Rev. Cancer 2009, 9, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, S.; Coward, J.I.; Bast, R.C., Jr.; Berchuck, A.; Berek, J.S.; Brenton, J.D.; Coukos, G.; Crum, C.C.; Drapkin, R.; Etemadmoghadam, D.; et al. Rethinking ovarian cancer: Recommendations for improving outcomes. Nat. Rev. Cancer 2011, 11, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Chandler, R.L.; Damrauer, J.S.; Raab, J.R.; Schisler, J.C.; Wilkerson, M.D.; Didion, J.P.; Starmer, J.; Serber, D.; Yee, D.; Xiong, J.; et al. Coexistent ARID1A-PIK3CA mutations promote ovarian clear-cell tumorigenesis through pro-tumorigenic inflammatory cytokine signalling. Nat. Commun. 2015, 6, 6118. [Google Scholar] [CrossRef] [PubMed]
- Moxley, K.M.; Wang, L.; Welm, A.L.; Bieniasz, M. Short-form Ron is a novel determinant of ovarian cancer initiation and progression. Genes Cancer 2016, 7, 169–181. [Google Scholar] [PubMed]
- Knight, P.G.; Glister, C. TGF-β superfamily members and ovarian follicle development. Reproduction 2006, 132, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Reader, K.L.; Gold, E. Activins and activin antagonists in the human ovary and ovarian cancer. Mol. Cell Endocrinol. 2015, 415, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Qiao, J.; Leung, P.C.K. Oocyte–somatic cell interactions in the human ovary—Novel role of bone morphogenetic proteins and growth differentiation factors. Hum. Reprod. Update 2017, 23, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.; Doraiswamy, V.; Parrott, J.A.; Skinner, M.K. Expression and action of transforming growth factor β (TGFβ1, TGFβ2, TGFβ3) in normal bovine ovarian surface epithelium and implications for human ovarian cancer. Mol. Cell Endocrinol. 2001, 182, 145–155. [Google Scholar] [CrossRef]
- Alsina-Sanchis, E.; Figueras, A.; Lahiguera, A.; Vidal, A.; Casanovas, O.; Graupera, M.; Villanueva, A.; Viñals, F. The TGFβ pathway stimulates ovarian cancer cell proliferation by increasing igf1r levels. Int. J. Cancer 2016, 139, 1894–1903. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Pangas, S.A.; Jorgez, C.J.; Graff, J.M.; Weinstein, M.; Matzuk, M.M. Redundant roles of SMAD2 and SMAD3 in ovarian granulosa cells in vivo. Mol. Cell Biol. 2008, 28, 7001–7011. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ren, Y.A.; Pangas, S.A.; Adams, J.; Zhou, W.; Castrillon, D.H.; Wilhelm, D.; Richards, J.S. Foxo1/3 and PTEN depletion in granulosa cells promotes ovarian granulosa cell tumor development. Mol. Endocrinol. 2015, 29, 1006–1024. [Google Scholar] [CrossRef] [PubMed]
- Pangas, S.A.; Li, X.; Umans, L.; Zwijsen, A.; Huylebroeck, D.; Gutierrez, C.; Wang, D.; Martin, J.F.; Jamin, S.P.; Behringer, R.R.; et al. Conditional deletion of SMAD1 and SMAD5 in somatic cells of male and female gonads leads to metastatic tumor development in mice. Mol. Cell Biol. 2008, 28, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Neil, J.R.; Schiemann, W.P. Transforming growth factor-β and the hallmarks of cancer. Cell. Signal. 2011, 23, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Helleman, J.; Jansen, M.P.; Burger, C.; van der Burg, M.E.; Berns, E.M. Integrated genomics of chemotherapy resistant ovarian cancer: A role for extracellular matrix, TGFβ and regulating micrornas. Int. J. Biochem. Cell Biol. 2010, 42, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Antony, M.L.; Nair, R.; Sebastian, P.; Karunagaran, D. Changes in expression, and/or mutations in TGF-β receptors (TGF-β RI and TGF-β RII) and SMAD 4 in human ovarian tumors. J. Cancer Res. Clin. Oncol. 2010, 136, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Kaklamani, V.G.; Hou, N.; Bian, Y.; Reich, J.; Offit, K.; Michel, L.S.; Rubinstein, W.S.; Rademaker, A.; Pasche, B. TGFBR1*6A and cancer risk: A meta-analysis of seven case-control studies. J. Clin. Oncol. 2003, 21, 3236–3243. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.; Lee, C.; Joseph, P.; Marchini, S.; Baccarini, A.; Kolev, V.; Romualdi, C.; Fruscio, R.; Shah, H.; Wang, F.; et al. Microrna-181A has a critical role in ovarian cancer progression through the regulation of the epithelial-mesenchymal transition. Nat. Commun. 2014, 5, 2977. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, M.; Zannoni, G.F.; Beltrame, L.; Martinelli, E.; DiFeo, A.; Paracchini, L.; Craparotta, I.; Mannarino, L.; Vizzielli, G.; Scambia, G.; et al. Identification of high-grade serous ovarian cancer mirna species associated with survival and drug response in patients receiving neoadjuvant chemotherapy: A retrospective longitudinal analysis using matched tumor biopsies. Ann. Oncol. 2016, 27, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, R.; Gobl, A.; Wilander, E.; Oberg, K.; Miyazono, K.; Funa, K. Expression and prognostic significance of TGF-β isotypes, latent TGF-β 1 binding protein, TGF-β type I and type II receptors, and endoglin in normal ovary and ovarian neoplasms. Lab. Investig. 1995, 73, 213–220. [Google Scholar] [PubMed]
- Rodon, L.; Gonzalez-Junca, A.; Inda Mdel, M.; Sala-Hojman, A.; Martinez-Saez, E.; Seoane, J. Active creb1 promotes a malignant TGFβ2 autocrine loop in glioblastoma. Cancer Discov. 2014, 4, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Gordinier, M.E.; Zhang, H.Z.; Patenia, R.; Levy, L.B.; Atkinson, E.N.; Nash, M.A.; Katz, R.L.; Platsoucas, C.D.; Freedman, R.S. Quantitative analysis of transforming growth factor β 1 and 2 in ovarian carcinoma. Clin. Cancer Res. 1999, 5, 2498–2505. [Google Scholar] [PubMed]
- Do, T.V.; Kubba, L.A.; Du, H.; Sturgis, C.D.; Woodruff, T.K. Transforming growth factor-β1, transforming growth factor-β2, and transforming growth factor-β3 enhance ovarian cancer metastatic potential by inducing a SMAD3-dependent epithelial-to-mesenchymal transition. Mol. Cancer Res. 2008, 6, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhu, Y.; Nilsson, M.; Sundfeldt, K. TGF-β isoforms induce EMT independent migration of ovarian cancer cells. Cancer Cell Int. 2014, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Komiyama, S.; Kurahashi, T.; Ishikawa, M.; Tanaka, K.; Komiyama, M.; Mikami, M.; Udagawa, Y. Expression of TGFSS1 and its receptors is associated with biological features of ovarian cancer and sensitivity to paclitaxel/carboplatin. Oncol. Rep. 2011, 25, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.I.; Jussila, T.; Fusenig, N.E.; Stenback, F. TGF-β isoforms are differentially expressed in increasing malignant grades of HaCaT keratinocytes, suggesting separate roles in skin carcinogenesis. J. Pathol. 2000, 190, 579–588. [Google Scholar] [CrossRef]
- Baldwin, R.L.; Tran, H.; Karlan, B.Y. Loss of c-Myc repression coincides with ovarian cancer resistance to transforming growth factor β growth arrest independent of transforming growth factor β/SMAD signaling. Cancer Res. 2003, 63, 1413–1419. [Google Scholar] [PubMed]
- Jozan, S.; Guerrin, M.; Mazars, P.; Dutaur, M.; Monsarrat, B.; Cheutin, F.; Bugat, R.; Martel, P.; Valette, A. Transforming growth factor β 1 (TGF-β 1) inhibits growth of a human ovarian carcinoma cell line (ovccr1) and is expressed in human ovarian tumors. Int. J. Cancer 1992, 52, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Kohan-Ivani, K.; Gabler, F.; Selman, A.; Vega, M.; Romero, C. Role of dihydrotestosterone (dht) on TGF-β1 signaling pathway in epithelial ovarian cancer cells. J. Cancer Res. Clin. Oncol. 2016, 142, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Bruchim, I.; Werner, H. Targeting IGF-1 signaling pathways in gynecologic malignancies. Expert. Opin. Ther. Targets 2013, 17, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Bruna, A.; Darken, R.S.; Rojo, F.; Ocana, A.; Penuelas, S.; Arias, A.; Paris, R.; Tortosa, A.; Mora, J.; Baselga, J.; et al. High TGF β-SMAD activity confers poor prognosis in glioma patients and promotes cell proliferation depending on the methylation of the pdgf-b gene. Cancer Cell. 2007, 11, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Dunfield, L.D.; Nachtigal, M.W. Inhibition of the antiproliferative effect of TGFβ by EGF in primary human ovarian cancer cells. Oncogene 2003, 22, 4745–4751. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Shi, H.; Xie, Y.; Zhao, Z.; Li, S.; Chang, C.; Cheng, X.; Li, Y. Ubiquitin specific protease 22 promotes cell proliferation and tumor growth of epithelial ovarian cancer through synergy with transforming growth factor β1. Oncol. Rep. 2015, 33, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn, P.J.; Rodon, L.; Gonzalez-Junca, A.; Dirac, A.; Gili, M.; Martinez-Saez, E.; Aura, C.; Barba, I.; Peg, V.; Prat, A.; et al. Usp15 stabilizes TGF-β receptor I and promotes oncogenesis through the activation of TGF-β signaling in glioblastoma. Nat. Med. 2012, 18, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Al-Salihi, M.A.; Herhaus, L.; Macartney, T.; Sapkota, G.P. Usp11 augments TGF β signalling by deubiquitylating Alk5. Open Biol. 2012, 2, 120063. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, F.; Drabsch, Y.; Gao, R.; Snaar-Jagalska, B.E.; Mickanin, C.; Huang, H.; Sheppard, K.A.; Porter, J.A.; Lu, C.X.; et al. Usp4 is regulated by Akt phosphorylation and directly deubiquitylates TGF-β type I receptor. Nat. Cell. Biol. 2012, 14, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef] [PubMed]
- Foley, O.W.; Rauh-Hain, J.A.; del Carmen, M.G. Recurrent epithelial ovarian cancer: An update on treatment. Oncology (Williston Park) 2013, 27, 288–294. [Google Scholar] [PubMed]
- Liao, S.; Liu, J.; Lin, P.; Shi, T.; Jain, R.K.; Xu, L. TGF-β blockade controls ascites by preventing abnormalization of lymphatic vessels in orthotopic human ovarian carcinoma models. Clin. Cancer Res. 2011, 17, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Shan, N.; Zhao, C.; Wang, Y.; Xu, F.; Li, J.; Yu, X.; Gao, L.; Yi, Z. Ly2109761 enhances cisplatin antitumor activity in ovarian cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 4923–4932. [Google Scholar] [PubMed]
- Neuzillet, C.; Tijeras-Raballand, A.; Cohen, R.; Cros, J.; Faivre, S.; Raymond, E.; de Gramont, A. Targeting the TGFβ pathway for cancer therapy. Pharmacol. Ther. 2015, 147, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Dituri, F.; Mazzocca, A.; Fernando, J.; Papappicco, P.; Fabregat, I.; De Santis, F.; Paradiso, A.; Sabbà, C.; Giannelli, G. Differential inhibition of the TGF-β signaling pathway in hcc cells using the small molecule inhibitor LY2157299 and the D10 monoclonal antibody against TGF-β receptor type II. PLoS ONE 2013, 8, e67109. [Google Scholar] [CrossRef]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsina-Sanchís, E.; Figueras, A.; Lahiguera, A.; Gil-Martín, M.; Pardo, B.; Piulats, J.M.; Martí, L.; Ponce, J.; Matias-Guiu, X.; Vidal, A.; et al. TGFβ Controls Ovarian Cancer Cell Proliferation. Int. J. Mol. Sci. 2017, 18, 1658. https://doi.org/10.3390/ijms18081658
Alsina-Sanchís E, Figueras A, Lahiguera A, Gil-Martín M, Pardo B, Piulats JM, Martí L, Ponce J, Matias-Guiu X, Vidal A, et al. TGFβ Controls Ovarian Cancer Cell Proliferation. International Journal of Molecular Sciences. 2017; 18(8):1658. https://doi.org/10.3390/ijms18081658
Chicago/Turabian StyleAlsina-Sanchís, Elisenda, Agnès Figueras, Alvaro Lahiguera, Marta Gil-Martín, Beatriz Pardo, Josep M. Piulats, Lola Martí, Jordi Ponce, Xavier Matias-Guiu, August Vidal, and et al. 2017. "TGFβ Controls Ovarian Cancer Cell Proliferation" International Journal of Molecular Sciences 18, no. 8: 1658. https://doi.org/10.3390/ijms18081658
APA StyleAlsina-Sanchís, E., Figueras, A., Lahiguera, A., Gil-Martín, M., Pardo, B., Piulats, J. M., Martí, L., Ponce, J., Matias-Guiu, X., Vidal, A., Villanueva, A., & Viñals, F. (2017). TGFβ Controls Ovarian Cancer Cell Proliferation. International Journal of Molecular Sciences, 18(8), 1658. https://doi.org/10.3390/ijms18081658