iTRAQ-Based Quantitative Proteomics Analysis on Rice Anther Responding to High Temperature



 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
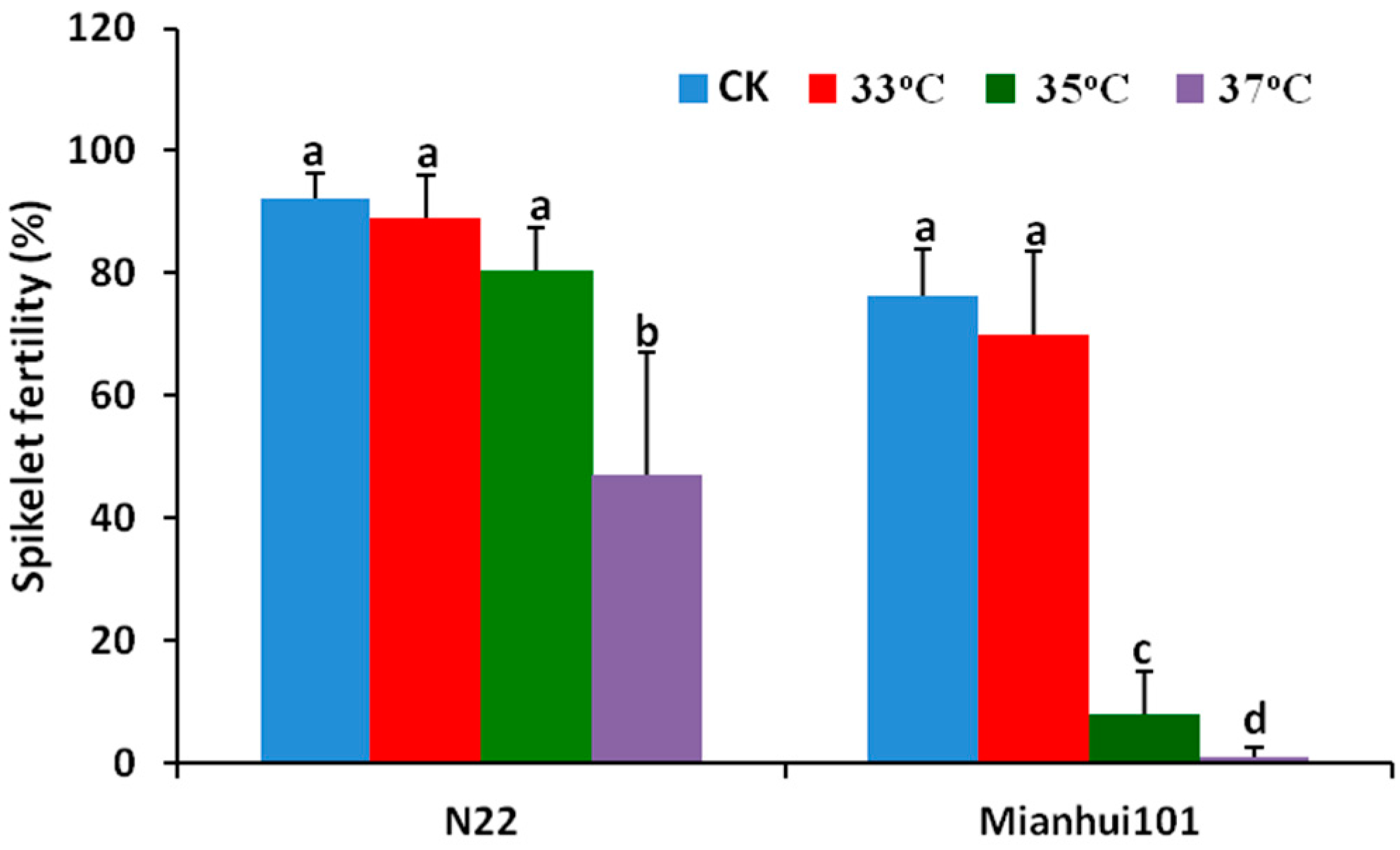
2.1. High Temperature Tolerance of N22 and Mianhui101
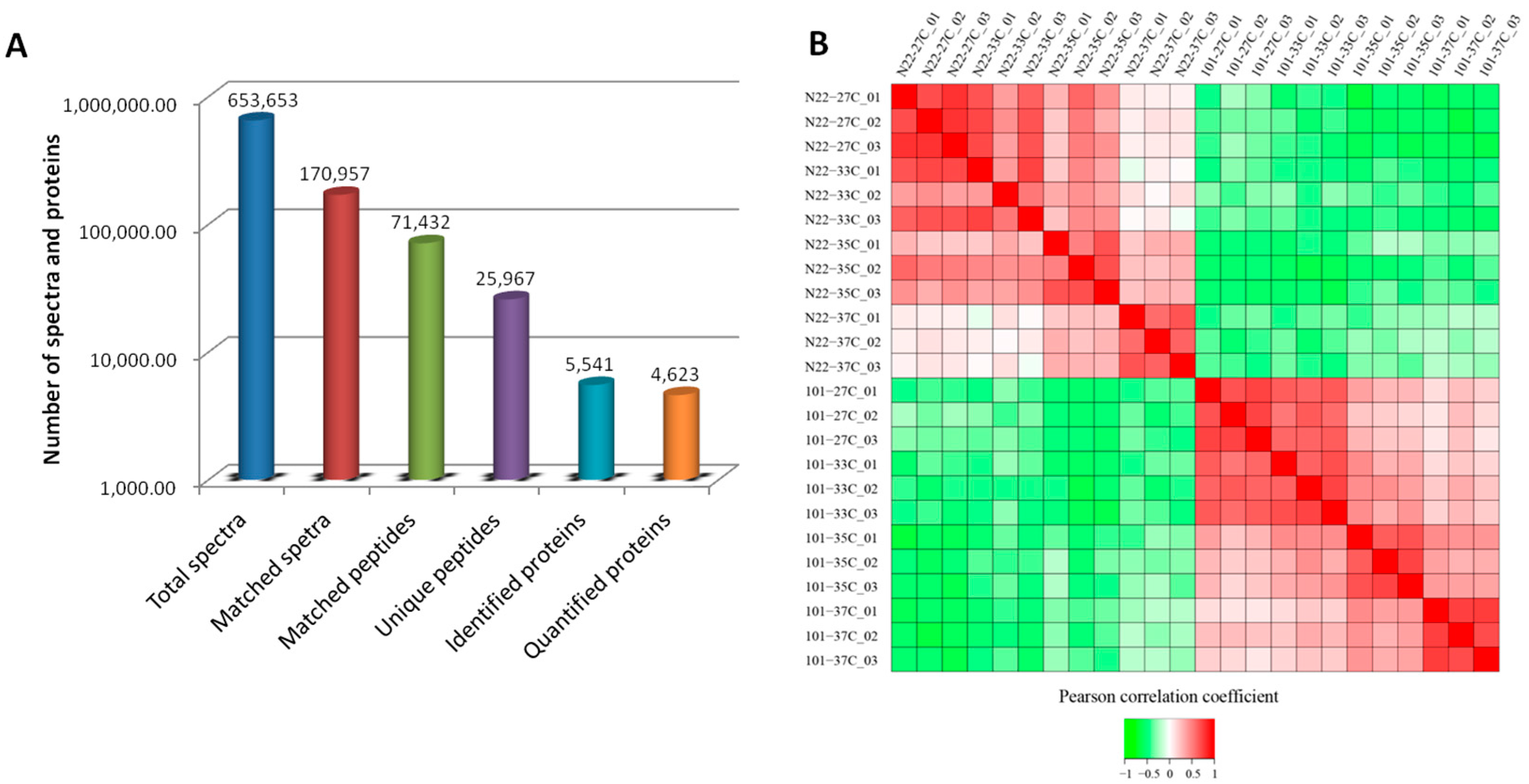
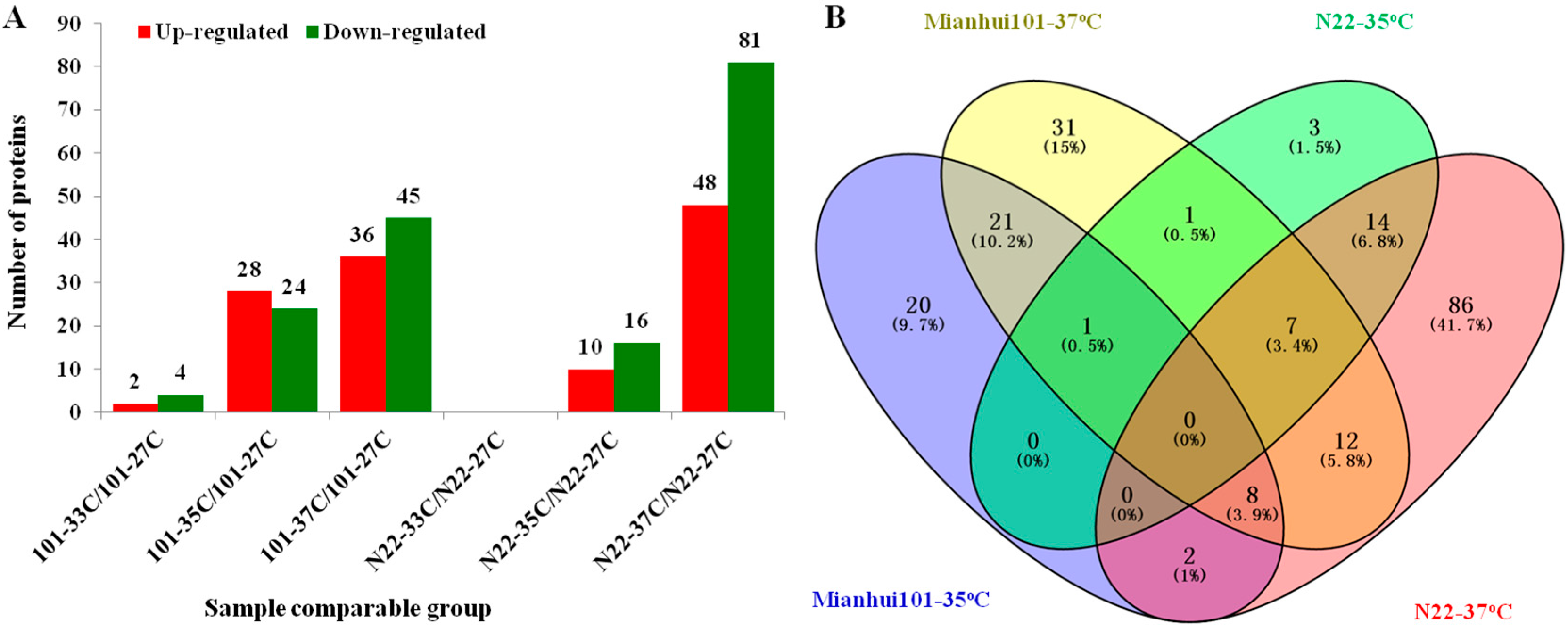
2.2. High Temperature Induced Proteome Changes in N22 and Mianhui101
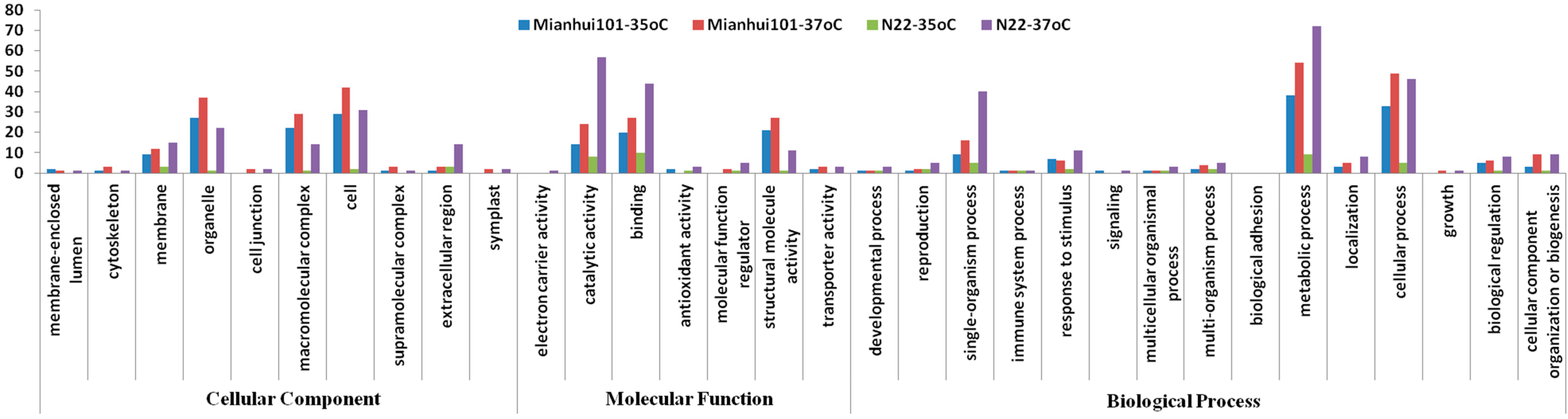
2.3. Functional Categorization and Gene Ontology (GO) Analysis of the Changed Proteins
2.4. N22 and Mianhui101 Specifically High Temperature Stress (HS) Responsive Proteins
3. Materials and Methods
3.1. Rice Materials and Growth
3.2. High Temperature Treatment and Sampling
3.3. Protein Extraction, Digestion and iTRAQ Labeling
3.4. Quantitative Proteomic Analysis by LC-MS/MS and Protein Identification by Database Searching
3.5. Bioinformatics Methods
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Chen, H.; Li, X.J.; Yang, M.F.; Liu, G.S.; Shen, S.H. A comparative proteomic analysis of rice seedlings under various high-temperature stresses. BBA Proteins Proteom. 2009, 1794, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Part A: Global and sectoral aspects. Contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change. In Climate Change 2014: Impacts, Adaptation, and Vulnerability; Field, B.C., Barros, V.R., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 1–32. [Google Scholar]
- Baker, J.T.; Allen, L.H.; Boote, K.J. Temperature effects on rice at elevated CO2 concentration. J. Exp. Bot. 1992, 43, 959–964. [Google Scholar] [CrossRef]
- Shah, F.; Huang, J.; Cui, K.; Nie, L.; Shah, T.; Chen, C.; Wang, K. Impact of high-temperature stress on rice plant and its traits related to tolerance. J. Agric. Sci. 2011, 149, 545–556. [Google Scholar] [CrossRef]
- Yoshida, S.; Satake, T.; Mackill, D.J. High Temperature Stress in Rice; IRRI Research Paper Series Number 67; International Rice Research Institute: Manila, Philippines, 1981. [Google Scholar]
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H.; Sheehy, J.E.; Thomas, J.M.G. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- Wassmann, R.; Jagadish, S.V.K.; Sumfleth, K.; Pathak, H.; Howell, G.; Ismail, A.; Heuer, S. Regional vulnerability of climate change impacts on Asian rice production and scope for adaptation. Adv. Agron. 2009, 102, 93–133. [Google Scholar]
- Sage, T.L.; Bagha, S.; Lundsgaard-Nielsen, V.; Branch, H.A.; Sultmanis, S.; Sage, R.F. The effect of high temperature stress on male and female reproduction in plants. Field Crops Res. 2015, 182, 30–42. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Craufurd, P.Q.; Wheeler, T.R. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Omasa, K.; Horie, T. High temperature at flowering inhibits swelling of pollen grains, a driving force for thecae dehiscence in rice (Oryza sativa L.). Plant Prod. Sci. 2000, 3, 430–434. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Muthurajan, R.; Oane, R.; Wheeler, T.R.; Heuer, S.; Bennett, J.; Craufurd, P.Q. Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K.; Murty, M.V.R.; Quick, W.P. Rice responses to rising temperatures—Challenges, perspectives and future directions. Plant Cell Environ. 2014, 38, 1686–1698. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, H.; Lee, W.; Lee, Y.; Kwon, S.W.; Lee, J. Quantitative shotgun proteomics analysis of rice anther proteins after exposure to high temperature. Int. J. Genom. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.P.; Deng, Z.W.; Zhou, X.L. Study of regional characteristics on mean annual temperature variation of near 117 years in China. Q. J. Appl. Meteorol. 1999, 10, 35–43. [Google Scholar]
- Yan, H.; Zhang, B.; Zhang, Y.; Chen, X.; Xiong, H.; Matsui, T.; Tian, X. High temperature induced glume closure resulted in lower fertility in hybrid rice seed production. Front. Plant Sci. 2017, 7, 1960. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K.; Craufurd, P.Q.; Wheeler, T.R. Phenotyping parents of mapping populations of rice for heat tolerance during anthesis. Crop Sci. 2008, 48, 1140–1146. [Google Scholar] [CrossRef]
- Ye, C.; Argayoso, M.A.; Redona, E.D.; Sierra, S.N.; Laza, M.A.; Dilla, C.J.; Mo, Y.; Thomson, M.J.; Chin, J.; Delaviña, C.B.; et al. Mapping QTL for heat tolerance at flowering stage in rice using SNP markers. Plant Breed. 2012, 131, 33–41. [Google Scholar] [CrossRef]
- Bahuguna, R.N.; Jha, J.; Pal, M.; Shah, D.; Lawas, L.M.; Khetarpal, S.; Jagadish, K.S. Physiological and biochemical characterization of NERICA-L-44: A novel source of heat tolerance at the vegetative and reproductive stages in rice. Physiol. Plant 2014, 154, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Endo, M.; Tsuchiya, T.; Hamada, K.; Kawamura, S.; Yano, K.; Ohshima, M.; Higashitani, A.; Watanabe, M.; Kawagishi-Kobayashi, M. High temperatures cause male sterility in rice plants with transcriptional alterations during pollen development. Plant Cell Physiol. 2009, 50, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Li, J.P.; Liu, A.L.; Zou, J.; Zhou, X.Y.; Xiang, J.H.; Rerksiri, W.; Peng, Y.; Xiong, X.; Chen, X. Expression profile in rice panicle: Insights into heat response mechanism at reproductive stage. PLoS ONE 2012, 7, e49652. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, Z.; Long, H.; Zhang, Z.; Hong, Y.; Zhang, X.; You, C.; Liang, W.; Ma, H.; Lu, P. Proteomic and phosphoproteomic analyses reveal extensive phosphorylation of regulatory proteins in developing rice anthers. Plant J. 2015, 84, 527–544. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, L.Q.; Zheng, W.Y.; Wang, R.F.; Yang, L.X. Genome-wide identification of microRNAs responsive to high temperature in rice (Oryza sativa) by high-throughput deep sequencing. J. Agron. Crop Sci. 2015, 201, 379–388. [Google Scholar] [CrossRef]
- Gonzalez-Schain, N.; Dreni, L.; Lawas, L.M.F.; Galbiati, M.; Colombo, L.; Heuer, S.; Jagadish, K.S.; Kater, M.M. Genome-wide transcriptome analysis during anthesis reveals new insights into the molecular basis of heat stress response in tolerant and sensitive rice varieties. Plant Cell Physiol. 2016, 57, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Latef, A.A.H.A.; Rasool, S.; Akram, N.A.; Ashraf, M.; Gucel, S. Role of proteomics in crop stress tolerance. Front. Plant Sci. 2016, 7, 1336. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Marowa, P.; Ding, A.; Kong, Y. Expansins: Roles in plant growth and potential applications in crop improvement. Plant Cell Rep. 2016, 35, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Guttman, M.; Li, L.C.; Cosgrove, D.J. Evolutionary divergence of β-expansin structure and function in grasses parallels emergence of distinctive primary cell wall traits. Plant J. 2015, 81, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Ge, X.; Ma, H. The rice OsDIL gene plays a role in drought tolerance at vegetative and reproductive stages. Plant Mol. Biol. 2013, 82, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Schoffl, F.; Prandl, R.; Reindl, A. Molecular responses to heat stress. In Molecular Responses to Cold, Drought, Heat and Salt Stress in Higher Plants; Shinozaki, K., Yamaguchi-Shinozaki, K., Eds.; R.G. Landes Co.: Austin, TX, USA, 1999; pp. 81–98. [Google Scholar]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Doring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Motangu, M.V.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochim. Biophys. Acta 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Mogk, A.; Schlieker, C.; Friedrich, K.L.; Schonfeld, H.J.; Vierling, E.; Bukau, B. Refolding of substrates bound to small Hsps relies on a disaggregation reaction mediated most efficiently by ClpB/DnaK. J. Biol. Chem. 2003, 278, 31033–31042. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; Scott, A.H.; Bryan, J.K.; Coleman, J.S. The methionine-rich low-molecular-weight chloroplast heat-shock protein: Evolutionary conservation and accumulation in relation to thermotolerance. Am. J. Bot. 1998, 85, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; Heckathom, S.A. The mitochondrial small heathock protein protects NADH: Ubiquinone oxidoreductase of the electron transport chain during heat stress in plants. FEBS Lett. 1998, 430, 246–250. [Google Scholar] [CrossRef]
- Luo, H.; Lee, J.Y.; Hu, Q.; Nelson-Vasilchik, K.; Eitas, T.K.; Lickwar, C.; Kausch, A.P.; Chandlee, J.M.; Hodges, T.K. RTS, a rice anther-specific gene is required for male fertility and its promoter sequence directs tissue-specific gene expression in different plant species. Plant Mol. Biol. 2006, 62, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, K.; Li, S.; Yang, P. Exploration of rice pistil responses during early post-pollination through a combined proteomic and transcriptomic analysis. J. Proteom. 2016, 131, 214–226. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Accession | Protein Description | N22-35 °C/ N22-27 °C | N22-37 °C/ N22-27 °C | Functional Category |
|---|---|---|---|---|
| Up-regulated | ||||
| A2WLG6 | Similar to 17.5 kDa class II heat shock protein | 1.58 ± 0.4 | 2.74 ± 0.72 | Stress response |
| A2X9T6 | Similar to Low molecular weight heat shock protein precursor (mitochondrial small heat shock protein 22) | 2.26 ± 0.48 | 2.93 ± 0.45 | Stress response |
| B8ARU4 | Peroxidase | 1.53 ± 0.14 | 1.9 ± 0.27 | Redox |
| Down-regulated | ||||
| A2WUN3 | Pollen Ole e 1 allergen and extensin domain containing protein | 0.46 ± 0.07 | 0.49 ± 0.05 | Growth and development |
| A2XHE1 | Aldose 1-epimerase | 0.53 ± 0.06 | 0.4 ± 0.04 | Metabolic |
| A2XNH0 | α-expansin OsEXPA7 | 0.65 ± 0.11 | 0.36 ± 0.003 | Cell wall |
| A2YNG3 | Similar to glycerophosphoryl diester phosphodiesterase | 0.45 ± 0.03 | 0.31 ± 0.04 | Metabolic |
| A2ZDK1 | Similar to β- d-xylosidase | 0.66 ± 0.02 | 0.66 ± 0.02 | Metabolic |
| A2ZFQ4 | Protease inhibitor, lipid transfer protein (LTP), Postmeiotic anther development | 0.52 ± 0.1 | 0.45 ± 0.05 | Growth and development |
| B8ARP2 | Similar to thaumatin-like protein | 0.45 ± 0.08 | 0.44 ± 0.06 | Chloroplast |
| B8B4J9 | Similar to subtilisin-like protease | 0.6 ± 0.03 | 0.49 ± 0.03 | Metabolic |
| B8BB22 | Exostosin-like family protein | 0.62 ± 0.09 | 0.63 ± 0.15 | Membrane part |
| B8BD35 | Similar to Ervatamin B (EC 3.4.22.-) (ERV-B) | 0.65 ± 0.09 | 0.57 ± 0.10 | Metabolic |
| Q40629 | Anther-specific protein RTS | 0.54 ± 0.13 | 0.53 ± 0.11 | Growth and development |
| Protein Accession | Protein Description | 101-35 °C/ 101-27 °C | 101-37 °C/ 101-27 °C | Functional Category |
|---|---|---|---|---|
| Up-regulated | ||||
| A2WMG6 | Salt stress root protein RS1 | 1.65 ± 0.24 | 1.51 ± 0.34 | Stress response |
| A2XCH7 | Tonoplast intrinsic protein (Tonoplast water channel) | 1.62 ± 0.16 | 1.58 ± 0.20 | transporter |
| A2Y1Q8 | Similar to TCH2 (TOUCH 2); calcium ion binding | 1.62 ± 0.15 | 1.78 ± 0.20 | Signaling |
| A2Y8H4 | Aldo/keto reductase domain containing protein | 1.54 ± 0.06 | 1.59 ± 0.13 | Metabolic |
| A2YCB4 | Thioredoxin fold domain containing protein | 1.57 ± 0.09 | 1.78 ± 0.19 | Redox |
| A2YI28 | Reticulon-like protein | 1.59 ± 0.26 | 2.02 ± 0.20 | Endoplasmic reticulum, ER |
| A2YM28 | Thiamine thiazole synthase, chloroplastic | 1.55 ± 0.12 | 1.8 ± 0.31 | Starch metabolic |
| A2YQ88 | Similar to PWWP domain containing protein | 1.51 ± 0.22 | 1.51 ± 0.29 | DNA binding |
| A2Z0W7 | Mannose-6-phosphate isomerase | 1.81 ± 0.07 | 1.89 ± 0.20 | Metabolic |
| B8B8G2 | Tubulin alpha chain | 1.54 ± 0.24 | 1.52 ± 0.23 | Cell structure |
| B8BF32 | HAD superfamily hydrolase-like, type 3 domain containing protein | 2.03 ± 0.59 | 3.15 ± 0.60 | Metabolic |
| Down-regulated | ||||
| A2X3J5 | 40S ribosomal protein S3a | 0.66 ± 0.17 | 0.63 ± 0.12 | Ribosome |
| A2X6N1 | 60S ribosomal protein L6 | 0.6 ± 0.02 | 0.61 ± 0.05 | Ribosome |
| A2XCC4 | Similar to 60S ribosomal protein L21 | 0.46 ± 0.07 | 0.56 ± 0.07 | Ribosome |
| A2XM46 | Similar to 60S ribosomal protein L13a-4 | 0.65 ± 0.05 | 0.62 ± 0.04 | Ribosome |
| A2YIS2 | Similar to 60S ribosomal protein L4 | 0.65 ± 0.07 | 0.66 ± 0.06 | Ribosome |
| A2YNM6 | Similar to 60S ribosomal protein L27a-3 | 0.59 ± 0.07 | 0.66 ± 0.05 | Ribosome |
| A2YVK0 | Similar to 60S ribosomal protein L34 | 0.6 ± 0.14 | 0.55 ± 0.10 | Ribosome |
| A2ZLS7 | Similar to 60S ribosomal protein L2 (fragment) | 0.61 ± 0.05 | 0.59 ± 0.05 | Ribosome |
| B8AHZ6 | 40S ribosomal protein S8 | 0.62 ± 0.03 | 0.63 ± 0.03 | Ribosome |
| B8B6Q3 | Similar to Pleiotropic drug resistance protein 3 | 0.66 ± 0.01 | 0.56 ± 0.01 | Transporter |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Q.; Zhang, W.; Zhang, Y.; Yan, H.; Liu, K.; Matsui, T.; Tian, X.; Yang, P. iTRAQ-Based Quantitative Proteomics Analysis on Rice Anther Responding to High Temperature. Int. J. Mol. Sci. 2017, 18, 1811. https://doi.org/10.3390/ijms18091811
Mu Q, Zhang W, Zhang Y, Yan H, Liu K, Matsui T, Tian X, Yang P. iTRAQ-Based Quantitative Proteomics Analysis on Rice Anther Responding to High Temperature. International Journal of Molecular Sciences. 2017; 18(9):1811. https://doi.org/10.3390/ijms18091811
Chicago/Turabian StyleMu, Qilin, Wenying Zhang, Yunbo Zhang, Haoliang Yan, Ke Liu, Tsutomu Matsui, Xiaohai Tian, and Pingfang Yang. 2017. "iTRAQ-Based Quantitative Proteomics Analysis on Rice Anther Responding to High Temperature" International Journal of Molecular Sciences 18, no. 9: 1811. https://doi.org/10.3390/ijms18091811
APA StyleMu, Q., Zhang, W., Zhang, Y., Yan, H., Liu, K., Matsui, T., Tian, X., & Yang, P. (2017). iTRAQ-Based Quantitative Proteomics Analysis on Rice Anther Responding to High Temperature. International Journal of Molecular Sciences, 18(9), 1811. https://doi.org/10.3390/ijms18091811