A Novel Workflow to Enrich and Isolate Patient-Matched EpCAMhigh and EpCAMlow/negative CTCs Enables the Comparative Characterization of the PIK3CA Status in Metastatic Breast Cancer


Abstract
:
1. Introduction
2. Results
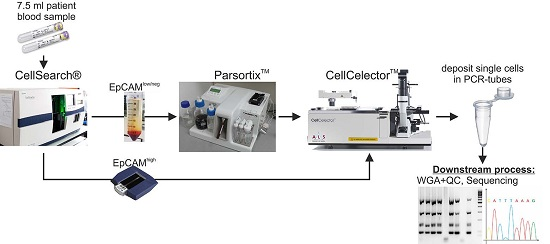
2.1. A Novel Workflow to Enrich and Isolate Patient-Matched EpCAMhigh and EpCAMlow/negative CTCs
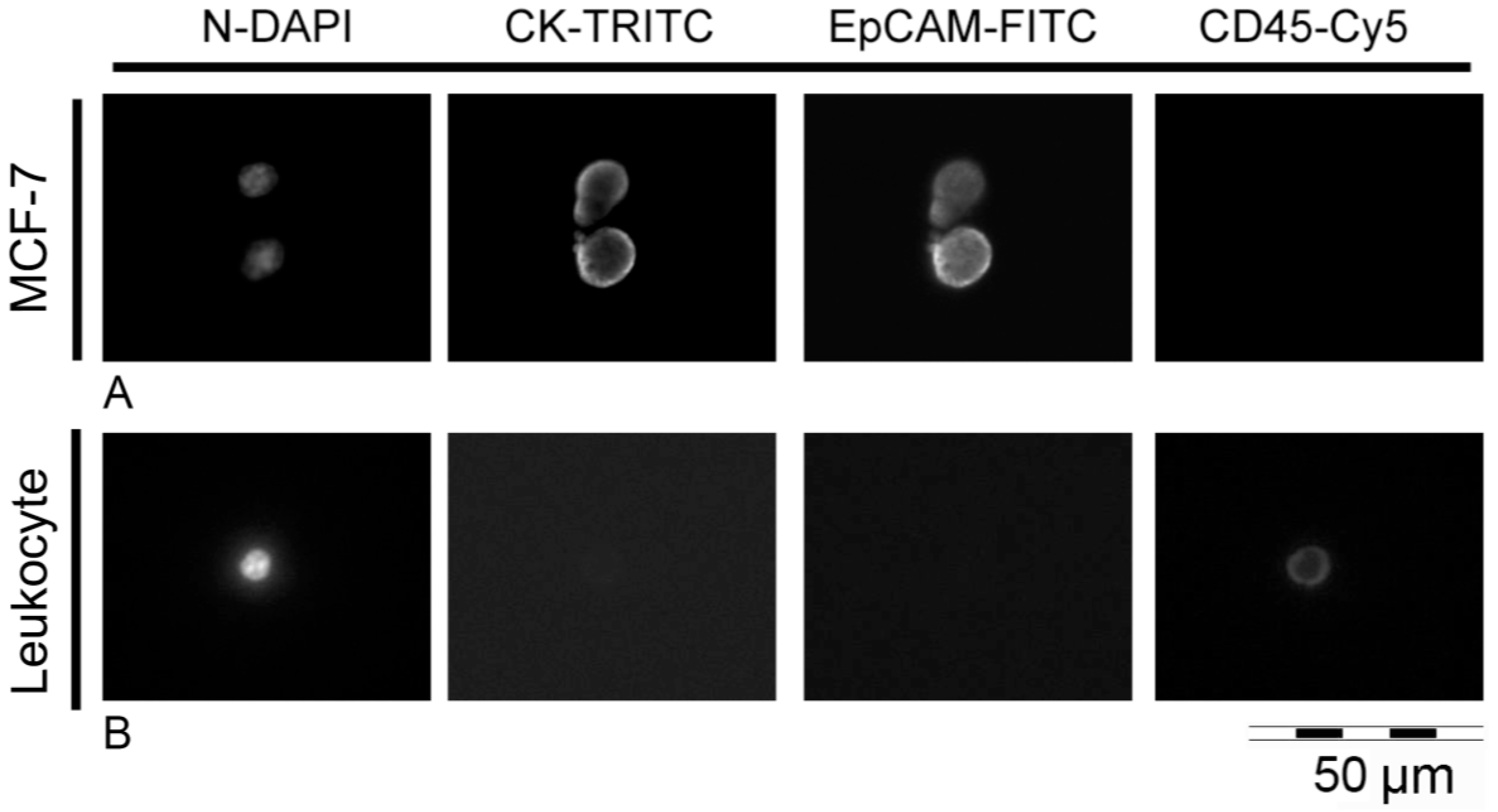
2.2. Validation of Immunostaining on Cytospins
2.3. Validation of Tumor Cell Enrichment via Parsortix™ System
2.4. Establishment of in Situ Staining of Captured Tumour Cells
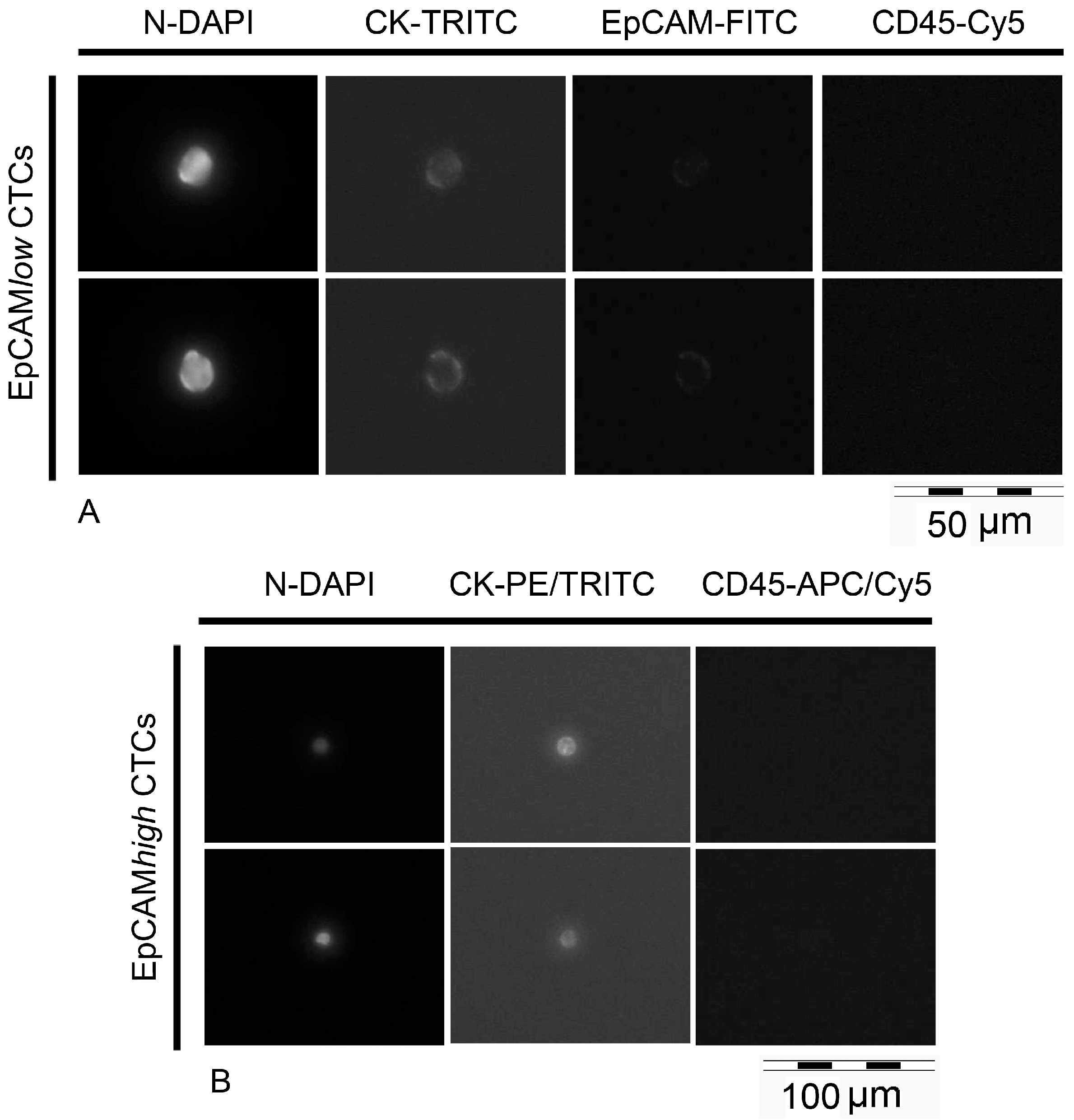
2.5. Processing of MBC Clinical Samples: Enrichment, Detection and Isolation of EpCAMhigh and EpCAMlow/negative Cells
2.6. Whole Genome Amplification of Single Isolated Cells
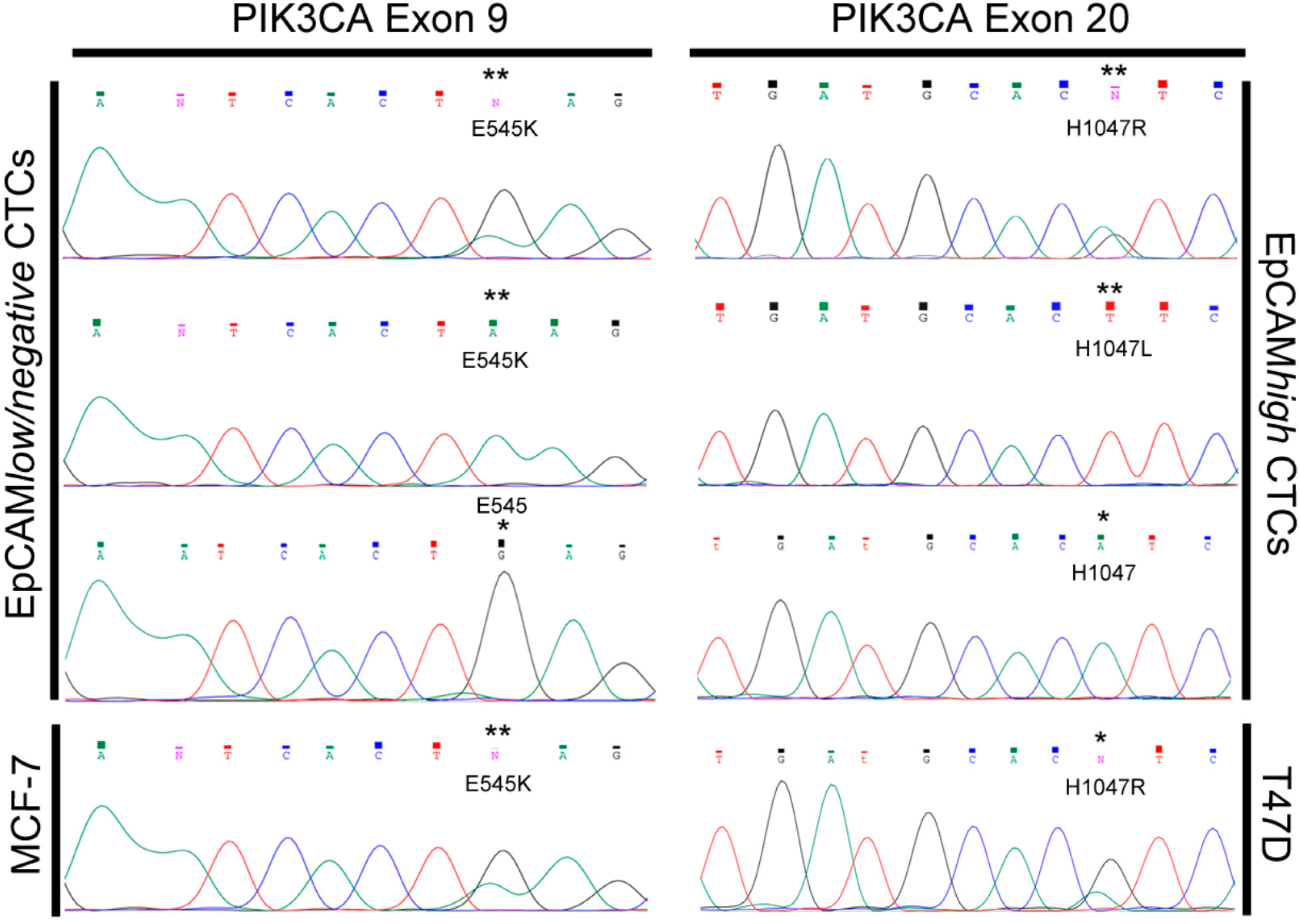
2.7. Mutational Analysis of PIK3CA Exons 9 and 20 in both CTC-Subpopulations
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. Patient Material
4.3. Enrichment and Enumeration of EpCAMhigh CTCs via CellSearch®
4.4. Enrichment of EpCAMlow/negative Cells with Parsortix™ System
4.5. Detection and Isolation of EpCAMhigh and EpCAMlow/negative Cells via CellCelector™
4.5.1. Selection Criteria
4.5.2. Cell Isolation Parameters
4.6. Whole Genome Amplification of Isolated CTCs
4.7. Sanger Sequencing of the PIK3CA Exons 9 and 20 on CTCs
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CTC | Circulating Tumor Cell |
| EpCAM | Epithelial Cell Adhesion Molecule |
| FDA | Food and Drug Administration |
| EMT | Epithelial to Mesenchymal Transition |
| MBC | Metastatic Breast Cancer |
| PI3K | Phosphatidylinositol 3-Kinase |
| PIK3CA | Phosphatidylinositol 3-Kinase Catalytic subunit Alpha |
| WGA | Whole Genome Amplification |
| WT | Wild-type |
References
- Yu, M.; Stott, S.; Toner, M.; Maheswaran, S.; Haber, D.A. Circulating tumor cells: Approaches to isolation and characterization. J. Cell Biol. 2011, 192, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Allard, W.J.; Matera, J.; Miller, M.C.; Repollet, M.; Connelly, M.C.; Rao, C.; Tibbe, A.G.J.; Uhr, J.W.; Terstappen, L.W.M.M. Tumor Cells Circulate in the Peripheral Blood of All Major Carcinomas but not in Healthy Subjects or Patients With Nonmalignant Diseases. Clin. Cancer Res. 2004, 10, 6897–6904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Riethdorf, S.; Wu, G.; Wang, T.; Yang, K.; Peng, G.; Liu, J.; Pantel, K. Meta-Analysis of the Prognostic Value of Circulating Tumor Cells in Breast Cancer. Clin. Cancer Res. 2012, 18, 5701–5710. [Google Scholar] [CrossRef] [PubMed]
- Scher, H.I.; Jia, X.; de Bono, J.S.; Fleisher, M.; Pienta, K.J.; Raghavan, D.; Heller, G. Circulating Tumor Cell Number as a Prognostic Marker in Progressive Castration-Resistant Prostate Cancer: Use in Clinical Practice and Clinical Trials. Lancet Oncol. 2009, 10, 233–239. [Google Scholar] [CrossRef]
- Alix-Panabières, C.; Pantel, K. Technologies for detection of circulating tumor cells: Facts and vision. Lab Chip 2013, 14, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Banys, M.; Müller, V.; Melcher, C.; Aktas, B.; Kasimir-Bauer, S.; Hagenbeck, C.; Hartkopf, A.; Fehm, T. Circulating tumor cells in breast cancer. Clin. Chim. Acta 2013, 423, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Poruk, K.E. Circulating Tumor Cell Phenotype Predicts Recurrence and Survival in Pancreatic Adenocarcinoma. Ann. Surg. 2016, 264, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Bardia, A.; Wittner, B.S.; Stott, S.L.; Smas, M.E.; Ting, D.T.; Isakoff, S.J.; Ciciliano, J.C.; Wells, M.N.; Shah, A.M.; et al. Circulating Breast Tumor Cells Exhibit Dynamic Changes in Epithelial and Mesenchymal Composition. Science 2013, 339, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Gorges, T.M.; Tinhofer, I.; Drosch, M.; Röse, L.; Zollner, T.M.; Krahn, T. Circulating tumour cells escape from EpCAM-based detection due to epithelial-to-mesenchymal transition. BMC Cancer 2012, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Bednarz-Knoll, N.; Alix-Panabières, C.; Pantel, K. Plasticity of disseminating cancer cells in patients with epithelial malignancies. Cancer Metastasis Rev. 2012, 31, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- De Wit, S.; van Dalum, G.van; Lenferink, A.T.M.; Tibbe, A.G.J.; Hiltermann, T.J.N.; Groen, H.J.M.; van Rijn, C.J.M.; Terstappen, L.W.M.M. The detection of EpCAM+ and EpCAM– circulating tumor cells. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Schneck, H.; Gierke, B.; Uppenkamp, F.; Behrens, B.; Niederacher, D.; Stoecklein, N.H.; Templin, M.F.; Pawlak, M.; Fehm, T.; Neubauer, H. EpCAM-Independent Enrichment of Circulating Tumor Cells in Metastatic Breast Cancer. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Aktas, B. Stem cell and epithelial-mesenchymal transition markers are frequently overexpressed in circulating tumor cells of metastatic breast cancer patients. Breast Cancer Res. 2009, 11, R46. [Google Scholar] [CrossRef] [PubMed]
- Mego, M. Prognostic Value of EMT-Circulating Tumor Cells in Metastatic Breast Cancer Patients Undergoing High-Dose Chemotherapy with Autologous Hematopoietic Stem Cell Transplantation. J. Cancer 2012, 3, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Yokobori, T. Plastin3 is a novel marker for circulating tumor cells undergoing the epithelial-mesenchymal transition and is associated with colorectal cancer prognosis. Cancer Res. 2013, 73, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Ueo, H. Circulating tumour cell-derived plastin3 is a novel marker for predicting long-term prognosis in patients with breast cancer. Br. J. Cancer 2015, 112, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Lampignano, R.; Schneck, H.; Neumann, M.; Fehm, T.; Neubauer, H. Enrichment, Isolation and Molecular Characterization of EpCAM-Negative Circulating Tumor Cells. Adv. Exp. Med. Biol. 2017, 994, 181–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lv, J.; Wang, L.; Wang, X.; Ye, L. The impact of heterogeneity in phosphoinositide 3-kinase pathway in human cancer and possible therapeutic treatments. Semin. Cell Dev. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Troxell, L.M. PIK3CA/AKT1 Mutations in Breast Carcinoma: A Comprehensive Review of Experimental and Clinical Studies. J. Clin. Exp. Pathol. 2012. [Google Scholar] [CrossRef]
- Bader, A.G.; Kang, S.; Vogt, P.K. Cancer-specific mutations in PIK3CA are oncogenic in vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Liu, Z.; Wang, L.; Shin, E.; Loda, M.F.; Roberts, T.M. The oncogenic properties of mutant p110α and p110β phosphatidylinositol 3-kinases in human mammary epithelial cells. Proc. Natl. Acad. Sci. USA 2005, 102, 18443–18448. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Angulo, A.M.; Blumenschein, G.R. Defining biomarkers to predict sensitivity to PI3K/Akt/mTOR pathway inhibitors in breast cancer. Cancer Treat. Rev. 2013, 39, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Loi, S.; Michiels, S.; Baselga, J.; Bartlett, J.M.S.; Singhal, S.K.; Sabine, V.S.; Sims, A.H.; Sahmoud, T.; Dixon, J.M.; Piccart, M.J.; et al. PIK3CA Genotype and a PIK3CA Mutation-Related Gene Signature and Response to Everolimus and Letrozole in Estrogen Receptor Positive Breast Cancer. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cizkova, M.; Dujaric, M.-E.; Lehmann-Che, J.; Scott, V.; Tembo, O.; Asselain, B.; Pierga, J.-Y.; Marty, M.; de Cremoux, P.; Spyratos, F.; et al. Outcome impact of PIK3CA mutations in HER2-positive breast cancer patients treated with trastuzumab. Br. J. Cancer 2013, 108, 1807–1809. [Google Scholar] [CrossRef] [PubMed]
- Hanker, A.B.; Pfefferle, A.D.; Balko, J.M.; Kuba, M.G.; Young, C.D.; Sánchez, V.; Sutton, C.R.; Cheng, H.; Perou, C.M.; Zhao, J.J.; et al. Mutant PIK3CA accelerates HER2-driven transgenic mammary tumors and induces resistance to combinations of anti-HER2 therapies. Proc. Natl. Acad. Sci. USA 2013, 110, 14372–14377. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.; Ciraolo, E.; Gulluni, F.; Hirsch, E. Targeting PI3K in Cancer: Any Good News? Front. Oncol. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Cizkova, M.; Susini, A.; Vacher, S.; Cizeron-Clairac, G.; Andrieu, C.; Driouch, K.; Fourme, E.; Lidereau, R.; Bièche, I. PIK3CA mutation impact on survival in breast cancer patients and in ERα, PR and ERBB2-based subgroups. Breast Cancer Res. 2012, 14, R28. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.N.; Sathyanarayanan, S.; Bacco, A.D.; Chi, A.; Zhang, T.; Chen, A.H.; Dolinski, B.; Kraus, M.; Roberts, B.; Arthur, W.; et al. Pathway-Based Identification of Biomarkers for Targeted Therapeutics: Personalized Oncology with PI3K Pathway Inhibitors. Sci. Transl. Med. 2010, 2, 43ra55. [Google Scholar] [CrossRef] [PubMed]
- Schneck, H.; Blassl, C.; Meier-Stiegen, F.; Neves, R.P.; Janni, W.; Fehm, T.; Neubauer, H. Analysing the mutational status of PIK3CA in circulating tumor cells from metastatic breast cancer patients. Mol. Oncol. 2013, 7, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Polzer, B.; Medoro, G.; Pasch, S.; Fontana, F.; Zorzino, L.; Pestka, A.; Andergassen, U.; Meier-Stiegen, F.; Czyz, Z.T.; Alberter, B.; et al. Molecular profiling of single circulating tumor cells with diagnostic intention. EMBO Mol. Med. 2014, 6, 1371–1386. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.H.D.; Schneck, H.; Decker, Y.; Schömer, S.; Franken, A.; Endris, V.; Pfarr, N.; Weichert, W.; Niederacher, D.; Fehm, T.; et al. Isolation and characterization of circulating tumor cells using a novel workflow combining the CellSearch® system and the CellCelector™. Biotechnol. Prog. 2017, 33, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Hollestelle, A.; Elstrodt, F.; Nagel, J.H.A.; Kallemeijn, W.W.; Schutte, M. Phosphatidylinositol-3-OH Kinase or RAS Pathway Mutations in Human Breast Cancer Cell Lines. Mol. Cancer Res. 2007, 5, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, Y.; Mukohara, T.; Shimada, H.; Saijo, N.; Hirai, M.; Minami, H. Association between gain-of-function mutations in PIK3CA and resistance to HER2-targeted agents in HER2-amplified breast cancer cell lines. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2010, 21, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Bidard, F.-C.; Peeters, D.J.; Fehm, T.; Nolé, F.; Gisbert-Criado, R.; Mavroudis, D.; Grisanti, S.; Generali, D.; Garcia-Saenz, J.A.; Stebbing, J.; et al. Clinical validity of circulating tumour cells in patients with metastatic breast cancer: A pooled analysis of individual patient data. Lancet Oncol. 2014, 15, 406–414. [Google Scholar] [CrossRef]
- Banys-Paluchowski, M.; Krawczyk, N.; Meier-Stiegen, F.; Fehm, T. Circulating tumor cells in breast cancer—Current status and perspectives. Crit. Rev. Oncol. Hematol. 2016, 97, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Cabel, L.; Proudhon, C.; Gortais, H.; Loirat, D.; Coussy, F.; Pierga, J.-Y.; Bidard, F.-C. Circulating tumor cells: Clinical validity and utility. Int. J. Clin. Oncol. 2017, 22, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Lustberg, M.B.; Balasubramanian, P.; Miller, B.; Garcia-Villa, A.; Deighan, C.; Wu, Y.; Carothers, S.; Berger, M.; Ramaswamy, B.; Macrae, E.R.; et al. Heterogeneous atypical cell populations are present in blood of metastatic breast cancer patients. Breast Cancer Res. BCR 2014, 16, R23. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, M.; Peddibhotla, S.; Yin, W.; T. Scamardo, A.; George, G.C.; Hong, D.S.; Marchetti, D. The isolation and characterization of CTC subsets related to breast cancer dormancy. Sci. Rep. 2015, 5, 17533. [Google Scholar] [CrossRef] [PubMed]
- Xu, L. Optimization and Evaluation of a Novel Size Based Circulating Tumor Cell Isolation System. PLoS ONE 2015, 10, e0138032. [Google Scholar] [CrossRef] [PubMed]
- Hvichia, G.E.; Parveen, Z.; Wagner, C.; Janning, M.; Quidde, J.; Stein, A.; Müller, V.; Loges, S.; Neves, R.P.; Stoecklein, N.H.; et al. A novel microfluidic platform for size and deformability based separation and the subsequent molecular characterization of viable circulating tumor cells. Int. J. Cancer 2016, 138, 2894–2904. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, T.M.; Riethdorf, S.; Nees, J.; Hartkopf, A.D.; Schönfisch, B.; Domschke, C.; Sprick, M.R.; Schütz, F.; Brucker, S.Y.; Stefanovic, S.; et al. Impact of apoptotic circulating tumor cells (aCTC) in metastatic breast cancer. Breast Cancer Res. Treat. 2016, 160, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Jansson, S.; Bendahl, P.-O.; Larsson, A.-M.; Aaltonen, K.E.; Rydén, L. Prognostic impact of circulating tumor cell apoptosis and clusters in serial blood samples from patients with metastatic breast cancer in a prospective observational cohort. BMC Cancer 2016, 16, 433. [Google Scholar] [CrossRef] [PubMed]
- Spiliotaki, M.; Mavroudis, D.; Kapranou, K.; Markomanolaki, H.; Kallergi, G.; Koinis, F.; Kalbakis, K.; Georgoulias, V.; Agelaki, S. Evaluation of proliferation and apoptosis markers in circulating tumor cells of women with early breast cancer who are candidates for tumor dormancy. Breast Cancer Res. BCR 2014, 16. [Google Scholar] [CrossRef] [PubMed]
- Kallergi, G.; Konstantinidis, G.; Markomanolaki, H.; Papadaki, M.A.; Mavroudis, D.; Stournaras, C.; Georgoulias, V.; Agelaki, S. Apoptotic Circulating Tumor Cells in Early and Metastatic Breast Cancer Patients. Mol. Cancer Ther. 2013, 12, 1886–1895. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Basso, U.; Celadin, R.; Zilio, F.; Pucciarelli, S.; Aieta, M.; Barile, C.; Sava, T.; Bonciarelli, G.; Tumolo, S.; et al. M30 Neoepitope Expression in Epithelial Cancer: Quantification of Apoptosis in Circulating Tumor Cells by CellSearch Analysis. Clin. Cancer Res. 2010, 16, 5233–5243. [Google Scholar] [CrossRef] [PubMed]
- Dirican, E.; Akkiprik, M.; Özer, A. Mutation distributions and clinical correlations of PIK3CA gene mutations in breast cancer. Tumor Biol. 2016, 37, 7033–7045. [Google Scholar] [CrossRef] [PubMed]
- Mukohara, T. PI3K mutations in breast cancer: Prognostic and therapeutic implications. Breast Cancer Targets Ther. 2015, 7, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Markou, A.; Farkona, S.; Schiza, C.; Efstathiou, T.; Kounelis, S.; Malamos, N.; Georgoulias, V.; Lianidou, E. PIK3CA Mutational Status in Circulating Tumor Cells Can Change During Disease Recurrence or Progression in Patients with Breast Cancer. Clin. Cancer Res. 2014, 20, 5823–5834. [Google Scholar] [CrossRef] [PubMed]
- Pestrin, M.; Salvianti, F.; Galardi, F.; De Luca, F.; Turner, N.; Malorni, L.; Pazzagli, M.; Di Leo, A.; Pinzani, P. Heterogeneity of PIK3CA mutational status at the single cell level in circulating tumor cells from metastatic breast cancer patients. Mol. Oncol. 2015, 9, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Gasch, C.; Oldopp, T.; Mauermann, O.; Gorges, T.M.; Andreas, A.; Coith, C.; Müller, V.; Fehm, T.; Janni, W.; Pantel, K.; et al. Frequent detection of PIK3CA mutations in single circulating tumor cells of patients suffering from HER2-negative metastatic breast cancer. Mol. Oncol. 2016, 10, 1330–1343. [Google Scholar] [CrossRef] [PubMed]
- Cristofanilli, M.; Budd, G.T.; Ellis, M.J.; Stopeck, A.; Matera, J.; Miller, M.C. Circulating tumor cells, disease progression, and survival in metastatic breast cancer. N. Engl. J. Med. 2004, 351. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Ogunniyi, A.O.; Du, M.; Du, M.; Kretschmann, M.; Eberhardt, J.; Love, J.C. Development and optimization of a process for automated recovery of single cells identified by microengraving. Biotechnol. Prog. 2010, 26, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.A.; Schmidt-Kittler, O.; Schardt, J.A.; Pantel, K.; Speicher, M.R.; Riethmüller, G. Comparative genomic hybridization, loss of heterozygosity, and DNA sequence analysis of single cells. Proc. Natl. Acad. Sci. USA 1999, 96, 4494–4499. [Google Scholar] [CrossRef] [PubMed]
- Hurst, C.D.; Zuiverloon, T.C.; Hafner, C.; Zwarthoff, E.C.; Knowles, M.A. A SNaPshot assay for the rapid and simple detection of four common hotspot codon mutations in the PIK3CA gene. BMC Res. Notes 2009, 2, 66. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Patient ID | EpCAMhigh CTCs | EpCAMlow/negative CTCs | ||||
|---|---|---|---|---|---|---|---|
| Sorted CTCs | High Integrity WGA Products | Low Integrity WGA Products | Sorted CTCs | High Integrity WGA Products | Low INTEGRITY WGA Products | ||
| 1 | I | 5 | 0 | 5 | 19 | 0 | 19 |
| 2 | V | 1 | 0 | 1 | 2 | 0 | 2 |
| 3 | VI | 11 | 5 | 6 | 24 | 3 | 21 |
| 4 | VIII | 8 | 0 | 8 | 12 | 0 | 12 |
| 5 | IX | 26 | 11 | 15 | 16 | 2 | 14 |
| 6 | X | 13 | 4 | 9 | 5 | 0 | 5 |
| 7 | XI | 7 | 3 | 4 | 6 | 0 | 6 |
| 8 | XVI | 9 | 1 | 8 | 27 | 1 | 26 |
| 9 | XXIV (2) | 6 | 1 | 5 | 4 | 0 | 4 |
| 10 | XXXV | 8 | 2 | 6 | 18 | 3 | 15 |
| 11 | XLI | 2 | 1 | 1 | 5 | 1 | 4 |
| 12 | XLVI | 7 | 2 | 5 | 4 | 0 | 4 |
| 13 | XLVII | 4 | 0 | 4 | 3 | 1 | 2 |
| 107 | 30 | 77 | 145 | 11 | 134 | ||
| 28% | 72% | 8% | 92% | ||||
| EpCAMhigh CTCs | |||||
| # | Patient ID | PIK3CA Exon 9 Mutational Analysis | PIK3CA Exon 20 Mutational Analysis | ||
| Sequenced CTCs | Mutational Status | Sequenced CTCs | Mutational Status | ||
| 1 | I | 2 | WT | 1 | WT |
| 2 | VI | 5 | WT | 2 | WT |
| 3 | IX | 13 | WT | 11 | 2: p.H1047L (c.CAT > CTT); 9: WT |
| 4 | X | 7 | WT | 5 | 2: p.H1047R (c.CAT > CGT); 3: WT |
| 5 | XI | 1 | WT | 3 | WT |
| 6 | XVI | 3 | WT | 3 | WT |
| 7 | XXXV | 1 | WT | 3 | WT |
| 8 | XXXVI | 1 | WT | 1 | WT |
| 9 | XLVII | 3 | WT | 3 | WT |
| 10 | XLVII | 2 | WT | 2 | 1: p.H1047L (c.CAT > CTT); 1: WT |
| 38 | 34 | ||||
| EpCAMlow/negative CTCs | |||||
| # | Patient ID | PIK3CA Exon 9 Mutational Analysis | PIK3CA Exon 20 Mutational Analysis | ||
| Sequenced CTCs | Mutational Status | Sequenced CTCs | Mutational Status | ||
| 1 | I | 10 | 1: p.E545K (c.CAG > AAG); 9: WT | 7 | WT |
| 2 | VI | 7 | WT | 7 | WT |
| 3 | IX | 5 | 1: p.E545K (c.CAG > AAG); 4: WT | 3 | WT |
| 4 | X | 2 | WT | 1 | WT |
| 5 | XI | 1 | WT | 1 | WT |
| 6 | XVI | 0 | n.d. | 2 | WT |
| 7 | XXXV | 8 | WT | 8 | WT |
| 8 | XXXVI | 3 | WT | 3 | WT |
| 9 | XLVII | 1 | WT | 1 | WT |
| 10 | XLVII | 2 | WT | 2 | WT |
| 39 | 35 | ||||
| PIK3CA Exon | Primer Name | Sequence (5′→3′) | Primer Length (bp) |
|---|---|---|---|
| 9 | forward | CATCCGATGTACCTGATTGAACTGCATGCAGACAAAGAACAGCTCAAAGCAA | 52 |
| reverse | CATTCCTTAGATAGCTCGGAAGTCCATTGCATTTTAGCACTTACCTGTGAC | 52 | |
| 20 | forward | CATCCGATGTACCTGATTGAACTGCATGCATTGATGACATTGCATACATTCG | 52 |
| reverse | CATTCCTTAGATAGCTCGGAAGTCCATTGCGTGGAAGATCCAATCCATTT | 50 | |
| Sequencing | forward | TCCGATGTACCTGATTGAAC | 20 |
| reverse | TTCCTTAGATAGCTCGGAAG | 20 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lampignano, R.; Yang, L.; Neumann, M.H.D.; Franken, A.; Fehm, T.; Niederacher, D.; Neubauer, H. A Novel Workflow to Enrich and Isolate Patient-Matched EpCAMhigh and EpCAMlow/negative CTCs Enables the Comparative Characterization of the PIK3CA Status in Metastatic Breast Cancer. Int. J. Mol. Sci. 2017, 18, 1885. https://doi.org/10.3390/ijms18091885
Lampignano R, Yang L, Neumann MHD, Franken A, Fehm T, Niederacher D, Neubauer H. A Novel Workflow to Enrich and Isolate Patient-Matched EpCAMhigh and EpCAMlow/negative CTCs Enables the Comparative Characterization of the PIK3CA Status in Metastatic Breast Cancer. International Journal of Molecular Sciences. 2017; 18(9):1885. https://doi.org/10.3390/ijms18091885
Chicago/Turabian StyleLampignano, Rita, Liwen Yang, Martin H. D. Neumann, André Franken, Tanja Fehm, Dieter Niederacher, and Hans Neubauer. 2017. "A Novel Workflow to Enrich and Isolate Patient-Matched EpCAMhigh and EpCAMlow/negative CTCs Enables the Comparative Characterization of the PIK3CA Status in Metastatic Breast Cancer" International Journal of Molecular Sciences 18, no. 9: 1885. https://doi.org/10.3390/ijms18091885
APA StyleLampignano, R., Yang, L., Neumann, M. H. D., Franken, A., Fehm, T., Niederacher, D., & Neubauer, H. (2017). A Novel Workflow to Enrich and Isolate Patient-Matched EpCAMhigh and EpCAMlow/negative CTCs Enables the Comparative Characterization of the PIK3CA Status in Metastatic Breast Cancer. International Journal of Molecular Sciences, 18(9), 1885. https://doi.org/10.3390/ijms18091885