SLC9A3 Protein Is Critical for Acrosomal Formation in Postmeiotic Male Germ Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Solute Carrier Family 9 Isoform 3 (SLC9A3)
1.2. SLC9A3 and Male Reproductive Tract
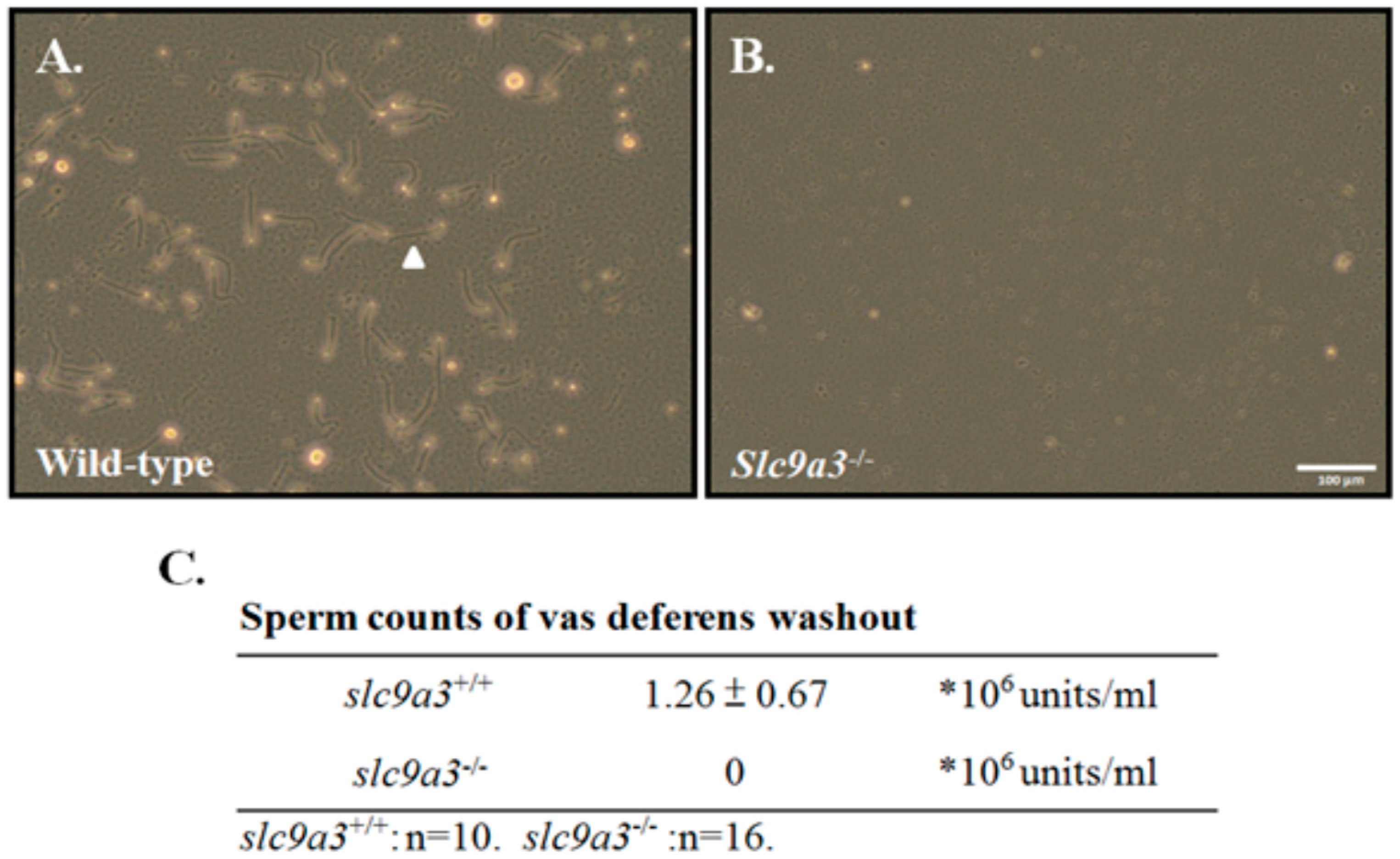
1.3. Loss of SLC9A3 Allele Causes Obstructive Azoospermia-Like Phenotype
2. Results
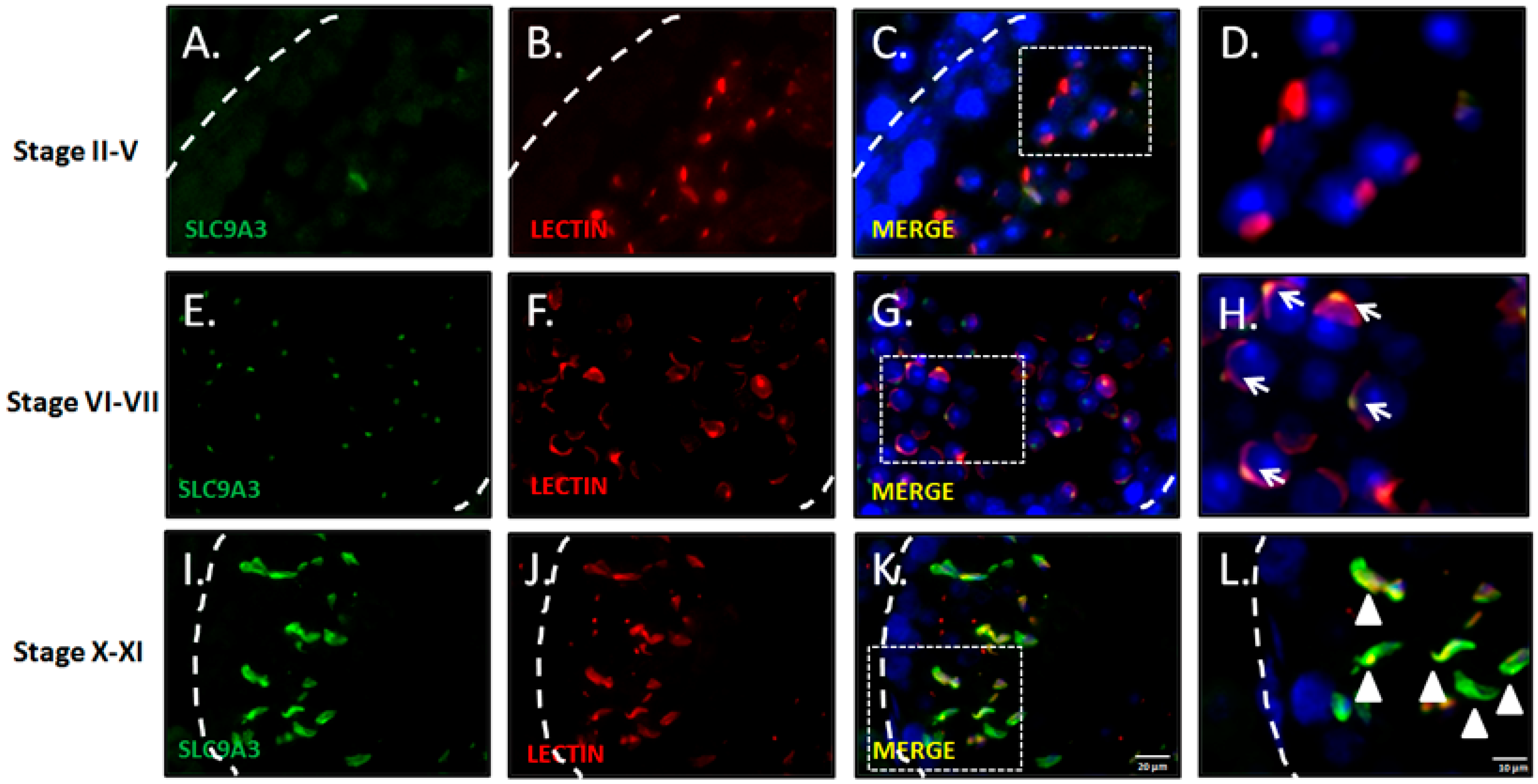
2.1. SLC9A3 is Specifically Expressed in Postmeiotic Male Germ Cells
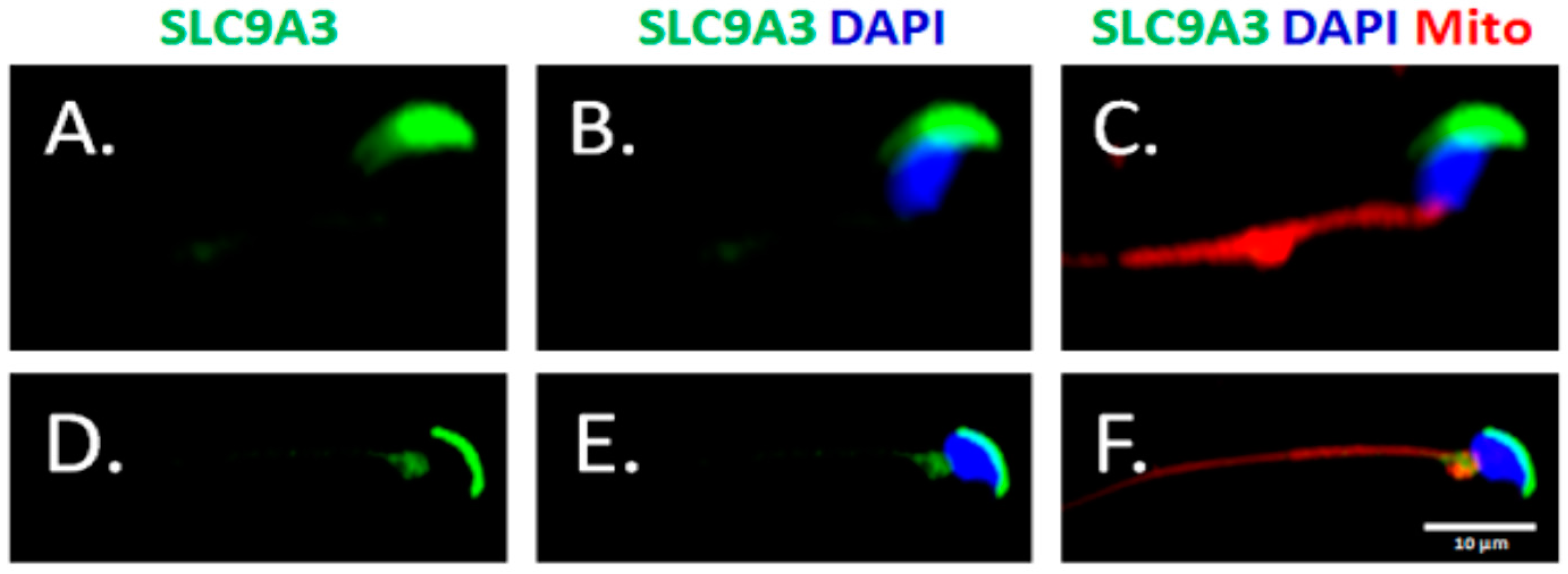
2.2. SLC9A3 is Involved in Acrosomal Formations
2.3. Loss of Slc9a3 Allele Disturbs the Spermiogenic Process of 35-Day-Old Mice
2.4. SLC9A3 is Essential for Acrosome Integrity
3. Experimental Section
3.1. Animals Preparation
3.2. Immunoblotting
3.3. Histological Analysis and Immunofluorescence Analysis
3.4. Separation of the Murine Male Germ Cell Populations
3.5. Electron Microscopy
4. Discussion
4.1. SLC9 Family Is Implicated in Sperm Formation
4.2. Oestrogen Receptor α Regulates SLC9A3 Expression in Male Reproduction
4.3. SLC9A3-CFTR Complexes and Male Germ Cell Production
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bookstein, C.; DePaoli, A.M.; Xie, Y.; Niu, P.; Musch, M.W.; Rao, M.C.; Chang, E.B. Na+/H+ exchangers, NHE-1 and NHE-3, of rat intestine. Expression and localization. J. Clin. Investig. 1994, 93, 106. [Google Scholar] [CrossRef] [PubMed]
- Hoogerwerf, W.A.; Tsao, S.C.; Devuyst, O.; Levine, S.A.; Yun, C.H.; Yip, J.W.; Cohen, M.E.; Wilson, P.D.; Lazenby, A.J.; Tse, C.M.; et al. NHE2 and NHE3 are human and rabbit intestinal brush-border proteins. Am. J. Physiol. 1996, 270, G29–G41. [Google Scholar] [CrossRef] [PubMed]
- Schultheis, P.J.; Clarke, L.L.; Meneton, P.; Miller, M.L.; Soleimani, M.; Gawenis, L.R.; Riddle, T.M.; Duffy, J.J.; Doetschman, T.; Wang, T.; et al. Renal and intestinal absorptive defects in mice lacking the NHE3 Na+/H+ exchanger. Nat. Genet. 1998, 19, 282–285. [Google Scholar] [PubMed]
- Hihnala, S.; Kujala, M.; Toppari, J.; Kere, J.; Holmberg, C.; Hoglund, P. Expression of SLC26A3, CFTR and NHE3 in the human male reproductive tract: Role in male subfertility caused by congenital chloride diarrhoea. Mol. Hum. Reprod. 2006, 12, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Bagnis, C.; Marsolais, M.; Biemesderfer, D.; Laprade, R.; Breton, S. Na+/H+-exchange activity and immunolocalization of NHE3 in rat epididymis. Am. J. Physiol. Renal Physiol. 2001, 280, F426–F436. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Lin, Y.H.; Wu, Y.N.; Chen, Y.L.; Lin, Y.C.; Cheng, C.Y.; Chiang, H.S. Loss of SLC9A3 decreases CFTR protein and causes obstructed azoospermia in mice. PLoS Genet. 2017, 13, e1006715. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Yun, C.C. Mechanisms of the regulation of the intestinal Na+/H+ exchanger NHE3. J. Biomed. Biotechnol. 2010, 2010, 238080. [Google Scholar] [CrossRef] [PubMed]
- Zachos, N.C.; Tse, M.; Donowitz, M. Molecular physiology of intestinal Na+/H+ exchange. Annu. Rev. Physiol. 2005, 67, 411–443. [Google Scholar] [CrossRef] [PubMed]
- Corvol, H.; Blackman, S.M.; Boelle, P.Y.; Gallins, P.J.; Pace, R.G.; Stonebraker, J.R.; Accurso, F.J.; Clement, A.; Collaco, J.M.; Dang, H.; et al. Genome-wide association meta-analysis identifies five modifier loci of lung disease severity in cystic fibrosis. Nat. Commun. 2015, 6, 8382. [Google Scholar] [CrossRef] [PubMed]
- Janecke, A.R.; Heinz-Erian, P.; Yin, J.; Petersen, B.S.; Franke, A.; Lechner, S.; Fuchs, I.; Melancon, S.; Uhlig, H.H.; Travis, S.; et al. Reduced sodium/proton exchanger NHE3 activity causes congenital sodium diarrhea. Hum. Mol. Genet. 2015, 24, 6614–6623. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J.; Kandasamy, R.A.; Shull, G.E. Molecular cloning of putative members of the Na/H exchanger gene family. cDNA cloning, deduced amino acid sequence, and mRNA tissue expression of the rat Na/H exchanger NHE-1 and two structurally related proteins. J. Boil. Chem. 1992, 267, 9331–9339. [Google Scholar]
- Pushkin, A.; Clark, I.; Kwon, T.H.; Nielsen, S.; Kurtz, I. Immunolocalization of NBC3 and NHE3 in the rat epididymis: Colocalization of NBC3 and the vacuolar H+-ATPase. J. Androl. 2000, 21, 708–720. [Google Scholar] [PubMed]
- Kaunisto, K.; Moe, O.W.; Pelto-Huikko, M.; Traebert, M.; Rajaniemi, H. An apical membrane Na+/H+ exchanger isoform, NHE-3, is present in the rat epididymal epithelium. Pflugers Archiv-Eur. J. Physiol. 2001, 442, 230–236. [Google Scholar] [CrossRef]
- Pholpramool, C.; Borwornpinyo, S.; Dinudom, A. Role of Na+/H+ exchanger 3 in the acidification of the male reproductive tract and male fertility. Clin. Exp. Pharmacol. Physiol. 2011, 38, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Clarke, L.; Nie, R.; Carnes, K.; Lai, L.W.; Lien, Y.H.; Verkman, A.; Lubahn, D.; Fisher, J.S.; Katzenellenbogen, B.S.; et al. Estrogen action and male fertility: Roles of the sodium/hydrogen exchanger-3 and fluid reabsorption in reproductive tract function. Proc. Natl. Acad. Sci. USA 2001, 98, 14132–14137. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.D.; Bernardino, R.L.; Neuhaus-Oliveira, A.; Sousa, M.; Sa, R.; Alves, M.G.; Oliveira, P.F. Physiology of Na+/H+ exchangers in the male reproductive tract: Relevance for male fertility. Biol. Reprod. 2014, 91, 11. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, E.; Shwachman, H.; Perlmutter, A.D.; Rule, A.; Khaw, K.T.; Holsclaw, D.S. Reproductive failure in males with cystic fibrosis. N. Engl. J. Med. 1968, 279, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Holsclaw, D.S.; Perlmutter, A.D.; Jockin, H.; Shwachman, H. Genital abnormalities in male patients with cystic fibrosis. J. Urol. 1971, 106, 568–574. [Google Scholar] [CrossRef]
- Kerem, E.; Corey, M.; Kerem, B.S.; Rommens, J.; Markiewicz, D.; Levison, H.; Tsui, L.C.; Durie, P. The relation between genotype and phenotype in cystic fibrosis—Analysis of the most common mutation (delta F508). N. Engl. J. Med. 1990, 323, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Cystic Fibrosis Genotype-Phenotype Consortium. Correlation between genotype and phenotype in patients with cystic fibrosis. N. Engl. J. Med. 1993, 329, 1308–1313. [Google Scholar]
- Kanavakis, E.; Tzetis, M.; Antoniadi, T.; Pistofidis, G.; Milligos, S.; Kattamis, C. Cystic fibrosis mutation screening in CBAVD patients and men with obstructive azoospermia or severe oligozoospermia. Mol. Hum. Reprod. 1998, 4, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.J.; Alper, O.M.; Wang, B.T.; Lee, M.H.; Lo, S.Y. Two novel null mutations in a Taiwanese cystic fibrosis patient and a survey of East Asian CFTR mutations. Am. J. Med. Genet. Part A 2003, 120A, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.C.; Wu, Y.N.; Chiang, H.S. Gene copy number variations in Asian patients with congenital bilateral absence of the vas deferens. Hum. Reprod. 2009, 24, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Grove, D.E.; Rosser, M.F.; Watkins, R.L.; Cyr, D.M. Analysis of CFTR folding and degradation in transiently transfected cells. Methods Mol. Boil. 2011, 741, 219–232. [Google Scholar]
- Lin, Y.H.; Lin, Y.M.; Kuo, Y.C.; Wang, Y.Y.; Kuo, P.L. Identification and characterization of a novel Rab GTPase-activating protein in spermatids. Int. J. Androl. 2010, 34, e358–e367. [Google Scholar] [CrossRef] [PubMed]
- Roos, A.; Boron, W.F. Intracellular pH. Physiol. Rev. 1981, 61, 296–434. [Google Scholar] [CrossRef] [PubMed]
- Donowitz, M.; Mohan, S.; Zhu, C.X.; Chen, T.E.; Lin, R.; Cha, B.; Zachos, N.C.; Murtazina, R.; Sarker, R.; Li, X. NHE3 regulatory complexes. J. Exp. Boil. 2009, 212, 1638–1646. [Google Scholar] [CrossRef] [PubMed]
- Fuster, D.G.; Alexander, R.T. Traditional and emerging roles for the SLC9 Na+/H+ exchangers. Pflugers Arch. Eur. J. Physiol. 2014, 466, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.M.; Schreiner, C.M.; Schultheis, P.J.; Miller, M.L.; Evans, R.L.; Vorhees, C.V.; Shull, G.E.; Scott, W.J. Targeted disruption of the murine Nhe1 locus induces ataxia, growth retardation, and seizures. Am. J. Physiol. 1999, 276, C788–C795. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; King, S.M.; Quill, T.A.; Doolittle, L.K.; Garbers, D.L. A new sperm-specific Na+/H+ exchanger required for sperm motility and fertility. Nat. Cell Boil. 2003, 5, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef] [PubMed]
- Eddy, E.M.; Washburn, T.F.; Bunch, D.O.; Goulding, E.H.; Gladen, B.C.; Lubahn, D.B.; Korach, K.S. Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinology 1996, 137, 4796–4805. [Google Scholar] [CrossRef] [PubMed]
- Joseph, A.; Shur, B.D.; Ko, C.; Chambon, P.; Hess, R.A. Epididymal hypo-osmolality induces abnormal sperm morphology and function in the estrogen receptor alpha knockout mouse. Biol. Reprod. 2010, 82, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Patrizio, P.; Ord, T.; Silber, S.J.; Asch, R.H. Cystic fibrosis mutations impair the fertilization rate of epididymal sperm from men with congenital absence of the vas deferens. Hum. Reprod. 1993, 8, 1259–1263. [Google Scholar] [CrossRef] [PubMed]
- Ahn, W.; Kim, K.H.; Lee, J.A.; Kim, J.Y.; Choi, J.Y.; Moe, O.W.; Milgram, S.L.; Lee, M.G. Regulatory interaction between the cystic fibrosis transmembrane conductance regulator and HCO3- salvage mechanisms in model systems and the mouse pancreatic duct. J. Boil. Chem. 2001, 276, 17236–17243. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ruan, Y.C.; Xu, W.M.; Chen, J.; Chan, H.C. Regulation of male fertility by CFTR and implications in male infertility. Hum. Reprod. Update 2012, 18, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.M.; Shi, Q.X.; Chen, W.Y.; Zhou, C.X.; Ni, Y.; Rowlands, D.K.; Yi Liu, G.; Zhu, H.; Ma, Z.G.; Wang, X.F.; et al. Cystic fibrosis transmembrane conductance regulator is vital to sperm fertilizing capacity and male fertility. Proc. Natl. Acad. Sci. USA 2007, 104, 9816–9821. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-Y.; Chiang, H.-S.; Cheng, C.-Y.; Wu, Y.-N.; Lin, Y.-C.; Liu, H.-C.; Tsai, W.-K.; Chen, Y.-L.; Lin, Y.-H. SLC9A3 Protein Is Critical for Acrosomal Formation in Postmeiotic Male Germ Cells. Int. J. Mol. Sci. 2018, 19, 103. https://doi.org/10.3390/ijms19010103
Wang Y-Y, Chiang H-S, Cheng C-Y, Wu Y-N, Lin Y-C, Liu H-C, Tsai W-K, Chen Y-L, Lin Y-H. SLC9A3 Protein Is Critical for Acrosomal Formation in Postmeiotic Male Germ Cells. International Journal of Molecular Sciences. 2018; 19(1):103. https://doi.org/10.3390/ijms19010103
Chicago/Turabian StyleWang, Ya-Yun, Han-Sun Chiang, Chiao-Yin Cheng, Yi-No Wu, Yung-Chih Lin, Hsuan-Che Liu, Wei-Kung Tsai, Yen-Lin Chen, and Ying-Hung Lin. 2018. "SLC9A3 Protein Is Critical for Acrosomal Formation in Postmeiotic Male Germ Cells" International Journal of Molecular Sciences 19, no. 1: 103. https://doi.org/10.3390/ijms19010103
APA StyleWang, Y.-Y., Chiang, H.-S., Cheng, C.-Y., Wu, Y.-N., Lin, Y.-C., Liu, H.-C., Tsai, W.-K., Chen, Y.-L., & Lin, Y.-H. (2018). SLC9A3 Protein Is Critical for Acrosomal Formation in Postmeiotic Male Germ Cells. International Journal of Molecular Sciences, 19(1), 103. https://doi.org/10.3390/ijms19010103