CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon


Abstract
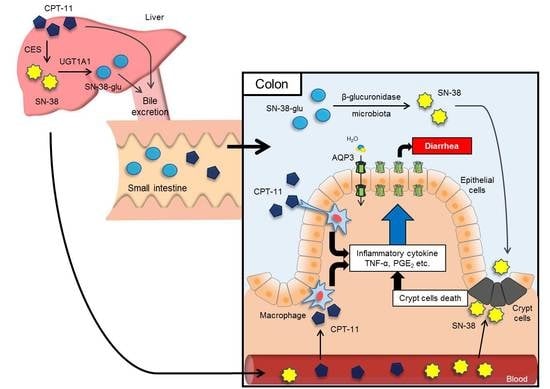
:
1. Introduction
2. Results
2.1. Effect of CPT-11 Dose and Administration Schedule on the Diarrhea Score and Mortality

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scale | Grade | Condition |
|---|---|---|
| 0 | Normal | Normal stool or absent |
| 1 | Slight | Slightly wet and soft stool |
| 2 | Moderate | Wet and unformed stool with moderate perianal staining of the coat |
| 3 | Severe | Watery stool with severe perianal staining of the coat |
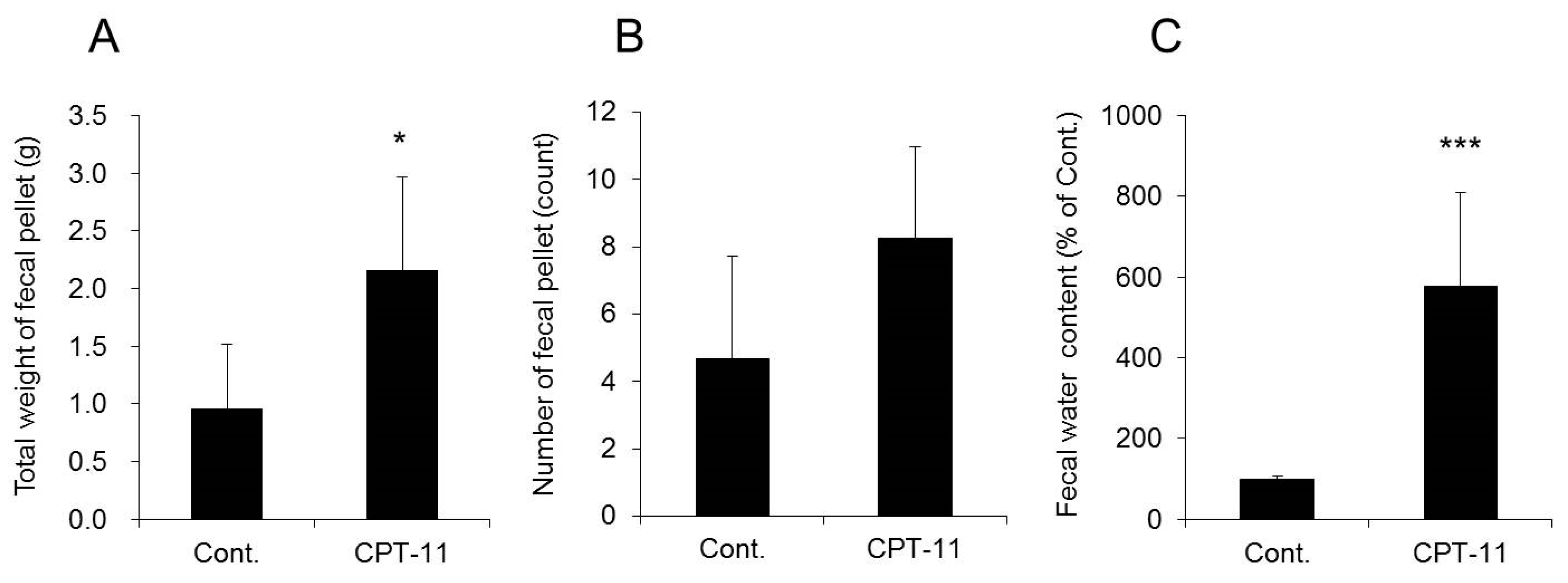
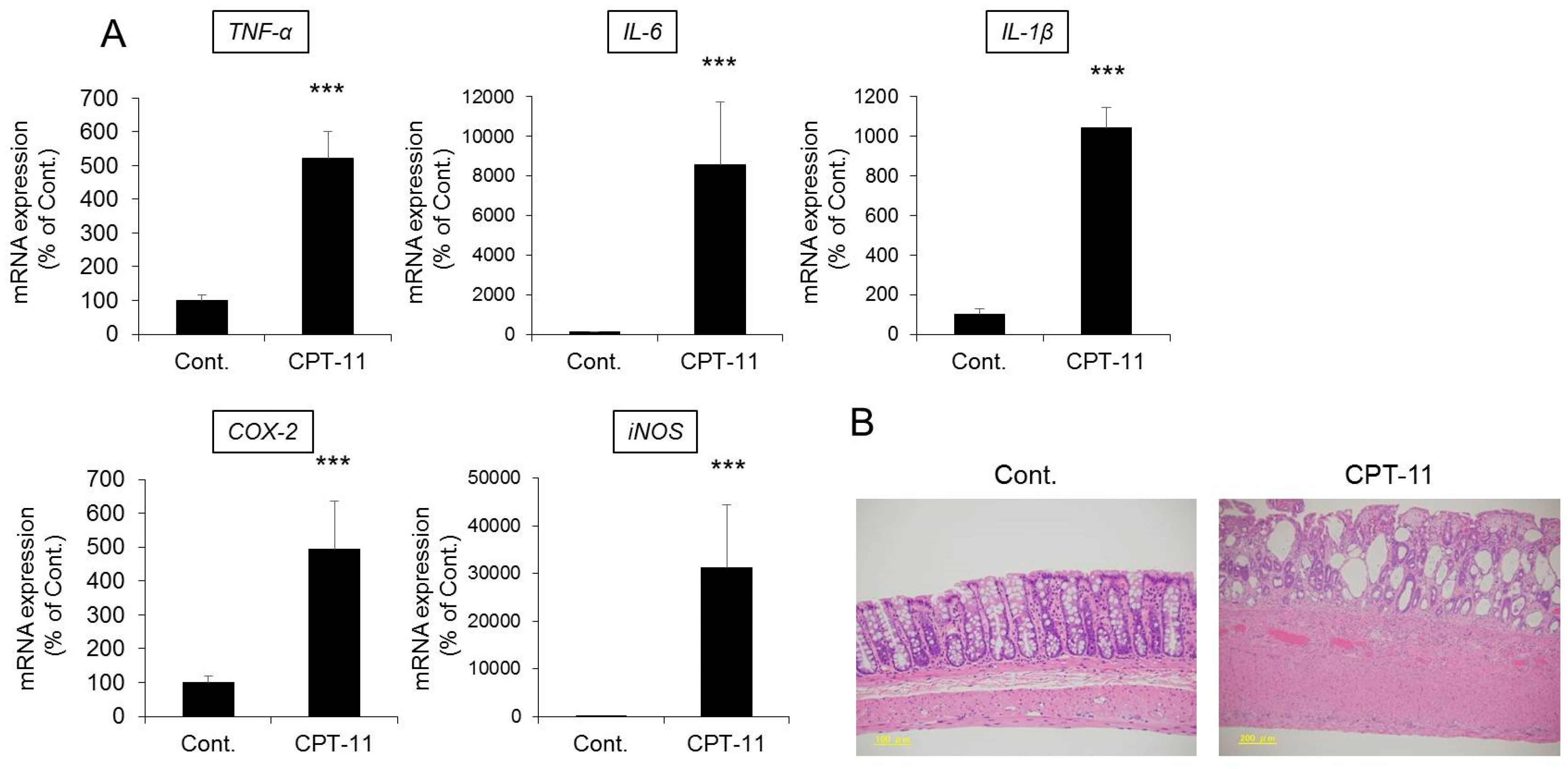
2.2. Evaluation of Diarrhea and Colitis in the CPT-11-Induced Delayed Diarrhea Rat Model
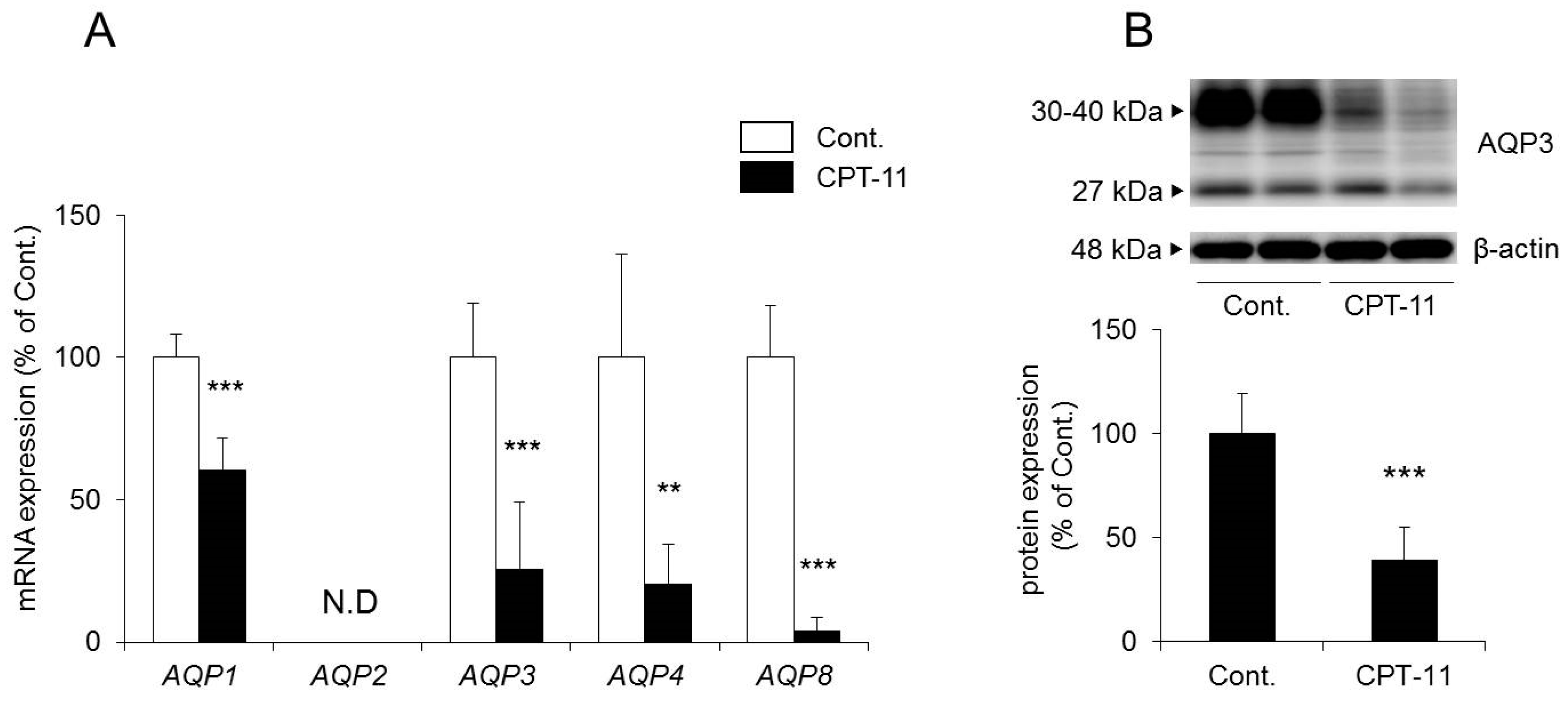
2.3. Changes in AQP in the Colon in the CPT-11-Induced Delayed Diarrhea Rat Model
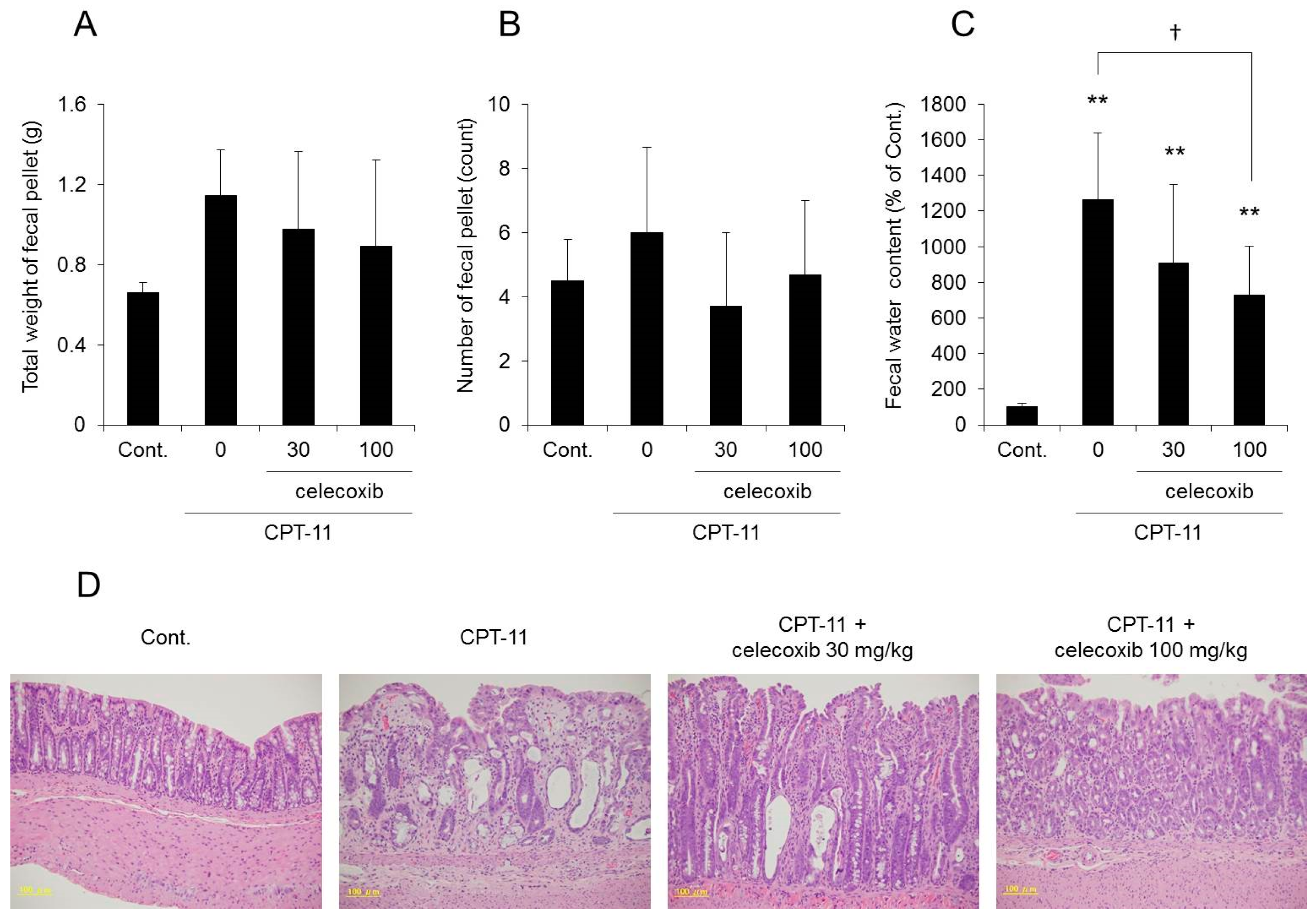
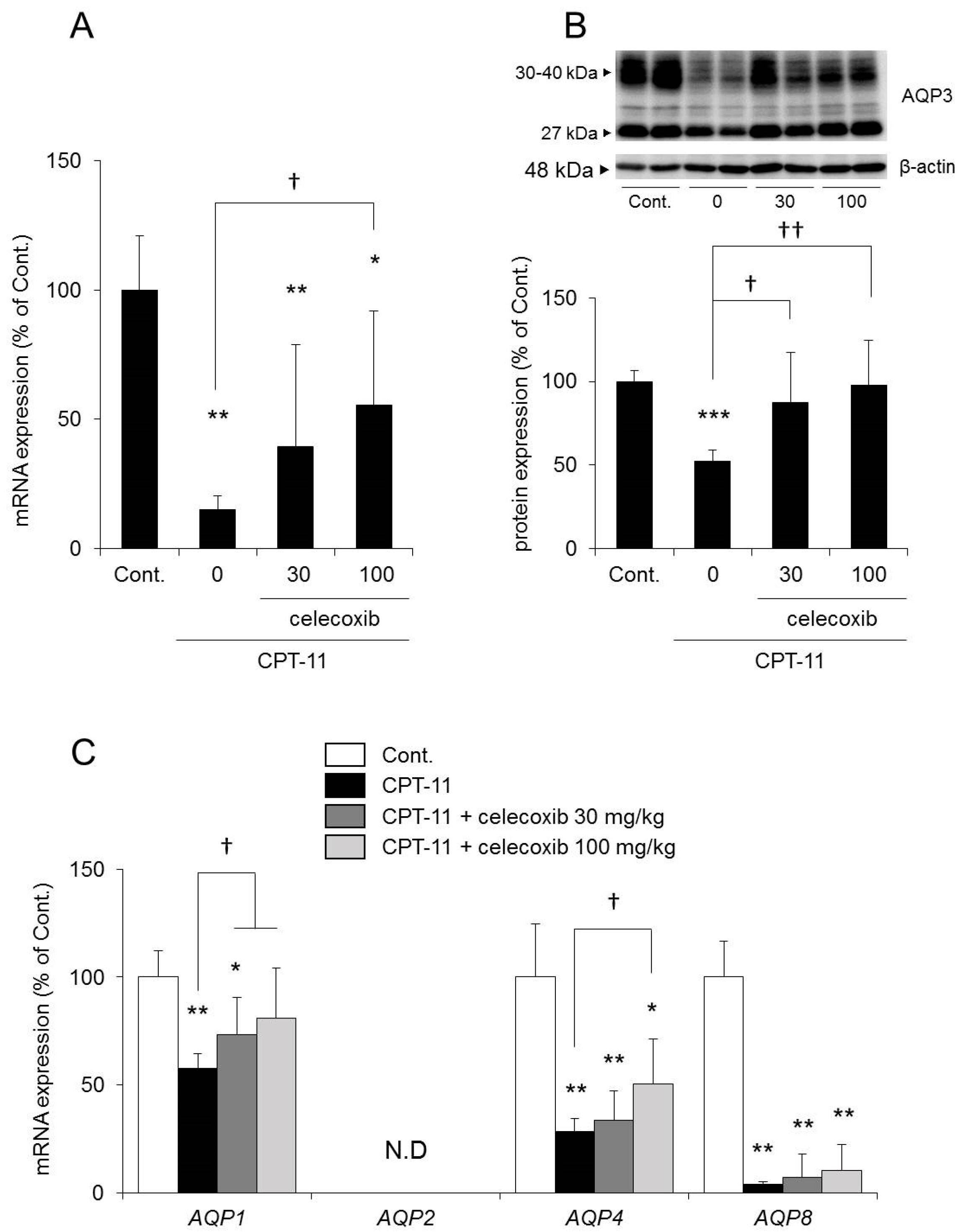
2.4. Effect of Celecoxib on the CPT-11-Induced Delayed Diarrhea Rat Model
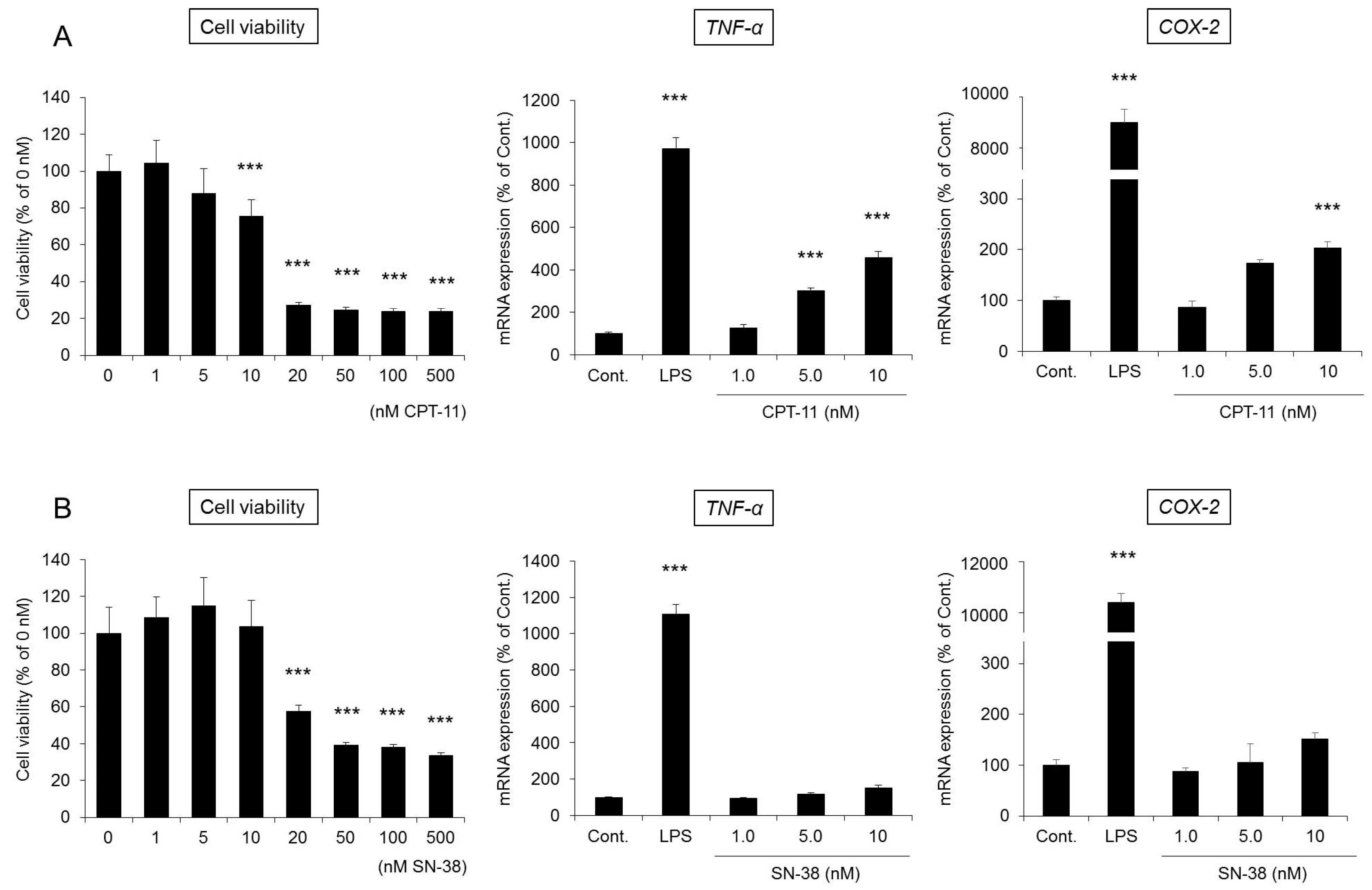
2.5. Involvement of Colonic Macrophages in CPT-11-Induced Delayed Diarrhea
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Treatment
4.4. Assessment of Diarrhea
4.5. HE Staining
4.6. RAW264 Cell Culture
4.7. WST-1 Assay
4.8. Total RNA Preparations and Real-Time RT-PCR
4.9. Extraction of the Plasma Membrane Fraction from the Rat Colons
4.10. Western Blotting
4.11. Immunohistochenistry
4.12. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AQP | Aquaporin |
| CFTR | Cystic fibrosis transmembrane conductance regulator |
| COX | Cyclooxygenase |
| CPT-11 | Camptothecin-11, irinotecan |
| DAPI | 4′,6-diamidino-2-phenylindole |
| HE | Hematoxylin and eosin |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| JNK | c-Jun N-terminal kinase |
| LPS | Lipopolysaccharide |
| NF-κB | Nuclear factor-kappa B |
| NLRP3 | NOD-like receptor protein-3 |
| PCR | polymerase chain reaction |
| PGE | Prostaglandin E |
| QOL | Quality of life |
| SN-38 | 7-ethyl-10-hydroxy-camptothecin |
| SN-38-glu | 7-ethyl-10-hydroxy-camptothecin glucronide |
| TNF | Tumor necrosis factor |
| UGT | Uridine diphosphate-glucuronosyltransferase |
| WST-1 | Water-soluble tetrazolium salt |
References
- Kawato, Y.; Aonuma, M.; Hirota, Y.; Kuga, H.; Sato, K. Intracellular roles of SN-38, a metabolite of the camptothecin derivative CPT-11, in the antitumor effect of CPT-11. Cancer Res. 1991, 51, 4187–4191. [Google Scholar] [PubMed]
- Shimada, Y.; Yoshino, M.; Wakui, A.; Nakao, I.; Futatsuki, K.; Sakata, Y.; Kambe, M.; Taguchi, T.; Ogawa, N. Phase II study of CPT-11, a new camptothecin derivative, in metastatic colorectal cancer. CPT-11 Gastrointestinal Cancer Study Group. J. Clin. Oncol. 1993, 11, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Bleiberg, H.; Cvitkovic, E. Characterisation and clinical management of CPT-11 (irinotecan)-induced adverse events: The European perspective. Eur. J. Cancer 1996, 32A (Suppl. 3), S18–S23. [Google Scholar] [CrossRef]
- Gandia, D.; Abigerges, D.; Armand, J.P.; Chabot, G.; Da Costa, L.; De Forni, M.; Mathieu-Boue, A.; Herait, P. CPT-11-induced cholinergic effects in cancer patients. J. Clin. Oncol. 1993, 11, 196–197. [Google Scholar] [CrossRef] [PubMed]
- Kawato, Y.; Sekiguchi, M.; Akahane, K.; Tsutomi, Y.; Hirota, Y.; Kuga, H.; Suzuki, W.; Hakusui, H.; Sato, K. Inhibitory activity of camptothecin derivatives against acetylcholinesterase in dogs and their binding activity to acetylcholine receptors in rats. J. Pharm. Pharmacol. 1993, 45, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Rothenberg, M.L.; Kuhn, J.G.; Burris, H.A., 3rd; Nelson, J.; Eckardt, J.R.; Tristan-Morales, M.; Hilsenbeck, S.G.; Weiss, G.R.; Smith, L.S.; Rodriguez, G.I.; et al. Phase I and pharmacokinetic trial of weekly CPT-11. J. Clin. Oncol. 1993, 11, 2194–2204. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.; King, C.D.; Whitington, P.F.; Green, M.D.; Roy, S.K.; Tephly, T.R.; Coffman, B.L.; Ratain, M.J. Genetic predisposition to the metabolism of irinotecan (CPT-11). Role of uridine diphosphate glucuronosyltransferase isoform 1A1 in the glucuronidation of its active metabolite (SN-38) in human liver microsomes. J. Clin. Investig. 1998, 101, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Rivory, L.P.; Bowles, M.R.; Robert, J.; Pond, S.M. Conversion of irinotecan (CPT-11) to its active metabolite, 7-ethyl-10-hydroxycamptothecin (SN-38), by human liver carboxylesterase. Biochem. Pharmacol. 1996, 52, 1103–2211. [Google Scholar] [CrossRef]
- Atsumi, R.; Okazaki, O.; Hakusui, H. Pharmacokinetics of SN-38 [(+)-(4S)-4,11-diethyl-4,9-dihydroxy-1H- pyrano[3′,4′:6,7]-indolizino[1,2-b]quinoline-3,14(4H,12H)-dione], an active metabolite of irinotecan, after a single intravenous dosing of 14C-SN-38 to rats. Biol. Pharm. Bull. 1995, 18, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Takasuna, K.; Hagiwara, T.; Hirohashi, M.; Kato, M.; Nomura, M.; Nagai, E.; Yokoi, T.; Kamataki, T. Involvement of β-glucuronidase in intestinal microflora in the intestinal toxicity of the antitumor camptothecin derivative irinotecan hydrochloride (CPT-11) in rats. Cancer Res. 1996, 56, 3752–3757. [Google Scholar] [PubMed]
- Gupta, E.; Lestingi, T.M.; Mick, R.; Ramirez, J.; Vokes, E.E.; Ratain, M.J. Metabolic fate of irinotecan in humans: Correlation of glucuronidation with diarrhea. Cancer Res. 1994, 54, 3723–3725. [Google Scholar] [PubMed]
- De Jong, F.A.; Kehrer, D.F.; Mathijssen, R.H.; Creemers, G.J.; de Bruijn, P.; van Schaik, R.H.; Planting, A.S.; van der Gaast, A.; Eskens, F.A.; Janssen, J.T.; et al. Prophylaxis of irinotecan-induced diarrhea with neomycin and potential role for UGT1A1*28 genotype screening: A double-blind, randomized, placebo-controlled study. Oncologist 2006, 11, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Flieger, D.; Klassert, C.; Hainke, S.; Keller, R.; Kleinschmidt, R.; Fischbach, W. Phase II clinical trial for prevention of delayed diarrhea with cholestyramine/levofloxacin in the second-line treatment with irinotecan biweekly in patients with metastatic colorectal carcinoma. Oncology 2007, 72, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Furman, W.L.; Crews, K.R.; Billups, C.; Wu, J.; Gajjar, A.J.; Daw, N.C.; Patrick, C.C.; Rodriguez-Galindo, C.; Stewart, C.F.; Dome, J.S.; et al. Cefixime allows greater dose escalation of oral irinotecan: A phase I study in pediatric patients with refractory solid tumors. J. Clin. Oncol. 2006, 24, 563–570. [Google Scholar] [CrossRef] [PubMed]
- McGregor, L.M.; Stewart, C.F.; Crews, K.R.; Tagen, M.; Wozniak, A.; Wu, J.; McCarville, M.B.; Navid, F.; Santana, V.M.; Houghton, P.J.; et al. Dose escalation of intravenous irinotecan using oral cefpodoxime: A phase I study in pediatric patients with refractory solid tumors. Pediatr. Blood Cancer 2012, 58, 372–379. [Google Scholar] [CrossRef] [PubMed]
- McNall-Knapp, R.Y.; Williams, C.N.; Reeves, E.N.; Heideman, R.L.; Meyer, W.H. Extended phase I evaluation of vincristine, irinotecan, temozolomide, and antibiotic in children with refractory solid tumors. Pediatr. Blood Cancer 2010, 54, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Kondo, T.; Kamiyama, Y.; Kano, Y.; Tominaga, K. Preventive effect of Kampo medicine (Hangeshashin-to) against irinotecan-induced diarrhea in advanced non-small-cell lung cancer. Cancer Chemother. Pharmacol. 2003, 51, 403–406. [Google Scholar] [PubMed]
- Sakata, Y.; Suzuki, H.; Kamataki, T. Preventive effect of TJ-14, a kampo (Chinese herb) medicine, on diarrhea induced by irinotecan hydrochloride (CPT-11). Gan Kagaku Ryoho. Cancer Chemother. 1994, 21, 1241–1244. [Google Scholar]
- Michael, M.; Brittain, M.; Nagai, J.; Feld, R.; Hedley, D.; Oza, A.; Siu, L.; Moore, M.J. Phase II study of activated charcoal to prevent irinotecan-induced diarrhea. J. Clin. Oncol. 2004, 22, 4410–4417. [Google Scholar] [CrossRef] [PubMed]
- Sergio, G.C.; Felix, G.M.; Luis, J.V. Activated charcoal to prevent irinotecan-induced diarrhea in children. Pediatr. Blood Cancer 2008, 51, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Abigerges, D.; Chabot, G.G.; Armand, J.P.; Herait, P.; Gouyette, A.; Gandia, D. Phase I and pharmacologic studies of the camptothecin analog irinotecan administered every 3 weeks in cancer patients. J. Clin. Oncol. 1995, 13, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Kobayashi, K.; Akiyama, Y.; Soma, T.; Handa, S.; Kudoh, S.; Kudo, K. Prevention of irinotecan (CPT-11)-induced diarrhea by oral alkalization combined with control of defecation in cancer patients. Int. J. Cancer 2001, 92, 269–275. [Google Scholar] [CrossRef]
- Hecht, J.R. Gastrointestinal toxicity or irinotecan. Oncology 1998, 12, 72–78. [Google Scholar] [PubMed]
- Ikarashi, N.; Baba, K.; Ushiki, T.; Kon, R.; Mimura, A.; Toda, T.; Ishii, M.; Ochiai, W.; Sugiyama, K. The laxative effect of bisacodyl is attributable to decreased aquaporin-3 expression in the colon induced by increased PGE2 secretion from macrophages. Am. J. Physiol.-Gastrointest. Liver Physiol. 2011, 301, G887–G895. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, N.; Kon, R.; Iizasa, T.; Suzuki, N.; Hiruma, R.; Suenaga, K.; Toda, T.; Ishii, M.; Hoshino, M.; Ochiai, W.; et al. Inhibition of aquaporin-3 water channel in the colon induces diarrhea. Biol. Pharm. Bull. 2012, 35, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Kon, R.; Ikarashi, N.; Hayakawa, A.; Haga, Y.; Fueki, A.; Kusunoki, Y.; Tajima, M.; Ochiai, W.; Machida, Y.; Sugiyama, K. Morphine-Induced Constipation Develops With Increased Aquaporin-3 Expression in the Colon via Increased Serotonin Secretion. Toxicol. Sci. 2015, 145, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Kon, R.; Ikarashi, N.; Nagoya, C.; Takayama, T.; Kusunoki, Y.; Ishii, M.; Ueda, H.; Ochiai, W.; Machida, Y.; Sugita, K.; et al. Rheinanthrone, a metabolite of sennoside A, triggers macrophage activation to decrease aquaporin-3 expression in the colon, causing the laxative effect of rhubarb extract. J. Ethnopharm. 2014, 152, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.J.; Bowen, J.M.; Inglis, M.R.; Cummins, A.G.; Keefe, D.M. Irinotecan causes severe small intestinal damage, as well as colonic damage, in the rat with implanted breast cancer. J. Gastroenterol. Hepatol. 2003, 18, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Logan, R.M.; Gibson, R.J.; Bowen, J.M.; Stringer, A.M.; Sonis, S.T.; Keefe, D.M. Characterisation of mucosal changes in the alimentary tract following administration of irinotecan: Implications for the pathobiology of mucositis. Cancer Chemother. Pharmacol. 2008, 62, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, P.; Cid, L.P.; Vio, C.P.; Sepulveda, F.V. Aquaporin-2, a regulated water channel, is expressed in apical membranes of rat distal colon epithelium. Am. J. Physiol.-Gastrointest. Liver Physiol. 2001, 281, G856–G863. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Yamamoto, T.; Tani, T.; Nihei, K.; Kondo, D.; Funaki, H.; Yaoita, E.; Kawasaki, K.; Sato, N.; Hatakeyama, K.; et al. Expression and localization of aquaporins in rat gastrointestinal tract. Am. J. Physiol. 1999, 276, C621–C627. [Google Scholar] [CrossRef] [PubMed]
- Silberstein, C.; Kierbel, A.; Amodeo, G.; Zotta, E.; Bigi, F.; Berkowski, D.; Ibarra, C. Functional characterization and localization of AQP3 in the human colon. Braz. J. Med. Biol. Res. 1999, 32, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Yde, J.; Keely, S.; Wu, Q.; Borg, J.F.; Lajczak, N.; O’Dwyer, A.; Dalsgaard, P.; Fenton, R.A.; Moeller, H.B. Characterization of AQPs in Mouse, Rat, and Human Colon and Their Selective Regulation by Bile Acids. Front. Nutr. 2016, 3, 46. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Hayakawa, T.; Aburada, M.; Komatsu, Y.; Kamataki, T. Preventive effects of Hange-shashin-to on irinotecan hydrochloride-caused diarrhea and its relevance to the colonic prostaglandin E2 and water absorption in the rat. Jpn. J. Pharmacol. 1997, 75, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Hayakawa, T.; Ishige, A.; Aburada, M.; Komatsu, Y. The effects of Hange-shashin-to on the content of prostaglandin E2 and water absorption in the large intestine of rats. Biol. Pharm. Bull. 1997, 20, 954–957. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Hayakawa, T.; Togashi, Y.; Kamataki, T. Relevance of irinotecan hydrochloride-induced diarrhea to the level of prostaglandin E2 and water absorption of large intestine in rats. Jpn. J. Pharmacol. 1997, 75, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Saitoh, K.; Ishige, A.; Komatsu, Y. Mechanisms by which Hange-shashin-to reduces prostaglandin E2 levels. Biol. Pharm. Bull. 1998, 21, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Trifan, O.C.; Durham, W.F.; Salazar, V.S.; Horton, J.; Levine, B.D.; Zweifel, B.S.; Davis, T.W.; Masferrer, J.L. Cyclooxygenase-2 inhibition with celecoxib enhances antitumor efficacy and reduces diarrhea side effect of CPT-11. Cancer Res. 2002, 62, 5778–5784. [Google Scholar] [PubMed]
- Zheng, S.; Brown, M.C.; Taffet, S.M. Lipopolysaccharide stimulates both nuclear localization of the nuclear factor kappa B 50-kDa subunit and loss of the 105-kDa precursor in RAW264 macrophage-like cells. J. Biol. Chem. 1993, 268, 17233–17239. [Google Scholar] [PubMed]
- Lima-Junior, R.C.; Freitas, H.C.; Wong, D.V.; Wanderley, C.W.; Nunes, L.G.; Leite, L.L.; Miranda, S.P.; Souza, M.H.; Brito, G.A.; Magalhaes, P.J.; et al. Targeted inhibition of IL-18 attenuates irinotecan-induced intestinal mucositis in mice. Br. J. Pharmacol. 2014, 171, 2335–2350. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Sato, T.; Hamada, N.; Yasue, M.; Ikari, A.; Kakinoki, B.; Takeguchi, N. Thromboxane A2, released by the anti-tumour drug irinotecan, is a novel stimulator of Cl− secretion in isolated rat colon. J. Physiol. 1997, 505 Pt 1, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, X.; Wang, W.; Li, L.; Xu, Q.; Wu, X.; Gu, Y. CPT-11 activates NLRP3 inflammasome through JNK and NF-kappaB signalings. Toxicol. Appl. Pharmacol. 2015, 289, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Bondy, C.; Chin, E.; Smith, B.L.; Preston, G.M.; Agre, P. Developmental gene expression and tissue distribution of the CHIP28 water-channel protein. Proc. Natl. Acad. Sci. USA 1993, 90, 4500–4504. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Marples, D. Expression of the AQP-1 water channel in normal human tissues: A semiquantitative study using tissue microarray technology. Am. J. Physiol.-Cell Physiol. 2004, 286, C529–C537. [Google Scholar] [CrossRef] [PubMed]
- Frigeri, A.; Nicchia, G.P.; Balena, R.; Nico, B.; Svelto, M. Aquaporins in skeletal muscle: Reassessment of the functional role of aquaporin-4. FASEB J. 2004, 18, 905–907. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, Y.; Takahashi, J.; Shibuya, S.; Inoue, M.; Kojima, H.; Oniki, H.; Arata, S.; Hara, H.; Jimi, T.; Shioda, S.; et al. Generation of muscle aquaporin 4 overexpressing transgenic mouse: Its characterization at RNA and protein levels including freeze-fracture study. Micron 2007, 38, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Jimi, T.; Wakayama, Y.; Murahashi, M.; Shibuya, S.; Inoue, M.; Hara, H.; Matsuzaki, Y.; Uemura, N. Aquaporin 4: Lack of mRNA expression in the rat regenerating muscle fiber under denervation. Neurosci. Lett. 2000, 291, 93–96. [Google Scholar] [CrossRef]
- Fischer, H.; Stenling, R.; Rubio, C.; Lindblom, A. Differential expression of aquaporin 8 in human colonic epithelial cells and colorectal tumors. BMC Physiol. 2001, 1, 1. [Google Scholar] [CrossRef]
- Laforenza, U.; Cova, E.; Gastaldi, G.; Tritto, S.; Grazioli, M.; LaRusso, N.F.; Splinter, P.L.; D’Adamo, P.; Tosco, M.; Ventura, U. Aquaporin-8 is involved in water transport in isolated superficial colonocytes from rat proximal colon. J. Nutr. 2005, 135, 2329–2336. [Google Scholar] [PubMed]
- Hara-Chikuma, M.; Verkman, A.S. Aquaporin-3 facilitates epidermal cell migration and proliferation during wound healing. J. Mol. Med. 2008, 86, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.H.; Verkman, A.S. Aquaporin-3-dependent cell migration and proliferation during corneal re-epithelialization. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4365–4372. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajah, J.R.; Zhao, D.; Verkman, A.S. Impaired enterocyte proliferation in aquaporin-3 deficiency in mouse models of colitis. Gut 2007, 56, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.; Zhang, W.; Ren, A.; Arora, K.; Sinha, C.; Yarlagadda, S.; Woodrooffe, K.; Schuetz, J.D.; Valasani, K.R.; de Jonge, H.R.; et al. Compartmentalized accumulation of cAMP near complexes of multidrug resistance protein 4 (MRP4) and cystic fibrosis transmembrane conductance regulator (CFTR) contributes to drug-induced diarrhea. J. Biol. Chem. 2015, 290, 11246–11257. [Google Scholar] [CrossRef] [PubMed]
- Stringer, A.M.; Gibson, R.J.; Bowen, J.M.; Logan, R.M.; Ashton, K.; Yeoh, A.S.; Al-Dasooqi, N.; Keefe, D.M. Irinotecan-induced mucositis manifesting as diarrhoea corresponds with an amended intestinal flora and mucin profile. Int. J. Exp. Pathol. 2009, 90, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Stringer, A.M.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Yeoh, A.S.; Keefe, D.M. Faecal microflora and β-glucuronidase expression are altered in an irinotecan-induced diarrhea model in rats. Cancer Biol. Ther. 2008, 7, 1919–1925. [Google Scholar] [CrossRef] [PubMed]
- Marples, D.; Knepper, M.A.; Christensen, E.I.; Nielsen, S. Redistribution of aquaporin-2 water channels induced by vasopressin in rat kidney inner medullary collecting duct. Am. J. Physiol. 1995, 269, C655–C664. [Google Scholar] [CrossRef] [PubMed]







| CPT-11 Dose (mg/kg/day) | Schedule | Total CPT-11 (mg/kg rat) | Average Diarrhea Grade | Body Weight Loss (% of Control) | Mortality (%) |
|---|---|---|---|---|---|
| 60 | 4 day | 240 | 0.2 | 5.1 | 0 |
| 80 | 4 day | 320 | 1.25 | 11.8 | 0 |
| 100 | 3 day | 300 | 2 | 16.1 | 0 |
| 100 | 4 day | 400 | 2.5 | 23.3 | 0 |
| 120 | 3 day | 360 | 2.5 | 26.7 | 0 |
| 120 | 4 day | 480 | 2.7 | 25.2 | 0 |
| 150 | 1 day | 150 | 0 | 0 | 50 |
| 150 | 2 day | 300 | 0 | 0 | 100 |
| Group | Incidence of Diarrhea Scale | Average Diarrhea Grade | Body Weight Loss (% of Control) | |
|---|---|---|---|---|
| Grade 2 (%) | Grade 3 (%) | |||
| Control | 0 | 0 | 0 | 0 |
| CPT-11 alone | 0 | 100 | 3 | 28.5 |
| + celecoxib 30 mg/kg | 42.9 | 57.1 | 2.6 | 26.7 |
| + celecoxib 100 mg/kg | 83.3 | 16.7 | 2.2 | 28.0 |
| Colon Condition | Control | CPT-11 Alone | CPT-11 + Celecoxib | |
|---|---|---|---|---|
| 30 mg/kg | 100 mg/kg | |||
| Disruption of epithelial cells | − | − | − | − |
| Disruption of crypt cells | − | ++ | + | + |
| Inflammatory cell infiltration | − | ++ | ++ | + |
| Swelling | − | + | − | − |
| Regeneration of epithelial cells | − | ++ | +++ | +++ |
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| rTNF-α | GAAACACACGAGACGCTGAAGT | CACTGGATCCCGGAATGTCGAT |
| rIL-1β | TCAGGCTTCCTTGTGCAAGTGT | ACAGGTCATTCTCCTCACTGTC |
| rIL-6 | TAGTCCTTCCTACCCCAACTTC | GCCGAGTAGACCTCATAGTGAC |
| rCOX-2 | GCTGATGACTGCCCAACTC | GATCCGGGATGAACTCTCTC |
| riNOS | CAAGCACATTTGGCAATGGA | GCCAAATACCGCATACCTGA |
| rAQP1 | CTGGTGCTGTGCGTTCTG | GTCCAAGAGCCACAGACAAG |
| rAQP2 | GGTTCCCAGTGCAGAGTAG | GAGGGTAGCTCAAGGCTTC |
| rAQP3 | CCCCTTGTGATGCCTCTC | CCCTAGCTGGCAGAGTTC |
| rAQP4 | AGGAGGACCCAGGCAATG | GGCAAGGTCTCATGCCATC |
| rAQP8 | TTGGGCTCCGCTCTCTTC | CAAGGCCAGCCCATGAG |
| rβ-actin | GCCACTGCCGCATCCTCTTG | CGGAACCGCTCATTGCCGAT |
| mTNF-α | ATGGACACCAAACATTTCCTGC | CCAGTGGAGAGCCGATTCC |
| mCOX-2 | CAGGGCCCTTCCTCCCGTAG | GCCTTGGGGGTCAGGGATGA |
| m18S rRNA | GTCTGTGATGCCCTTAGATG | AGCTTATGACCCGCACTTAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kon, R.; Tsubota, Y.; Minami, M.; Kato, S.; Matsunaga, Y.; Kimura, H.; Murakami, Y.; Fujikawa, T.; Sakurai, R.; Tomimoto, R.; et al. CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon. Int. J. Mol. Sci. 2018, 19, 170. https://doi.org/10.3390/ijms19010170
Kon R, Tsubota Y, Minami M, Kato S, Matsunaga Y, Kimura H, Murakami Y, Fujikawa T, Sakurai R, Tomimoto R, et al. CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon. International Journal of Molecular Sciences. 2018; 19(1):170. https://doi.org/10.3390/ijms19010170
Chicago/Turabian StyleKon, Risako, Yuika Tsubota, Moe Minami, Saki Kato, Yukari Matsunaga, Hiroshi Kimura, Yuta Murakami, Tetsuya Fujikawa, Ryoya Sakurai, Rei Tomimoto, and et al. 2018. "CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon" International Journal of Molecular Sciences 19, no. 1: 170. https://doi.org/10.3390/ijms19010170
APA StyleKon, R., Tsubota, Y., Minami, M., Kato, S., Matsunaga, Y., Kimura, H., Murakami, Y., Fujikawa, T., Sakurai, R., Tomimoto, R., Machida, Y., Ikarashi, N., & Sugiyama, K. (2018). CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon. International Journal of Molecular Sciences, 19(1), 170. https://doi.org/10.3390/ijms19010170