Proteomic Analysis Reveals the Dynamic Role of Silicon in Alleviation of Hyperhydricity in Carnation Grown In Vitro




Abstract
:1. Introduction
2. Results and Discussion
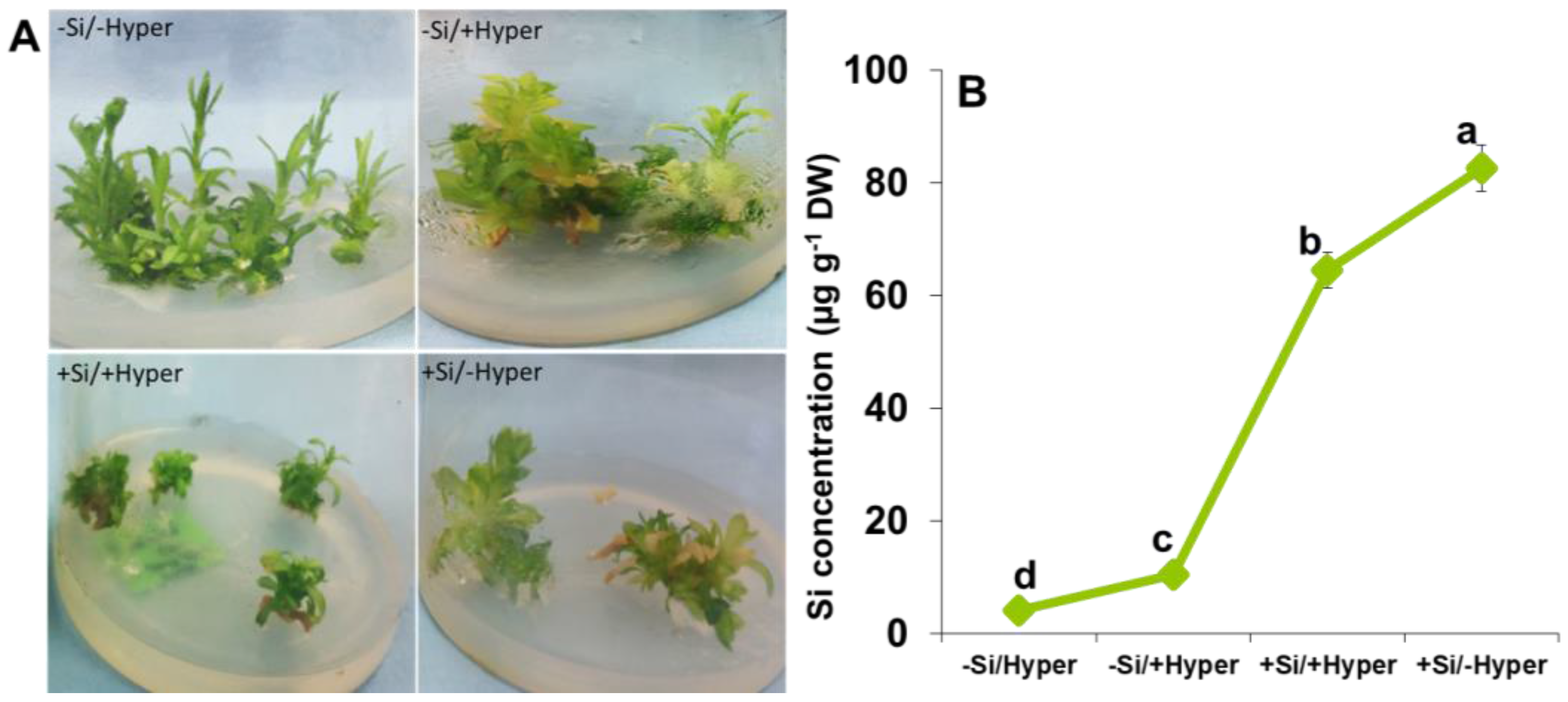
2.1. Morphological Changes and Si Concentration in Response to Hyperhydricity and Si Supply
2.2. Oxidative Stress in Response to Hyperhydricity and Si Supply
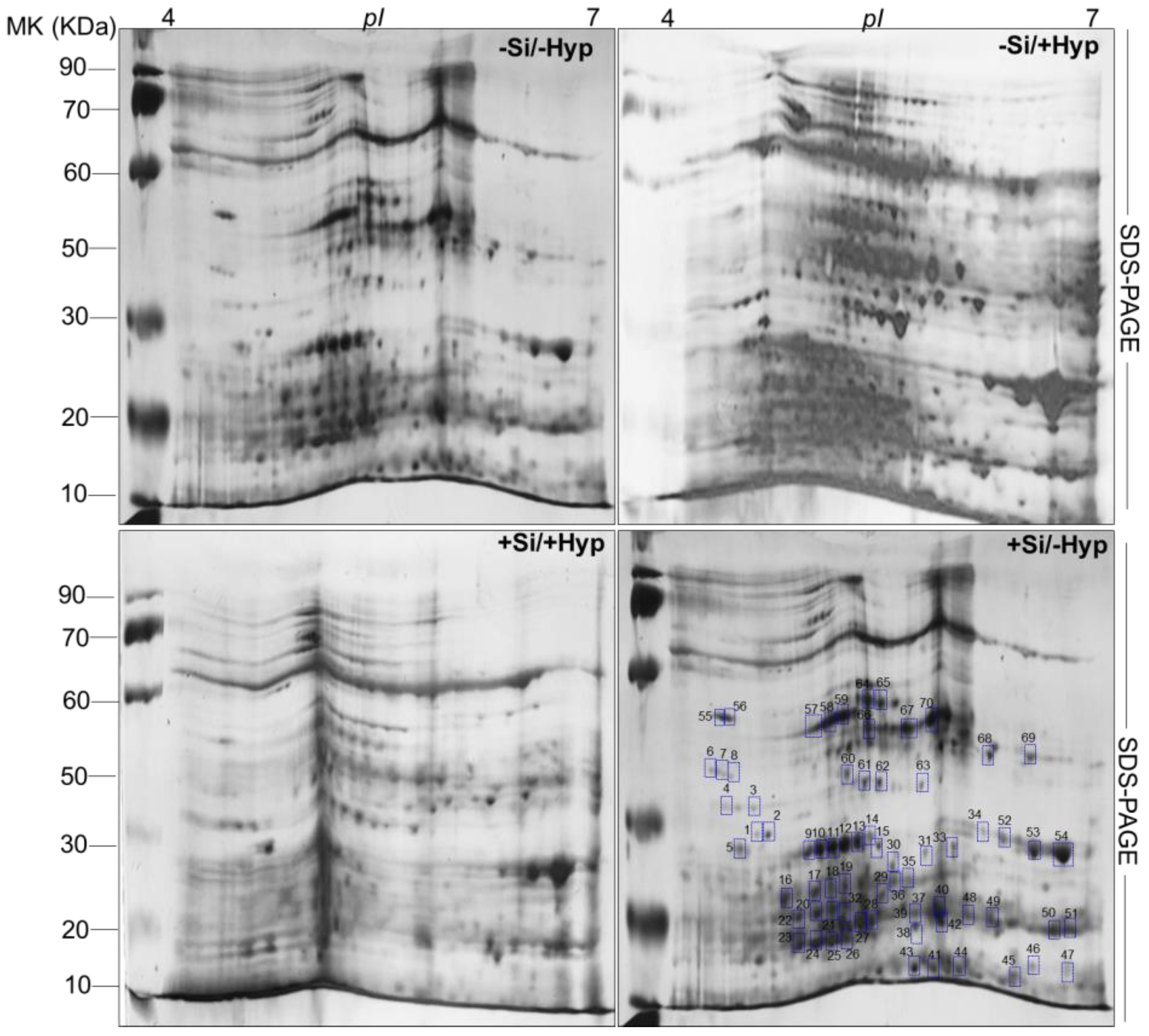
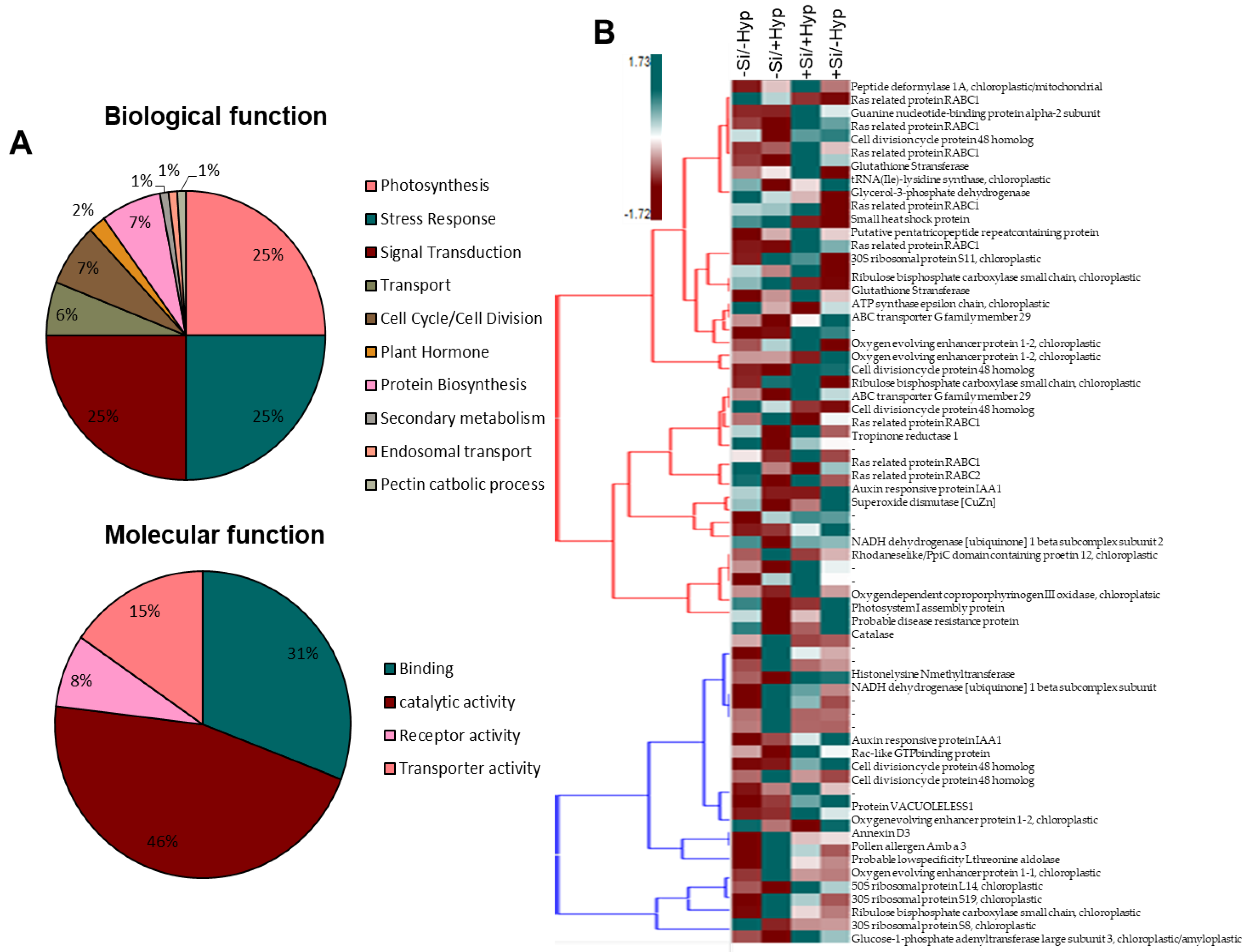
2.3. Carnation Proteome Changes in Response to Hyperhydricity and Si Supply
2.3.1. Proteins Related to Stress Response
2.3.2. Proteins Related to Photosynthesis
2.3.3. Proteins Related to Signal Transduction
2.4. Validation of Selected Differentially Expressed Proteins
2.5. Protein–Protein Interaction Network
2.6. Ultrastructure of Chloroplasts
3. Materials and Methods
3.1. Plant Material, Culture Conditions and Treatments
3.2. Si-Concentration Determination
3.3. DAB and NBT Staining
3.4. Protein Sample Preparation for 2-DE
3.5. Two-Dimensional Gel Electrophoresis (2-DE) and Staining
3.6. Image and Data Analysis
3.7. Protein in Gel Digestion
3.8. Protein Identification Using MALDI-TOF MS and MS/MS Analysis
3.9. Protein Functional Classifications and Hierarchical Clustering
3.10. Western Blots (Immunoblots)
3.11. Protein–Protein Interactions
3.12. Ultrastructure of Chloroplasts
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ali, A.; Afrasiab, H.; Naz, S.; Rauf, M.; Iqbal, J. An efficient protocol for in vitro propagation of carnation (Dianthus caryophyllus L.). Pak. J. Bot. 2008, 40, 111–121. [Google Scholar]
- Kanwar, J.K.; Kumar, S. Influence of growth regulators and explants on shoot regeneration in carnation. Hortic. Sci. 2009, 4, 140–146. [Google Scholar]
- Altvorst, A.G.; Koehorst, H.J.J.; Bruinsma, T.; Jansen, J.; Custers, J.B.M.; Jong, J.; Dons, J.J.M. Adventitious shoot formation from in vitro leaf explants of carnation (Dianthus caryophyllus L.). Sci. Hortic. 1992, 51, 223–235. [Google Scholar] [CrossRef]
- Brar, M.S.; AlKhayri, J.M.; Klingaman, G.L. In vitro shoot multiplication of carnation axillary buds and nodes. Vitro 1995, 31, 61A. [Google Scholar]
- Olmos, E.; Hellín, E. Ultrastructural differences of hyperhydric and normal leaves from regenerated carnation plants. Sci. Hortic. 1998, 75, 91–101. [Google Scholar] [CrossRef]
- Correl, M.J.; Wu, Y.; Weathers, P.J. Controlling hyperhydration of carnations (Dianthus caryophyllus L.) grown in a mist reactor. Biotechnol. Bioeng. 2001, 71, 307–314. [Google Scholar] [CrossRef]
- Debergh, P.; Aitken-Christie, J.; Cohen, D.; Grout, B.; von Arnold, S.; Zimmerman, R.H.; Ziv, M. Reconsideration of the term ‘vitrification’ as used in micropropagation. Plant Cell Tissue Organ Cult. 1992, 30, 135–140. [Google Scholar] [CrossRef]
- Saher, S.; Piqueras, A.; Hellin, E.; Olmos, E. Prevention of hyperhydricity in micropropagated carnation shoots by bottom cooling: Implications of oxidative stress. Plant Cell Tissue Organ Cult. 2005, 81, 149–158. [Google Scholar] [CrossRef]
- Muneer, S.; Soundararajan, P.; Jeong, B.R. Proteomic and antioxidant analysis elucidates the underlying mechanism of tolerance to hyperhydricity stress in in vitro shoot cultures of Dianthus caryophyllus. J. Plant Growth Regul. 2016, 35, 667–679. [Google Scholar] [CrossRef]
- Picoli, E.A.T.; Otoni, W.C.; Figueira, M.L.; Carolino, S.M.B.; Almeida, R.S.; Silva, E.A.M.; Carvalho, C.R.; Fontes, E.P.B. Hyperhydricity in in vitro eggplant regenerated plants: Structural characteristics and involvement of BiP (binding protein). Plant Sci. 2001, 160, 857–868. [Google Scholar] [CrossRef]
- Franck, T.; Kevers, C.; Gaspar, T. Protective enzymatic systems against activated oxygen species compared in normal and vitrified shoots of Prunus avium L. raised in vitro. Plant Growth Regul. 1995, 16, 253–256. [Google Scholar] [CrossRef]
- Jausoro, V.; Llorente, B.E.; Apóstolo, N.M. Structural differences between hyperhydric and normal in vitro shoots of Handroanthus impetiginosus (Mart. ex DC) Mattos (Bignoniaceae). Plant Cell Tissue Organ Cult. 2010, 101, 183–191. [Google Scholar] [CrossRef]
- Kevers, C.; Coumans, M.; Coumans-Gilles, F.; Gaspar, T. Physiological and biochemical events leading to vitrification or shoots cultured in vitro. Physiol. Plant. 1984, 61, 69–74. [Google Scholar] [CrossRef]
- Fontes, M.A.; Otoni, W.C.; Carolino, S.M.B.; Brommonschenkel, S.H.; Fontes, E.P.B.; Fári, M.; Louro, R.P. Hyperhydricity in pepper plants regenerated in vitro: Involvement of BiP (Binding protein) and ultrastructural aspects. Plant Cell Rep. 1999, 19, 81–87. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Park, S.Y.; Ali, M.B.; Shin, K.S.; Paek, K.Y. Hyperhydricity in apple: Ultrastructural and physiological aspects. Tree Physiol. 2005, 26, 377–388. [Google Scholar] [CrossRef]
- Lai, C.C.; Lin, H.M.; Nalawade, S.M.; Fang, W.; Tsay, H.S. Hyperhydricity in shoot cultures of Scrophularia yoshimurae can be effectively reduced by ventilation of culture vessels. J. Plant Physiol. 2005, 162, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Dewir, Y.H.; Chakrabarty, D.; Ali, M.B.; Hahn, E.J.; Paek, K.Y. Lipid peroxidation and antioxidant enzyme activities of Euphorbia millii hyperhydric shoots. Environ. Exp. Bot. 2006, 58, 93–99. [Google Scholar] [CrossRef]
- Balen, B.; Tkalec, M.; Pavoković, D.; Pevalek-Kozlina, B.; Krsnik-Rasol, M. Growth conditions in in vitro culture can induce oxidative stress in Mammillaria gracilis tissues. J. Plant Growth Regul. 2009, 28, 36–45. [Google Scholar] [CrossRef]
- Van den Dries, N.; Giannì, S.; Czerednik, A.; Krens, F.A.; de Klerk, G.J. Flooding of the apoplast is a key factor in the development of hyperhydricity. J. Exp. Bot. 2013, 64, 5221–5230. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Jiang, F.; Wu, Z. The apoplastic oxidative burst as a key factor of hyperhydricity in garlic plantlet in vitro. Plant Cell Tissue Organ Cult. 2014, 120, 571–584. [Google Scholar] [CrossRef]
- Wu, Z.; Chen, L.J.; Long, Y.J. Analysis of ultrastructure and reactive oxygen species of hyperhydric garlic (Allium sativum L.) shoot. In Vitro Cell. Dev. Biol. Plant 2009, 45, 483–490. [Google Scholar] [CrossRef]
- Muneer, S.; Kim, T.H.; Qureshi, M.I. Fe modulates Cd-induced oxidative stress and the expression of stress responsive proteins in the nodules of Vigna radiata. Plant Growth Regul. 2012, 68, 421–433. [Google Scholar] [CrossRef]
- Muneer, S.; Jeong, B.R. Proteomic analysis of salt-stress responsive proteins in roots of tomato (Lycopersicon esculentum L.) plants towards silicon efficiency. Plant Growth Regul. 2015, 77, 133–146. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Ann. Rev. Plant Phyiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.G.; Manivannan, A.; Soundararajan, P.; Jeong, B.R. Physiological and proteomic analysis in chloroplasts of Solanum lycopersicum L, under silicon efficiency and salinity stress. Int. J. Mol. Sci. 2014, 15, 21803–21824. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of silicon as an essential element for higher plants. Comments Agric. Food Chem. 1990, 2, 99–122. [Google Scholar]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Savvas, D.; Giotis, D.; Chatzieustratiou, E.; Bakea, M.; Patakioutas, G. Silicon supply in soiless cultivation of zucchini alleviates stress induced by salinity and powdery mildew infections. Environ. Exp. Bot. 2009, 35, 633–639. [Google Scholar]
- Neumann, D.; Nieden, U. Silicon and heavy metal tolerance of higher plants. Phytochemistry 2001, 56, 685–692. [Google Scholar] [CrossRef]
- Nwugo, C.C.; Huerta, A.J. Effects of silicone nutrition on cadmium uptake, growth and photosynthesis of rice plants exposed to low-level cadmium. Plant Soil 2008, 311, 73–86. [Google Scholar] [CrossRef]
- Pontigo, S.; Godoy, K.; Jiménez, H.; Gutiérrez-Moraga, A.; Mora, M.D.L.L.; Cartes, P. Silicon-mediated alleviation of aluminum toxicity by modulation of Al/Si uptake and antioxidant performance in ryegrass plants. Front. Plant Sci. 2017, 8, 642. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultivar and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Kumar, D.; Chauhan, D.K. Impact of exogenous silicon addition on chromium uptake, growth, mineral elements, oxidative stress, antioxidant capacity, and leaf and root structures in rice seedlings exposed to hexavalent chromium. Acta Physiol. Plant. 2012, 34, 279–289. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Kumar, D.; Chauhan, D.K. Rice seedlings under cadmium stress: Effect of silicon on growth, cadmium uptake, oxidative stress, antioxidant capacity and root and leaf structures. Chem. Ecol. 2012, 28, 281–291. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K.; Rai, A.K. Silicon-mediated alleviation of Cr (VI) toxicity in wheat seedlings as evidenced by chlorophyll florescence, laser induced breakdown spectroscopy and anatomical changes. Ecotoxicol. Environ. Saf. 2015, 113, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxová, M.; Lux, A. Application of silicone enhanced drought tolerance in Sorghum bicolor. Physiol. Plant. 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Jeong, B.R. Silicon decreases Fe deficiency responses by improving photosynthesis and maintaining composition of thylakoid multiprotein complex proteins in soybean plants (Glycine max L.). J. Plant Growth Regul. 2015, 34, 485–498. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Afzal, R.M.; Ahmad, R.; Mujeeb, F.; Sarwar, A.; Ali, L. Alleviation of detrimental effects of NaCl by silicon nutrition in salt-sensitive and salt-tolerant genotypes of sugarcane (Saccharum officinarum L.). Plant Soil 2010, 326, 381–391. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan; Elsevier Science: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Ma, J.F. Silicon transporters in higher plants. In MIPs and Their Role in the Exchange of Metalloids; Jahn, T.P., Bienert, G.P., Eds.; Lands Bioscience and Springer Science; Business Media: Austin, New York, NY, USA, 2010; pp. 99–109. [Google Scholar]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.A. Cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Alvard, D.; Cote, F.; Teisson, C. Comparison of methods of liquid medium culture for banana micropropagation. Plant Cell Tissue Organ Cult. 1993, 32, 55–60. [Google Scholar] [CrossRef]
- Snyman, S.J.; Nkwanyana, P.D.; Watt, M.P. Alleviation of hyperhydricity of sugarcane plantlets produced in RITA® vessels and genotypic and phenotypic characterization of acclimated plants. S. Afr. J. Bot. 2011, 77, 685–692. [Google Scholar] [CrossRef]
- Muneer, S.; Park, Y.; Jeong, B.R. Red and blue light emitting diodes (LEDs) participate in mitigation of hyperhydricity in in vitro grown carnation genotypes (Dianthus Caryophyllus). J. Plant Growth Regul. 2017. [Google Scholar] [CrossRef]
- Gaspar, T.; Kevers, C.; Franck, T.; Bisbis, B.; Billard, J.P.; Huault, C.; Le Dily, F.; Petit-Paly, G.; Rideau, M.; Penel, C.; et al. Paradoxical results in the results of hyperhydric tissues considered as being under stress: Questions for a debate. Bulg. J. Plant Physiol. 1995, 21, 80–97. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Breusegem, F.V. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Clephan, A.L.; Davies, W.J. Rapid low temperature-induced stomatal closure occurs in cold-tolerant Commelina communis leaves but not in cold-sensitive tobacco leaves, via a mechanism that involves apoplastic calcium but not abscisic acid. Plant Physiol. 2001, 126, 1566–1578. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Saibo, N.J.M.; Lourenço, T.; Oliveira, M.M. Transcription factors and regulation of photosynthetic and related metabolism under environmental stresses. Ann. Bot. 2009, 103, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Giraudat, J.; Schroeder, J.I. Cell signaling and gene regulation—Plant signal transduction pathways: Graying of the black boxes—Editorial overview. Curr. Opin. Plant Biol. 2001, 4, 379–381. [Google Scholar] [CrossRef]
- Ruggiero, B.; Koiwa, H.; Manabe, Y.; Quist, T.M.; Inan, G.; Saccardo, F.; Joly, R.J.; Hasegawa, P.M.; Bressan, R.A.; Maggio, A. Uncoupling the effects of abscisic acid on plant growth and water relations. Analysis of sto1/nced3, an abscisic acid-deficient but salt stress-tolerant mutant in Arabidopsis. Plant Physiol. 2004, 136, 3134–3147. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, X.; Yang, S.; Zhou, Y.; Dong, C.; Ren, J.; Sun, X.; Yang, Y. Comparative physiological and proteomic analysis reveals the leaf response to cadmium-induced stress in poplar (Populus yunnanensis). PLoS ONE 2015, 10, e0137396. [Google Scholar] [CrossRef] [PubMed]
- Dixit, G.; Singh, A.P.; Kumar, A.; Dwivedi, S.; Deeba, F.; Kumar, S.; Suman, S.; Adhikari, B.; Shukla, Y.; Trivedi, P.K.; et al. Sulfur alleviates arsenic toxicity by reducing its accumulation and modulating proteome, amino acids and thiol metabolism in rice leaves. Sci. Rep. 2015, 5, 16205. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Ko, C.H.; Wei, H.; Chen, Y.; Jeong, B.R. Physiological and proteomic investigations to study the response of tomato graft unions under temperature stress. PLoS ONE 2016, 11, e0157439. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Timperio, A.M.; D’Amici, G.M.; Barta, C.; Loreto, F.; Zolla, L. Proteomic, pigment composition, and organization of thylakoid membrane in iron deficient-plant spinach leaves. J. Exp. Bot. 2007, 58, 3695–3710. [Google Scholar] [CrossRef] [PubMed]
- Fagioni, M.; D’Amici, G.M.; Timperio, A.M.; Zolla, L. Proteomic analysis of multiprotein complexes in the thylakoids membrane upon cadmium treatment. J. Proteome Res. 2009, 8, 310–326. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Manivannan, A.; Cho, Y.S.; Jeong, B.R. Exogenous supplementation of silicon improved the recovery of hyperhydric shoots in Dianthus caryophyllus L. by stabilizing the physiology and protein expression. Front. Plant Sci. 2017, 8, 738. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F.A. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Protein Name | Plant Species | Accession Number | Mr Value | Calcu. pI/Exp. pI | Sequence Coverage |
|---|---|---|---|---|---|---|
| Stress Response | ||||||
| 11 | Small heat shock protein | Triticum aestivum | HS21C_WHEAT | 26579 | 9.6/6.1 | 28 |
| 12 | Putative pentatricopeptide repeat containing protein | Arabidopsis thaliana | PPR7_ARATH | 47546 | 6.8/6.2 | 17 |
| 17 | Glutathione S-transferase | Arabidopsis thaliana | GSTU5_ARATH | 25984 | 5.4/5.9 | 31 |
| 33 | Superoxide dismutase (CuZn) | Picea abies | SODC2_PICAB | 2230 | 8.3/6.5 | 100 |
| 42 | Probable disease resistance protein | Arabidopsis thaliana | DRL24_ARATH | 81866 | 7.7/6.5 | 21 |
| 43 | Catalase | Solanum melongena | CATA_SOLME | 56584 | 6.8/6.6 | 26 |
| 58 | Annexin D3 | Arabidopsis thaliana | ANXD3_ARATH | 36233 | 6.0/5.8 | 21 |
| Photosynthesis | ||||||
| 16 | Ribulose bisphosphate carboxylase small chain, chloroplastic | Chlamydomonas moewusii | RBS_CHLMO | 18896 | 9.4/5.3 | 29 |
| 18 | ATP synthase epsilon chain, chloroplastic | Pinus koraiensis | ATPE_PINKO | 15118 | 8.8/6.1 | 36 |
| 21 | Oxygen evolving enhancer protein 1-2, chloroplastic | Arabidopsis thaliana | PSBO2_ARATH | 34998 | 5.9/6.1 | 21 |
| 22 | Oxygen evolving enhancer protein 1-2, chloroplastic | Arabidopsis thaliana | PSBO2_ARATH | 34998 | 5.9/5.3 | 21 |
| 24 | Ribulose bisphosphate carboxylase small chain, chloroplastic | Chlamydomonas moewusii | RBS_CHLMO | 18896 | 9.4/6.0 | 42 |
| 37 | Rhodanese like/PpiC domain containing protein 12, chloroplastic | Arabidopsis thaliana | STR12_ARATH | 32959 | 8.6/6.6 | 31 |
| 40 | Oxygen dependent coproporphyrinogen III oxidase, chloroplastic | Hordeum vulgare | HEM6_HORVU | 43529 | 8.0/6.5 | 48 |
| 41 | Photosystem I assembly protein | Chaetosphaeridium globosum | YCF4_CHAGL | 21209 | 9.6/6.4 | 28 |
| 57 | Oxygen evolving enhancer protein 1-2, chloroplastic | Arabidopsis thaliana | PSBO2_ARATH | 34998 | 5.9/4.3 | 21 |
| 61 | Oxygen evolving enhancer protein 1-1, chloroplastic | Arabidopsis thaliana | PSBO1_ARATH | 35121 | 5.5/5.5 | 37 |
| 67 | Ribulose bisphosphate carboxylase small chain, chloroplastic | Chlamydomonas moewusii | RBS_CHLMO | 18896 | 9.4/5.7 | 36 |
| 70 | Glucose-1-phosphate adenyl transferase large subunit 3, chloroplastic/amyloplastic | Solanum tuberosum | GLGL3_SOLTU | 53569 | 8.9/5.8 | 24 |
| Signal transduction | ||||||
| 1 | Peptide deformylase 1A, chloroplastic/mitochondrial | Arabidopsis thaliana | DEF1A_ARATH | 29977 | 8.6/5.0 | 37 |
| 2 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/5.2 | 30 |
| 3 | Guanine nucleotide-binding protein α-2 subunit | Pisum sativum | GPA2_PEA | 44641 | 5.81/5.0 | 25 |
| 4 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/4.8 | 30 |
| 6 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/4.9 | 36 |
| 7 | Glutathione S-transferase | Arabidopsis thaliana | GSTU5_ARATH | 25984 | 5.4/4.8 | 36 |
| 10 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/6.0 | 30 |
| 13 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/6.3 | 30 |
| 27 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/6.3 | 31 |
| 30 | Ras related protein RABC1 | Arabidopsis thaliana | RABC1_ARATH | 23516 | 5.6/6.3 | 31 |
| 31 | Ras related protein RABC2 | Arabidopsis thaliana | RABC1_ARATH | 23517 | 5.6/6.4 | 32 |
| Cell cycle/Cell division | ||||||
| 5 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | 89275 | 5.0/4.9 | 15 |
| 23 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | 89275 | 5.0/5.9 | 17 |
| 26 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | 89275 | 5.0/6.2 | 15 |
| 53 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | 89275 | 5.0/6.8 | 18 |
| 54 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | 89275 | 5.0/6.9 | 18 |
| Transport | ||||||
| 19 | ABC transporter G family member 29 | Arabidopsis thaliana | AB29G_ARATH | 160195 | 8.5/6.2 | 9 |
| 25 | ABC transporter G family member 29 | Arabidopsis thaliana | AB29G_ARATH | 160195 | 8.5/5.3 | 13 |
| 36 | NADH dehydrogenase [ubiquinone] 1 β subcomplex subunit 2 | Arabidopsis thaliana | NDUB2_ARATH | 7563 | 8.9/6.3 | 69 |
| 47 | NADH dehydrogenase [ubiquinone] 1 β subcomplex subunit | Arabidopsis thaliana | NDUB2_ARATH | 7563 | 8.9/7.0 | 52 |
| Plant hormone | ||||||
| 32 | Auxin responsive protein IAA1 | Arabidopsis thaliana | IAA1_ARATH | 19019 | 7.6/6.5 | 40 |
| 51 | Auxin responsive protein IAA1 | Arabidopsis thaliana | IAA1_ARATH | 19019 | 7.6/7.0 | 40 |
| Protein biosynthesis | ||||||
| 8 | tRNA(Ile)-lysidine synthase, chloroplastic | Zygnema circumcarinatum | TILS_ZYGCR | 48914 | 9.8/4.9 | 20 |
| 14 | 30S ribosomal protein S11, chloroplastic | Gnetum parvifolium | RR11_GNEPA | 14467 | 11.0/6.4 | 35 |
| 65 | 50S ribosomal protein L14, chloroplastic | Brachypodium distachyon | RK14_BRADI | 13586 | 8.5/5.6 | 50 |
| 66 | 30S ribosomal protein S19, chloroplastic | Platanus occidentalis | RR19_PLAOC | 10593 | 10.7/6.4 | 55 |
| 69 | 30S ribosomal protein S8, chloroplastic | Acutodesmus obliquus | RR8_ACUOB | 15189 | 9.9/6.7 | 38 |
| Secondary metabolism | ||||||
| 9 | Glycerol-3-phosphate dehydrogenase | Oryza sativa | GPDA_ORYSJ | 46733 | 9.7/5.9 | 18 |
| 28 | Tropinone reductase 1 | Datura stramonium | TRN1_DATST | 29598 | 6.1/6.4 | 18 |
| 46 | Histone lysine N methyltransferase | Arabidopsis thaliana | ATXR4_ARATH | 36111 | 6.3/6.8 | 35 |
| 52 | Rac-like GTPbinding protein | Arabidopsis thaliana | RAC10_ARATH | 23863 | 7.5/6.5 | 53 |
| 60 | Probable low specificity l-threonine aldolase | Arabidopsis thaliana | THA1_ARATH | 38917 | 6.9/5.6 | 42 |
| Endosomal Transport | ||||||
| 56 | Protein VACUOLELESS1 | Arabidopsis thaliana | VCL1_ARATH | 96570 | 5.5/4.2 | 24 |
| Pectin catabolic process | ||||||
| 59 | Pollen allergen Amb a 3 | Ambrosia artemisiifolia | MPAA3_AMBEL | 11368 | 6.1/5.6 | 59 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muneer, S.; Wei, H.; Park, Y.G.; Jeong, H.K.; Jeong, B.R. Proteomic Analysis Reveals the Dynamic Role of Silicon in Alleviation of Hyperhydricity in Carnation Grown In Vitro. Int. J. Mol. Sci. 2018, 19, 50. https://doi.org/10.3390/ijms19010050
Muneer S, Wei H, Park YG, Jeong HK, Jeong BR. Proteomic Analysis Reveals the Dynamic Role of Silicon in Alleviation of Hyperhydricity in Carnation Grown In Vitro. International Journal of Molecular Sciences. 2018; 19(1):50. https://doi.org/10.3390/ijms19010050
Chicago/Turabian StyleMuneer, Sowbiya, Hao Wei, Yoo Gyeong Park, Hai Kyoung Jeong, and Byoung Ryong Jeong. 2018. "Proteomic Analysis Reveals the Dynamic Role of Silicon in Alleviation of Hyperhydricity in Carnation Grown In Vitro" International Journal of Molecular Sciences 19, no. 1: 50. https://doi.org/10.3390/ijms19010050
APA StyleMuneer, S., Wei, H., Park, Y. G., Jeong, H. K., & Jeong, B. R. (2018). Proteomic Analysis Reveals the Dynamic Role of Silicon in Alleviation of Hyperhydricity in Carnation Grown In Vitro. International Journal of Molecular Sciences, 19(1), 50. https://doi.org/10.3390/ijms19010050