Immunological Heterogeneity of Healthy Peripheral Blood Stem Cell Donors—Effects of Granulocyte Colony-Stimulating Factor on Inflammatory Responses

Abstract
1. Introduction
2. Results
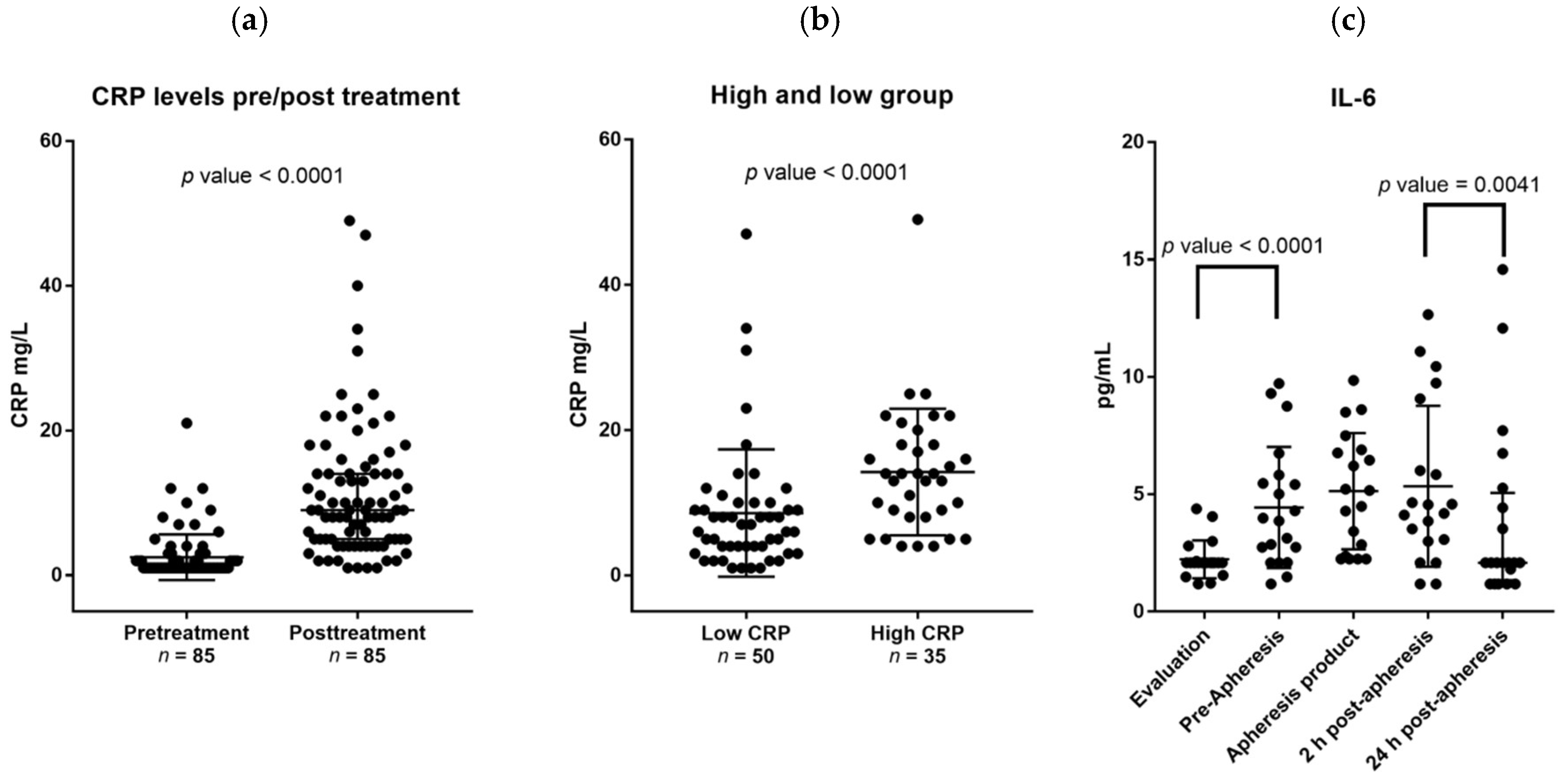
2.1. Healthy Stem Cell Donors Are Heterogeneous with Regard to Ongoing Acute Phase Reaction and the G-CSF Therapy Causes a Further Increase of CRP Levels for a Subset of Donors
2.2. G-CSF Therapy of Healthy Stem Cell Donors Is Associated with Increased Serum Levels of IL-6 Whereas the Levels of Other IL-6 Family Members Are Not Altered during Stem Cell Mobilization
2.3. CRP Levels during G-CSF Therapy Are Significantly Correlated with the Oncostatin M Serum Levels but There Is No Association with the Corresponding Serum IL-6 Levels
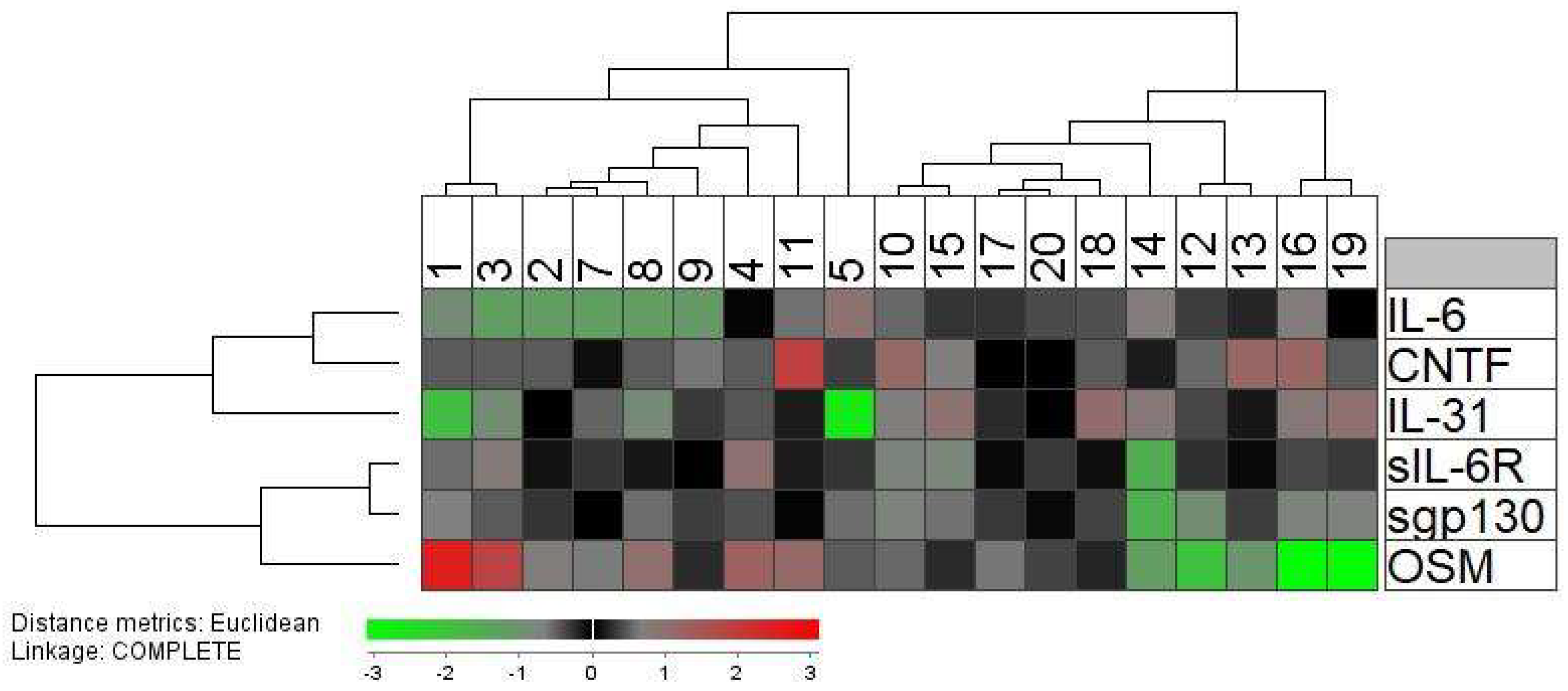
2.4. Systemic (Serum) Levels of IL-6 Family Cytokines and Especially the Oncostatin M Levels Vary between Donors Both When Tested before and during G-CSF Therapy
2.5. The Levels of Immunocompetent Cell Subsets in Peripheral Blood and Allogeneic Stem Cell Grafts Vary between Healthy Donors: Studies of Associations between Serum Levels of IL6 Family Cytokines, Circulating Immunocompetent Cells, and Graft Content of Immunocompetent Cells
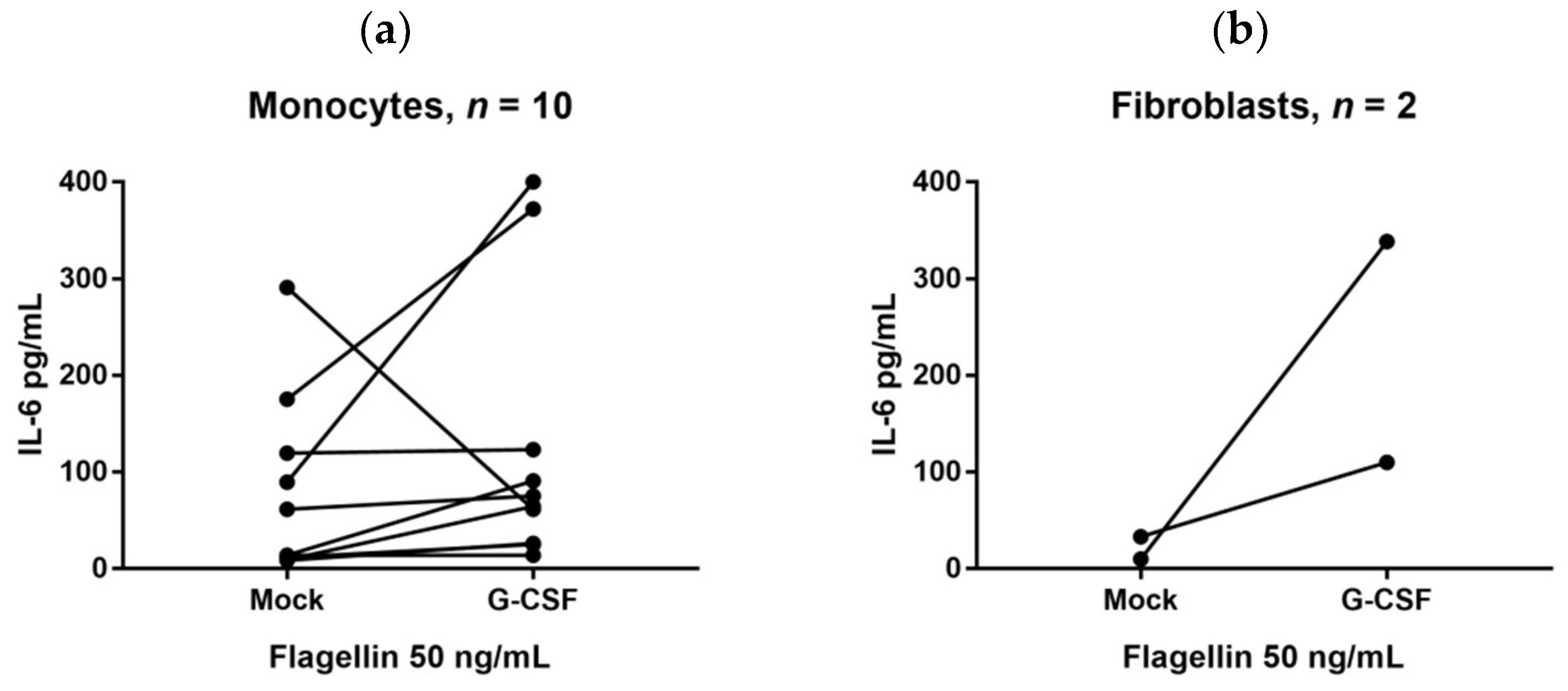
2.6. G-CSF Can Modulate IL-6 Release by Immunocompetent and Mesenchymal Cells
3. Discussion
4. Material and Methods
4.1. Patient Studies and Donor Samples
4.2. Flow Cytometric Analysis
4.3. In Vitro Culture of Monocytes and Fibroblasts
4.4. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALLO-SCT | Allogeneic stem cell transplantation |
| CNTF | Ciliary neutrophilic factor |
| gp130 | Glycoprotein 130 |
| GVHD | Graft-versus-host disease |
| G-CSF | Granulocyte-colony stimulating factor |
| IL-31 | Interleukin-31 |
| IL-6 | Interleukin-6 |
| LPS | Lipopolysaccharide |
| MCSs | Mesenchymal stem cells |
| OSM | Oncostatin M |
| sIL-6R | Soluble IL-6 receptor |
| TLR | Toll-like receptor. |
References
- Ballen, K.K.; King, R.J.; Chitphakdithai, P.; Bolan, C.D., Jr.; Agura, E.; Hartzman, R.J.; Kernan, N.A. The national marrow donor program 20 years of unrelated donor hematopoietic cell transplantation. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2008, 14, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Shaw, B.E.; Confer, D.L.; Hwang, W.; Pulsipher, M.A. A review of the genetic and long-term effects of G-CSF injections in healthy donors: A reassuring lack of evidence for the development of haematological malignancies. Bone Marrow Transplant. 2015, 50, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Boneberg, E.M.; Hareng, L.; Gantner, F.; Wendel, A.; Hartung, T. Human monocytes express functional receptors for granulocyte colony-stimulating factor that mediate suppression of monokines and interferon-gamma. Blood 2000, 95, 270–276. [Google Scholar] [PubMed]
- Boneberg, E.M.; Hartung, T. Granulocyte colony-stimulating factor attenuates LPS-stimulated IL-1β release via suppressed processing of proIL-1β, whereas TNF-α release is inhibited on the level of proTNF-α formation. Eur. J. Immunol. 2002, 32, 1717–1725. [Google Scholar] [CrossRef]
- Sloand, E.M.; Kim, S.; Maciejewski, J.P.; van Rhee, F.; Chaudhuri, A.; Barrett, J.; Young, N.S. Pharmacologic doses of granulocyte colony-stimulating factor affect cytokine production by lymphocytes in vitro and in vivo. Blood 2000, 95, 2269–2274. [Google Scholar] [PubMed]
- Hartung, T.; Docke, W.D.; Gantner, F.; Krieger, G.; Sauer, A.; Stevens, P.; Volk, H.D.; Wendel, A. Effect of granulocyte colony-stimulating factor treatment on ex vivo blood cytokine response in human volunteers. Blood 1995, 85, 2482–2489. [Google Scholar] [PubMed]
- Rutella, S.; Lemoli, R.M. Regulatory T cells and tolerogenic dendritic cells: From basic biology to clinical applications. Immunol. Lett. 2004, 94, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Rutella, S.; Zavala, F.; Danese, S.; Kared, H.; Leone, G. Granulocyte colony-stimulating factor: A novel mediator of T cell tolerance. J. Immunol. 2005, 175, 7085–7091. [Google Scholar] [CrossRef] [PubMed]
- Lysak, D.; Hrabetova, M.; Vrzalova, J.; Koza, V.; Navratilova, J.; Svoboda, T.; Jungova, A.; Topolcan, O. Changes of cytokine levels during granulocyte-colony-stimulating factor stem cell mobilization in healthy donors: Association with mobilization efficiency and potential predictive significance. Transfusion 2011, 51, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Czerw, T.; Labopin, M.; Schmid, C.; Cornelissen, J.J.; Chevallier, P.; Blaise, D.; Kuball, J.; Vigouroux, S.; Garban, F.; Lioure, B.; et al. High CD3+ and CD34+ peripheral blood stem cell grafts content is associated with increased risk of graft-versus-host disease without beneficial effect on disease control after reduced-intensity conditioning allogeneic transplantation from matched unrelated donors for acute myeloid leukemia—An analysis from the acute leukemia working party of the european society for blood and marrow transplantation. Oncotarget 2016, 7, 27255–27266. [Google Scholar] [CrossRef] [PubMed]
- Pulsipher, M.A.; Chitphakdithai, P.; Logan, B.R.; Shaw, B.E.; Wingard, J.R.; Lazarus, H.M.; Waller, E.K.; Seftel, M.; Stroncek, D.F.; Lopez, A.M.; et al. Acute toxicities of unrelated bone marrow versus peripheral blood stem cell donation: Results of a prospective trial from the national marrow donor program. Blood 2013, 121, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.S.; Wagle, M.; Matous, S.; Swanson, R.S.; Pihan, G.; Lowry, P.A.; Stewart, F.M.; Heard, S.O. Spontaneous splenic rupture following administration of granulocyte colony-stimulating factor (G-CSF): Occurrence in an allogeneic donor of peripheral blood stem cells. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 1997, 3, 45–49. [Google Scholar]
- Hatfield, K.J.; Melve, G.K.; Bruserud, O. Granulocyte colony-stimulating factor alters the systemic metabolomic profile in healthy donors. Metabolomics 2017, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Kiyokawa, N.; Taguchi, T.; Suzuki, K.; Sekino, T.; Mimori, K.; Suzuki, T.; Nakajima, H.; Katagiri, Y.U.; Fujimura, J.; et al. Granulocyte colony-stimulating factor directly affects human monocytes and modulates cytokine secretion. Exp. Hematol. 2002, 30, 1115–1123. [Google Scholar] [CrossRef]
- Anasetti, C. What are the most important donor and recipient factors affecting the outcome of related and unrelated allogeneic transplantation? Best Pract. Res. Clin. Haematol. 2008, 21, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Bruserud, Ø.; Melve, G.K.; Gedde-Dahl, T.; Tvedt, T.H.A. Immunological heterogeneity of healthy peripheral blood stem cell donors—Preharvesting donor characteristics, additional heterogeneity induced by granulocyte colony-stimulating factor and possible importance for outcome after allotransplantation. Expert Rev. Hematol. 2018, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.J.; Xu, L.P.; Wang, Y.; Zhang, X.H.; Chen, H.; Chen, Y.H.; Wang, F.R.; Han, W.; Sun, Y.Q.; Yan, C.H.; et al. Controlled, randomized, open-label trial of risk-stratified corticosteroid prevention of acute graft-versus-host disease after haploidentical transplantation. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Bruserud, Ø.; Melve, G.K.; Gedde-Dahl, T.; Tvedt, T.H.A. Immunological heterogeneity of healthy peripheral blood stem cell donors induced by granulocyte colony-stimulating factor—Is this important for outcome after allotransplantation? Expert Rev. Hematol. 2018. [Google Scholar] [CrossRef]
- Fidyk, W.; Mitrus, I.; Ciomber, A.; Smagur, A.; Chwieduk, A.; Glowala-Kosinska, M.; Giebel, S. Evaluation of proinflammatory and immunosuppressive cytokines in blood and bone marrow of healthy hematopoietic stem cell donors. Cytokine 2018, 102, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Impola, U.; Larjo, A.; Salmenniemi, U.; Putkonen, M.; Itala-Remes, M.; Partanen, J. Graft immune cell composition associates with clinical outcome of allogeneic hematopoietic stem cell transplantation in patients with aml. Front. Immunol. 2016, 7, 523. [Google Scholar] [CrossRef] [PubMed]
- Melve, G.K.; Ersvaer, E.; Eide, E.E.; Kristoffersen, E.K.; Bruserud, Ø. Peripheral blood stem cell mobilization in healthy donors by granulocyte colony-stimulating factor causes preferential mobilization of lymphocyte subsets. Front. Immunol. 2018, 9, 845. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. IL-6 trans-signaling via the soluble IL-6 receptor: Importance for the pro-inflammatory activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; von Aulock, S.; Zedler, S.; Schinkel, C.; Hartung, T.; Faist, E. Perioperative recombinant human granulocyte colony-stimulating factor (filgrastim) treatment prevents immunoinflammatory dysfunction associated with major surgery. Ann. Surg. 2004, 239, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Baumann, H.; Ziegler, S.F.; Mosley, B.; Morella, K.K.; Pajovic, S.; Gearing, D.P. Reconstitution of the response to leukemia inhibitory factor, oncostatin M, and ciliary neurotrophic factor in hepatoma cells. J. Biol. Chem. 1993, 268, 8414–8417. [Google Scholar] [PubMed]
- Zaucha, J.M.; Gooley, T.; Bensinger, W.I.; Heimfeld, S.; Chauncey, T.R.; Zaucha, R.; Martin, P.J.; Flowers, M.E.; Storek, J.; Georges, G.; et al. CD34 cell dose in granulocyte colony-stimulating factor-mobilized peripheral blood mononuclear cell grafts affects engraftment kinetics and development of extensive chronic graft-versus-host disease after human leukocyte antigen-identical sibling transplantation. Blood 2001, 98, 3221–3227. [Google Scholar] [PubMed]
- Melve, G.K.; Ersvssr, E.; Kittang, A.O.; Bruserud, O. The chemokine system in allogeneic stem-cell transplantation: A possible therapeutic target? Expert Rev. Hematol. 2011, 4, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Tvedt, T.H.A.; Ersvaer, E.; Tveita, A.A.; Bruserud, O. Interleukin-6 in allogeneic stem cell transplantation: Its possible importance for immunoregulation and as a therapeutic target. Front. Immunol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Melve, G.K.; Ersvaer, E.; Paulsen Rye, K.; Bushra Ahmed, A.; Kristoffersen, E.K.; Hervig, T.; Reikvam, H.; Hatfield, K.J.; Bruserud, O. The healthy donor profile of immunoregulatory soluble mediators is altered by stem cell mobilization and apheresis. Cytotherapy 2018, 20, 740–754. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Dalmaroni, M.J.; Gerswhin, M.E.; Adamopoulos, I.E. The critical role of toll-like receptors—From microbial recognition to autoimmunity: A comprehensive review. Autoimmun. Rev. 2016, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, L.; Chen, S. Exogenous or endogenous toll-like receptor ligands: Which is the MVP in tumorigenesis? Cell. Mol. Life Sci. 2012, 69, 935–949. [Google Scholar] [CrossRef] [PubMed]
- Reshef, R.; Huffman, A.P.; Gao, A.; Luskin, M.R.; Frey, N.V.; Gill, S.I.; Hexner, E.O.; Kambayashi, T.; Loren, A.W.; Luger, S.M.; et al. High graft CD8 cell dose predicts improved survival and enables better donor selection in allogeneic stem-cell transplantation with reduced-intensity conditioning. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 2392–2398. [Google Scholar] [CrossRef] [PubMed]
- Danby, R.D.; Zhang, W.; Medd, P.; Littlewood, T.J.; Peniket, A.; Rocha, V.; Roberts, D.J. High proportions of regulatory T cells in pbsc grafts predict improved survival after allogeneic haematopoietic SCT. Bone Marrow Transplant. 2016, 51, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Schoergenhofer, C.; Schwameis, M.; Wohlfarth, P.; Brostjan, C.; Abrams, S.T.; Toh, C.H.; Jilma, B. Granulocyte colony-stimulating factor (G-CSF) increases histone-complexed DNA plasma levels in healthy volunteers. Clin. Exp. Med. 2017, 17, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Spiel, A.O.; Bartko, J.; Schwameis, M.; Firbas, C.; Siller-Matula, J.; Schuetz, M.; Weigl, M.; Jilma, B. Increased platelet aggregation and in vivo platelet activation after granulocyte colony-stimulating factor administration. A randomised controlled trial. Thromb. Haemost. 2011, 105, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Tvedt, T.H.; Lie, S.A.; Reikvam, H.; Rye, K.P.; Lindas, R.; Gedde-Dahl, T.; Ahmed, A.B.; Bruserud, O. Pretransplant levels of CRP and interleukin-6 family cytokines; effects on outcome after allogeneic stem cell transplantation. Int. J. Mol. Sci. 2016, 17, 1823. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Kwon, Y.W.; Cho, H.J.; Shin, J.I.; Kim, Y.J.; Lee, S.E.; Youn, S.W.; Lee, H.C.; Kang, H.J.; Shaul, P.W.; et al. G-CSF exerts dual effects on endothelial cells—Opposing actions of direct enos induction versus indirect CRP elevation. J. Mol. Cell. Cardiol. 2008, 45, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Yap, S.H.; Moshage, H.J.; Hazenberg, B.P.; Roelofs, H.M.; Bijzet, J.; Limburg, P.C.; Aarden, L.A.; van Rijswijk, M.H. Tumor necrosis factor (TNF) inhibits interleukin (IL)-1 and/or IL-6 stimulated synthesis of c-reactive protein (CRP) and serum amyloid a (SAA) in primary cultures of human hepatocytes. Biochim. Biophys. Acta 1991, 1091, 405–408. [Google Scholar]
- Garbers, C.; Monhasery, N.; Aparicio-Siegmund, S.; Lokau, J.; Baran, P.; Nowell, M.A.; Jones, S.A.; Rose-John, S.; Scheller, J. The interleukin-6 receptor Asp358Ala single nucleotide polymorphism rs2228145 confers increased proteolytic conversion rates by adam proteases. Biochim. Biophys Acta 2014, 1842, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Mackiewicz, A.; Schooltink, H.; Heinrich, P.C.; Rose-John, S. Complex of soluble human IL-6-receptor/IL-6 up-regulates expression of acute-phase proteins. J. Immunol. 1992, 149, 2021–2027. [Google Scholar] [PubMed]
- Hermanns, H.M. Oncostatin m and interleukin-31: Cytokines, receptors, signal transduction and physiology. Cytokine Growth Factor Rev. 2015, 26, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Pothoven, K.L.; Schleimer, R.P. The barrier hypothesis and oncostatin M: Restoration of epithelial barrier function as a novel therapeutic strategy for the treatment of type 2 inflammatory disease. Tissue Barriers 2017, 5, e1341367. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.D. The enigmatic cytokine oncostatin m and roles in disease. ISRN Inflamm. 2013, 2013, 512103. [Google Scholar] [CrossRef] [PubMed]
- Janssens, K.; Slaets, H.; Hellings, N. Immunomodulatory properties of the IL-6 cytokine family in multiple sclerosis. Ann. N. Y. Acad. Sci. 2015, 1351, 52–60. [Google Scholar]
- Wahl, A.F.; Wallace, P.M. Oncostatin m in the anti-inflammatory response. Ann. Rheum. Dis. 2001, 60, iii75–iii80. [Google Scholar] [PubMed]
- Knorr, D.A.; Bachanova, V.; Verneris, M.R.; Miller, J.S. Clinical utility of natural killer cells in cancer therapy and transplantation. Semin. Immunol. 2014, 26, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Clausen, J.; Petzer, A.L.; Vergeiner, B.; Enk, M.; Stauder, R.; Gastl, G.; Gunsilius, E. Optimal timing for the collection and in vitro expansion of cytotoxic CD56+ lymphocytes from patients undergoing autologous peripheral blood stem cell transplantation. J. Hematother. Stem Cell Res. 2001, 10, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Clausen, J.; Enk, M.; Vergeiner, B.; Eisendle, K.; Petzer, A.L.; Gastl, G.; Gunsilius, E. Suppression of natural killer cells in the presence of CD34+ blood progenitor cells and peripheral blood lymphocytes. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2004, 10, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; DiPietro, L.A. Toll-like receptor function in acute wounds. Adv. Wound Care 2017, 6, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Worel, N.; Buser, A.; Greinix, H.T.; Hagglund, H.; Navarro, W.; Pulsipher, M.A.; Nicoloso de Faveri, G.; Bengtsson, M.; Billen, A.; Espino, G.; et al. Suitability criteria for adult related donors: A consensus statement from the worldwide network for blood and marrow transplantation standing committee on donor issues. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2015, 21, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Liseth, K.; Ersvaer, E.; Abrahamsen, J.F.; Nesthus, I.; Ryningen, A.; Bruserud, O. Long-term cryopreservation of autologous stem cell grafts: A clinical and experimental study of hematopoietic and immunocompetent cells. Transfusion 2009, 49, 1709–1719. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Covariate | Univariate | Multivariate | ||||
|---|---|---|---|---|---|---|
| Coefficient | SE 1 | p-Value | Coefficient | SE 1 | p-Value | |
| Pre G-CSF CRP level | 1.48 | 0.31 | <0.01 | 1.40 | 0.32 | <0.01 |
| Age 2 | 5.30 | 2.45 | 0.03 | 2.16 | 2.39 | 0.37 |
| Mediator | Before G-CSF | During G-CSF (Pre-apheresis) | Graft Supernatant | 2 h after Apheresis | 24 h after Apheresis |
|---|---|---|---|---|---|
| IL-6 | 2.1 | 3.9 | 5.2 | 4.4 | 2.1 |
| (1.2–4.4) | (1.2–9.7) | (2.2–9.9) | (1.2–12.7) | (1.2–14.2) | |
| sgp130 | 19,197 | 17,239 | 22,985 | 17,429 | 18,914 |
| (86–26,942) | (7004–28,049) | (7666–36,063) | (9723–40,714) | (10,596–32,561) | |
| sIL-6R | 4400 | 3952 | 6101 | 4401 * | 4692 |
| (26–6189) | (1932–7938) | (2103–11,681) | (2181–11,843) | (2252–12,936) | |
| IL-31 | 6.7 | 6.4 | 37.8 | 5.3 ** | 6.7 ** |
| (3.6–21.8) | (3.6–19.5) | (5.8–76.8) | (3.6–15.3) | (3.6–9.8) | |
| OSM | 29 | 31 | 94 | 32 ** | 36 ** |
| (7–214) | (8–229) | (11–538) | (8–137) | (10–214) | |
| CNTF | 624 | 571 | 677 | 649 | 571 * |
| (470–1543) | (470–2019) | (494–2507) | (470–1892) | (470–1710) |
| The Peripheral Blood Levels of Immunocompetent Cell Subsets | ||||||||
| Leukocyte subset | Peripheral Blood Level 1 | IL-6 | sIL-6R | sgp130 | IL-31 | OSM | CNTF | CRP |
| T cells, total (CD3+) | 3.31 (1.29–4.17) | −0.042 | −0.508 | −0.697 ** | 0.244 | −0.511 | 0.654 * | 0.156 |
| CD4+ T cells | 2.54 (0.92–3.47) | 0.046 | −0.582 * | −0.609 * | 0.354 | −0.495 | 0.427 | 0.229 |
| CD8+ T cells | 0.60 (0.24–1.08) | −0.135 | −0.205 | −0.557 * | 0.104 | −0.275 | 0.555 * | 0.097 |
| B cells (CD19+) | 0.41 (0.21–1.77) | 0.289 | −0.310 | −0.719 ** | 0.525 | −0.423 | 0.507 | 0.384 |
| NK-cells (CD3− CD56+) | 0.30 (0.07–0.77) | −0.449 | 0.165 | 0.181 | −0.020 | 0.366 | −0.074 | −0.249 |
| Total monocytes | 2.4 (0.90–3.9) | −0.065 | −0.164 | −0.296 | 0.276 | −0.046 | 0.362 | 0.210 |
| CD34+ cells | 40.2 (16.7–148) | −0.21 | −0.32 | −0.54 * | 0.47 | 0.37 | 0.33 | 0.045 |
| The Graft Composition of Immunocompetent Cell Subsets | ||||||||
| Leukocyte subset | Graft Level 1 | IL-6 | sIL-6R | sgp130 | IL-31 | OSM | CNTF | CRP |
| T cells, total (CD3+) | 22.78 (8.41–42.81) | −0.088 | 0.328 | 0.294 | 0.097 | −0.074 | −0.358 | −0.539 * |
| CD4+ T cells | 17.55 (6.02–31.66) | 0.073 | 0.459 | 0.516 | −0.162 | −0.196 | −0.176 | −0.444 |
| CD8+ T cells | 4.64 (1.30–9.74) | −0.068 | 0.336 | 0.204 | 0.087 | −0.007 | 0.268 | −0.592 * |
| B cells (CD19+) | 3.63 (0.00–12.76) | 0.534 * | 0.363 | 0.169 | 0.184 | 0.385 | 0.277 | −0.622 * |
| NK-cells (CD3− CD56+) | 1.79 (0.40–5.50) | −0.121 | 0.253 | 0.433 | 0.315 | 0.415 | 0.121 | −0.407 |
| Total monocytes | 12.91 (1.93–25.23) | −0.248 | 0.071 | 0.100 | 0.248 | 0.324 | −0.054 | −0.256 |
| CD34+ cells | 0.43 (0.085–201) | −0.026 | 0.319 | 0.125 | −0.258 | −0.088 | 0.400 | −0.009 |
| Agonist | Agonist Concentration | IL-6 Supernatant Levels (pg/mL) | Healthy Monocyte Donors | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| PAM3CSK4 (TLR1/2) | 1 and 5 ng/mL | 10.8 (3.1–372) | ||||||||||
| LPS (TLR4) | 5 and 10 ng/mL | 3.1 (3.1–281) | nt | nt | nt | nt | nt | nt | nt | |||
| Flagelin (TLR5) | 10 and 50 ng/mL | 13.9 (3.1–291) | ||||||||||
| R848 (TLR7 > TLR8) | 50 and 100 ng/mL | 3.1 (3.1–180) | ||||||||||
| R837 (TLR7/TLR8) | 0.5 and 1 mg/mL | 188 (3.1–395) | ||||||||||
| Recipients (n = 85) | Characteristics |
|---|---|
| Age, median and range (Years) | 47 (18–70) |
| Diagnosis (number) | |
| AML, de novo | 37 |
| AML secondary to myelodysplastic syndrome | 17 |
| Myelodysplastic syndrome, high-risk | 2 |
| Acute lymphoblastic leukemia | 15 |
| Chronic myeloid leukemia | 3 |
| Myelofibrosis/Myeloproliferative neoplasia, unspecified | 6 |
| Chronic myelomonocytic leukemia | 2 |
| Chronic lymphocytic leukemia | 2 |
| Hodgkin’s lymphoma | 1 |
| Leukemia patients not in remission at transplantation | 1 |
| aGVHD requiring high dose steroid treatment (number) 1 | 38 |
| Conditioning regimes (number) | |
| Busulfan + cyclophosphamide (myeloablative condition) | 66 |
| Fludarabine + busulfan (reduced intensity conditioning) | 16 |
| Others | 3 |
| Stem cell source (number) | |
| Peripheral blood mobilized stem cells | 85 |
| Bone marrow grafts | 0 |
| DONORS (n = 85) | |
| Sibling/other family donors | 78/7 |
| Female/Male | 54/31 |
| Age; median (range) | 49 (18–77) |
| Female donor to male recipient | 19 |
| Number of CMV positive recipients | 60 |
| CMV positive donor to CMV negative recipient | 15 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tvedt, T.H.A.; Melve, G.K.; Tsykunova, G.; Ahmed, A.B.; Brenner, A.K.; Bruserud, Ø. Immunological Heterogeneity of Healthy Peripheral Blood Stem Cell Donors—Effects of Granulocyte Colony-Stimulating Factor on Inflammatory Responses. Int. J. Mol. Sci. 2018, 19, 2886. https://doi.org/10.3390/ijms19102886
Tvedt THA, Melve GK, Tsykunova G, Ahmed AB, Brenner AK, Bruserud Ø. Immunological Heterogeneity of Healthy Peripheral Blood Stem Cell Donors—Effects of Granulocyte Colony-Stimulating Factor on Inflammatory Responses. International Journal of Molecular Sciences. 2018; 19(10):2886. https://doi.org/10.3390/ijms19102886
Chicago/Turabian StyleTvedt, Tor Henrik Anderson, Guro K. Melve, Galina Tsykunova, Aymen Bushra Ahmed, Annette K. Brenner, and Øystein Bruserud. 2018. "Immunological Heterogeneity of Healthy Peripheral Blood Stem Cell Donors—Effects of Granulocyte Colony-Stimulating Factor on Inflammatory Responses" International Journal of Molecular Sciences 19, no. 10: 2886. https://doi.org/10.3390/ijms19102886
APA StyleTvedt, T. H. A., Melve, G. K., Tsykunova, G., Ahmed, A. B., Brenner, A. K., & Bruserud, Ø. (2018). Immunological Heterogeneity of Healthy Peripheral Blood Stem Cell Donors—Effects of Granulocyte Colony-Stimulating Factor on Inflammatory Responses. International Journal of Molecular Sciences, 19(10), 2886. https://doi.org/10.3390/ijms19102886