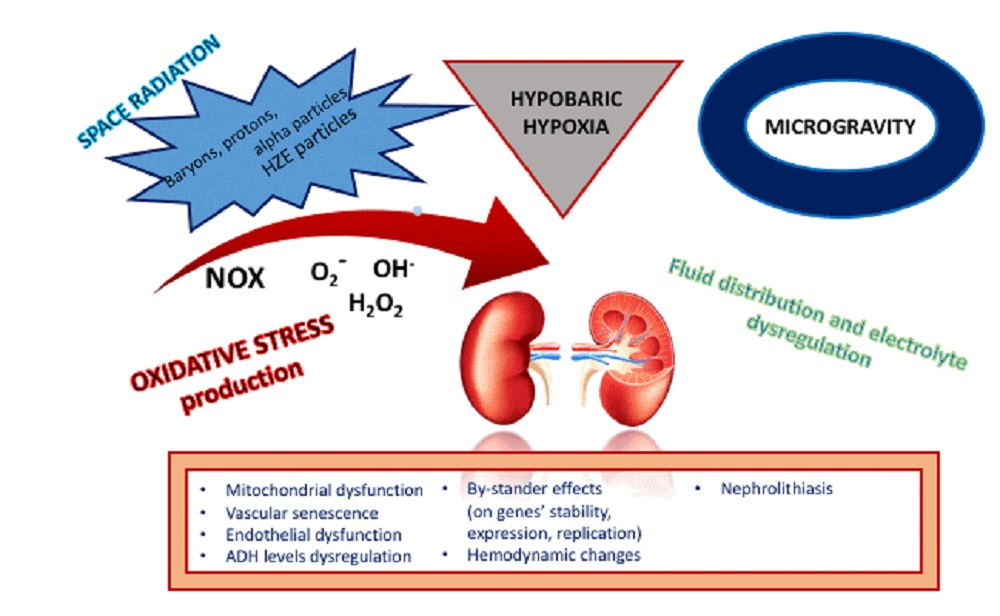
Oxidative Stress and the Kidney in the Space Environment



 and
and 
Abstract
:
1. Introduction
1.1. The Induction of Oxidative Stress in Space: How The Story Starts
1.2. Fluid Balance in Space and Hypobaric Hypoxia
1.3. The Kidney, Mitochondria and Oxidative Stress
1.4. Mitochondrial Changes after Radiation Exposure
1.5. NADPH Oxidases, Endothelial Function, and Oxidative Stress in Space: What We Know about the Renal Response
1.6. NADPH Oxidases in the Pathophysiology of Hypertension: How Does the Story Go in Space?
1.7. Space and Other Kidney-Related Manifestations
2. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pietrofesa, R.A.; Turowski, J.B.; Arguiri, E.; Milovanova, T.N.; Solomides, C.C.; Thom, S.R. Oxidative Lung Damage Resulting from Repeated Exposure to Radiation and Hyperoxia Associated with Space Exploration. J. Pulm. Respir. Med. 2013, 30, 61–75. [Google Scholar]
- Law, J.; Mathers, C.H.; Fondy, S.R.E.; Vanderploeg, J.M.; Kerstman, E.L. NASA’s human system risk management approach and its applicability to commercial spaceflight. Aviat. Space Environ. Med. 2013, 84, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Hellweg, C.E.; Baumstark-Khan, C. Getting ready for the manned mission to Mars: The astronauts’ risk from space radiation. Naturwissenschaften 2007, 5, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Karmali, F.; Shelhamer, M. The dynamics of parabolic flight: Flight characteristics and passenger percepts. Acta Astronaut 2008, 63, 594–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.; Askew, C.D.; Brümmer, V.; Kleinert, J.; Guardiera, S.; Abel, T. The effect of parabolic flight on perceived physical, motivational and psychological state in men and women: Correlation with neuroendocrine stress parameters and electrocortical activity. Stress 2009, 23, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Goswami, N. Falls and Fall-Prevention in Older Persons: Geriatrics Meets Spaceflight! Front Physiol. 2017, 8, 603. [Google Scholar] [CrossRef] [PubMed]
- Townsend, L.W.; Shinn, J.L.; Wilson, J.W. Interplanetary crew exposure estimates for the August 1972 and October 1989 solar particle events. Radiat. Res. 1991, 126, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A. Biophysics of NASA radiation quality factors. Radiat Prot Dosimetry. 2015, 166, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Dietze, G.; Bartlett, D.T.; Cool, D.A.; Cucinotta, F.A.; Jia, X.; McAulay, I.R.; Pelliccioni, M.; Petrov, V.; Reitz, G.; Sato, T. ICRP publication 123: Assessment of radiation exposure of astronauts in space. Ann. ICRP 2013, 42, 1–339. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.S.; Bloch, P.; Ware, J.H.; Zhou, Z.; Donahue, J.J.; Guan, J. Detection of oxidative stress induced by low- and high-linear energy transfer radiation in cultured human epithelial cells. Radiat. Res. 2005, 163, 364–368. [Google Scholar] [CrossRef]
- Li, M.; Gonon, G.; Buonanno, M.; Autsavapromporn, N.; de Toledo, S.M.; Pain, D. Health risks of space exploration: Targeted and nontargeted oxidative injury by high-charge and high-energy particles. Antioxid. Redox. Signal. 2014, 20, 1501–1523. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Arfat, Y.; Wang, H.; Goswami, N. Muscle Atrophy Induced by Mechanical Unloading: Mechanisms and Potential Countermeasures. Front Physiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Stein, T.P. Space flight and oxidative stress. Nutrition 2002, 18, 867–871. [Google Scholar] [CrossRef]
- Mao, X.W.; Pecaut, M.J.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Bouxsein, M. Spaceflight environment induces mitochondrial oxidative damage in ocular tissue. Radiat. Res. 2013, 180, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Tseng, B.P.; Giedzinski, E.; Izadi, A.; Suarez, T.; Lan, M.L.; Tran, K.K. Functional consequences of radiation-induced oxidative stress in cultured neural stem cells and the brain exposed to charged particle irradiation. Antioxid. Redox. Signal. 2014, 20, 1410–1422. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Nishiyama, N.C.; Pecaut, M.J.; Campbell-Beachler, M.; Gifford, P.; Haynes, K.E. Simulated microgravity and low-dose/low-dose-rate radiation induces oxidative damage in the mouse brain. Radiat. Res. 2016, 185, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Pecaut, M.J.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Bouxsein, M.L. Biological and metabolic response in STS-135 space-flown mouse skin. Free Radic. Res. 2014, 28, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Datta, K.; Suman, S.; Kallakury, B.V.S.; Fornace, A.J. Exposure to heavy ion radiation induces persistent oxidative stress in mouse intestine. PLoS ONE 2012, 7, e42224. [Google Scholar] [CrossRef] [PubMed]
- Versari, S.; Longinotti, G.; Barenghi, L.; Maier, J.A.M.; Bradamante, S. The challenging environment on board the International Space Station affects endothelial cell function by triggering oxidative stress through thioredoxin interacting protein overexpression: The ESA-SPHINX experiment. FASEB J. 2013, 27, 4466–4475. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Martin, S.G. The thioredoxin system: A key target in tumour and endothelial cells. Br. J. Radiol. 2008, 81, S57–S68. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-B.; Han, Y.; Zhang, S.; Cui, N.-H.; Liu, Z.-J.; Huang, Z.-L. Associations of polymorphisms in TXNIP and gene-environment interactions with the risk of coronary artery disease in a Chinese Han population. J. Cell Mol. Med. 2016, 20, 2362–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Chng, W.-J. Roles of thioredoxin binding protein (TXNIP) in oxidative stress, apoptosis and cancer. Mitochondrion 2013, 13, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Okumura, H.; Guo, R.; Naruse, K. Effect of oxidative stress on cardiovascular system in response to gravity. Int. J. Mol. Sci. 2017, 18, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, K.A.; Röcker, L.; Gauer, O.H.; Krause, R.; Leach, C.; Wicke, H.J. Venous pressure in man during weightlessness. Science 1984, 225, 218–219. [Google Scholar] [CrossRef] [PubMed]
- Gauer, O.H.; Henry, J.P.; Sieker, H.O. Cardiac receptors and fluid volume control. Prog Cardiovasc. Dis. 1961, 1, 1–26. [Google Scholar] [CrossRef]
- Drummer, C.; Norsk, P.; Heer, M. Water and sodium balance in space. Am. J. Kidney Dis. 2001, 38, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Norsk, P. Renal adjustments to microgravity. Pflugers Arch. 2000, 441, R62–R65. [Google Scholar] [CrossRef] [PubMed]
- Leach, C.S.; Alfrey, C.P.; Suki, W.N.; Leonard, J.I.; Rambaut, P.C.; Inners, L.D. Regulation of body fluid compartments during short-term spaceflight. J. Appl. Physiol. 1996, 81, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Kramer, H.J.; Heer, M.; Cirillo, M.; De Santo, N.G. Renal hemodynamics in space. Am. J. Kidney Dis. 2001, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Drummer, C.; Valenti, G.; Cirillo, M.; Perna, A.; Bellini, L.; Nenov, V. Vasopressin, Hypercalciuria and Aquaporin—The Key Elements for Impaired Renal Water Handling in Astronauts? Nephron 2002, 92, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Gaspare De Santo, N.; Cirillo, M.; Valenti, G.; Perna, A.; Anastasio, P.; Drummer, C. Renal function in space: The link between osteoporosis, hypercalciuria, and aquaporins. J. Ren. Nutr. 2005, 15, 1831–1888. [Google Scholar] [CrossRef]
- Tokumaru, O. Prevention of decompression sickness during extravehicular activity in space: A review. Boei Ika Daigakko Zasshi 1997, 22, 223–232. [Google Scholar] [PubMed]
- Webb, J.T.; Pilmanis, A.A. Fifty years of decompression sickness research at Brooks AFB, TX: 1960–2010. Aviat. Space Environ. Med. 2011, 82, A1–A25. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb-Rumyantzev, A.S.; Alper, S.L. Short-term responses of the kidney to high altitude in mountain climbers. Nephrol. Dial. Transplant. 2014, 29, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Farías, J.G.; Herrera, E.A.; Carrasco-Pozo, C.; Sotomayor-Zárate, R.; Cruz, G.; Morales, P. Pharmacological models and approaches for pathophysiological conditions associated with hypoxia and oxidative stress. Pharmacol. Ther. 2016, 158, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Mori, T.; Huang, T.; Lombard, J.H. Effect of high-salt diet on NO release and superoxide production in rat aorta. Am. J. Physiol. Circ. Physiol. 2004, 286, H575–H583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boegehold, M.A. Effect of dietary salt on arteriolar nitric oxide in striated muscle of normotensive rats. Am. J. Physiol. Circ. Physiol. 1993, 264, H1810–H1816. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, K.M.; Clifton, P.M.; Keogh, J.B. Endothelial function is impaired after a high-salt meal in healthy subjects. Am. J. Clin. Nutr. 2011, 93, 500–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H. Oxidative Stress: Introductory Remarks. Oxidat. Stress 1985, 5, 1–8. [Google Scholar]
- Lushchak, V.I. Chemico-Biological Interactions Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 18, 62–73. [Google Scholar]
- Rizzo, A.M.; Berselli, P.; Zava, S.; Montorfano, G.; Negroni, M.; Corsetto, P. Endogenous Antioxidants and Radical Scavengers; Springer: Boston, MA, USA, 2010; pp. 52–67. [Google Scholar]
- Epstein, F.H. Oxygen and renal metabolism. Kidney Int. 1997, 51, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Che, R.; Yuan, Y.; Huang, S.; Zhang, A. Mitochondrial dysfunction in the pathophysiology of renal diseases. Am. J. Physiol. Ren. Physiol. 2014, 306, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Pavlakou, P.; Liakopoulos, V.; Eleftheriadis, T.; Mitsis, M.; Dounousi, E. Oxidative Stress and Acute Kidney Injury in Critical Illness: Pathophysiologic Mechanisms—Biomarkers—Interventions, and Future Perspectives. Oxid. Med. Cell. Longev. 2017, 18, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Szumiel, I. Ionizing radiation-induced oxidative stress, epigenetic changes and genomic instability: The pivotal role of mitochondria. Int. J. Radiat. Biol. 2015, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Goto, S.; Kawakatsu, M.; Urata, Y.; Li, T.S. Mitochondrial dysfunction, a probable cause of persistent oxidative stress after exposure to ionizing radiation. Free. Radic. Res. 2012, 46, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indo, H.P.; Majima, H.J.; Terada, M.; Suenaga, S.; Tomita, K.; Yamada, S. Changes in mitochondrial homeostasis and redox status in astronauts following long stays in space. Sci. Rep. 2016, 6, 39015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, J.L.; Mohamadzadeh, M. Macrophages and chemokines as mediators of angiogenesis. Front. Physiol. 2013, 4, 159–162. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, L.A. Angiogenesis and scar formation in healing wounds. Curr. Opin. Rheumatol. 2013, 25, 87–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-W.; West, X.Z.; Byzova, T.V. Inflammation and oxidative stress in angiogenesis and vascular disease. J. Mol. Med. 2013, 9, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar] [CrossRef] [PubMed]
- El-Refaei, M.F.; Abduljawad, S.H.; Alghamdi, A.H. Alternative Medicine in Diabetes—Role of Angiogenesis, Oxidative Stress, and Chronic Inflammation. Rev. Diabet. Stud. 2014, 11, 231–244. [Google Scholar] [CrossRef] [PubMed]
- West, X.Z.; Malinin, N.L.; Merkulova, A.A.; Tischenko, M.; Kerr, B.A.; Borden, E.C. Oxidative stress induces angiogenesis by activating TLR2 with novel endogenous ligands. Nature 2010, 467, 972–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera, S.; Martínez, R.; Gormaz, J.G.; Gajardo, A.; Galleguillos, F.; Rodrigo, R. Novel relationships between oxidative stress and angiogenesis-related factors in sepsis: New biomarkers and therapies. Ann. Med. 2015, 47, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Takac, I.; Schröder, K.; Brandes, R.P. The Nox Family of NADPH Oxidases: Friend or Foe of the Vascular System? Curr. Hypertens. Rep. 2012, 14, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Ebrahem, Q.; Renganathan, K.; Sears, J.; Vasanji, A.; Gu, X.; Lu, L. Carboxyethylpyrrole oxidative protein modifications stimulate neovascularization: Implications for age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 13480–13484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duni, A.; Liakopoulos, V.; Rapsomanikis, K.-P.; Dounousi, E. Chronic Kidney Disease and Disproportionally Increased Cardiovascular Damage: Does Oxidative Stress Explain the Burden? Oxid. Med. Cell. Longev. 2017, 2017, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lassègue, B.; San Martín, A.; Griendling, K.K. Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ. Res. 2012, 110, 1364–1390. [Google Scholar] [CrossRef] [PubMed]
- Yentrapalli, R.; Azimzadeh, O.; Barjaktarovic, Z.; Sarioglu, H.; Wojcik, A.; Harms-Ringdahl, M. Quantitative proteomic analysis reveals induction of premature senescence in human umbilical vein endothelial cells exposed to chronic low-dose rate gamma radiation. Proteomics 2013, 110, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Podlutsky, A.; Sosnowska, D.; Tucsek, Z.; Toth, P.; Deak, F. Ionizing radiation promotes the acquisition of a senescence-associated secretory phenotype and impairs angiogenic capacity in cerebromicrovascular endothelial cells: Role of increased DNA damage and decreased DNA repair capacity in microvascular radiosensitivity. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1443–1457. [Google Scholar] [PubMed]
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The senescence-associated secretory phenotype: The dark side of tumor suppression. Annu. Rev. Pathol. 2010, 5, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, S.V.; Ermakov, A.V.; Alekseeva, A.Y.; Smirnova, T.D.; Glebova, K.V.; Efremova, L.V. Role of extracellular DNA oxidative modification in radiation induced bystander effects in human endotheliocytes. Mutat. Res. Mol. Mech. Mutagen. 2012, 729, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Azimzadeh, O.; Sievert, W.; Sarioglu, H.; Merl-Pham, J.; Yentrapalli, R.; Bakshi, M.V. Integrative Proteomics and Targeted Transcriptomics Analyses in Cardiac Endothelial Cells Unravel Mechanisms of Long-Term Radiation-Induced Vascular Dysfunction. J. Proteome Res. 2015, 14, 1203–1219. [Google Scholar] [CrossRef] [PubMed]
- Rabelink, T.J.; Zonneveld, A. Coupling eNOS Uncoupling to the Innate Immune Response. Arterioscler Thromb. Vasc. Biol. 2006, 26, 2585–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedeek, M.; Nasrallah, R.; Touyz, R.M.; Hébert, R.L. NADPH oxidases, reactive oxygen species, and the kidney: Friend and foe. J. Am. Soc. Nephrol. 2013, 24, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.G.; Schnermann, J. Integrated control of Na transport along the nephron. Clin. J. Am. Soc. Nephrol. 2015, 10, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Drummer, C.; Heer, M.; Dressendrfer, R.A.; Strasburger, C.J.; Gerzer, R. Reduced natriuresis during weightlessness. Clin. Investig. 1993, 71, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Valentin, J.P.; Humphreys, M.H. Urodilatin: A paracrine renal natriuretic peptide. Semin. Nephrol. 1993, 13, 61–70. [Google Scholar] [PubMed]
- Drummer, C.; Heer, M.; Herten, M.; Gerzer, R. Regulation of volume homeostasis in reduced gravity: Possible involvement of atrial natriuretic peptide, renal natriuretic peptide and cyclic GMP, Projektbericht, DLR-Interner Bericht. Anthrorack Spacelab Mission 1997, 22, 55–61. [Google Scholar]
- Drummer, C.; Hesse, C.; Baisch, F.; Norsk, P.; Elmann-Larsen, B.; Gerzer, R. Water and sodium balances and their relation to body mass changes in microgravity. Eur. J. Clin. Investig. 2000, 30, 1066–1075. [Google Scholar] [CrossRef]
- Gill, P.S.; Wilcox, C.S. NADPH Oxidases in the Kidney. Antioxid. Redox. Signal. 2006, 8, 1597–1607. [Google Scholar] [CrossRef] [PubMed]
- Han, H.J.; Lee, Y.J.; Park, S.H.; Lee, J.H.; Taub, M. High glucose-induced oxidative stress inhibits Na+/glucose cotransporter activity in renal proximal tubule cells. Am. J. Physiol. Physiol. 2005, 288, F988–F996. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.P.; Jandeleit-Dahm, K.A.M. The role of NADPH oxidase in vascular disease—Hypertension, atherosclerosis stroke. Curr. Pharm. Des. 2015, 21, 5933–5944. [Google Scholar] [CrossRef] [PubMed]
- Dikalova, A.; Clempus, R.; Lassègue, B.; Cheng, G.; McCoy, J.; Dikalov, S. Nox1 Overexpression Potentiates Angiotensin II-Induced Hypertension and Vascular Smooth Muscle Hypertrophy in Transgenic Mice. Circulation 2005, 112, 2668–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.D.; Xu, S.; Johns, D.G.; Du, Y.; Quinn, M.T.; Cayatte, A.J. Role of NADPH oxidase in the vascular hypertrophic and oxidative stress response to angiotensin II in mice. Circ. Res. 2001, 88, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Salazar, G. NADPH oxidases and mitochondria in vascular senescence. Int. J. Mol. Sci. 2018, 19, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Sirker, A.; Zhang, M.; Shah, A.M. NADPH oxidases in cardiovascular disease: Insights from in vivo models and clinical studies. Basic Res. Cardiol. 2011, 106, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yi, F.-X.; Spurrier, J.L.; Bobrowitz, C.A.; Zou, A.-P. Production of superoxide through NADH oxidase in thick ascending limb of Henle’s loop in rat kidney. Am. J. Physiol. Ren. Physiol. 2002, 282, F1111–F1119. [Google Scholar] [CrossRef] [PubMed]
- Laursen, J.B.; Rajagopalan, S.; Galis, Z.; Tarpey, M.; Freeman, B.A.; Harrison, D.G. Role of superoxide in angiotensin II-induced but not catecholamine-induced hypertension. Circulation 1997, 95, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.Z.; Majid, D.S.A. Assessment of Renal Functional Phenotype in Mice Lacking gp91PHOX Subunit of NADPH. Oxidase. Hypertension 2004, 43, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Kitiyakara, C.; Chabrashvili, T.; Chen, Y.; Blau, J.; Karber, A.; Aslam, S. Salt intake, oxidative stress, and renal expression of NADPH oxidase and superoxide dismutase. J. Am. Soc. Nephrol. 2003, 14, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Rowe, W.J. Potential renovascular hypertension, space missions, and the role of magnesium. Int. J. Nephrol. Renovasc. Dis. 2009, 2, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Guan, Y.; Zhang, L.; Tian, Y.; Zhang, Y.; Wang, S. Depressive effects of chronic intermittent hypobaric hypoxia on renal vascular hypertension through enhancing baroreflex. Chin. J. Physiol. 2016, 59, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.P.C.; Segreto, H.R.C.; Boim, M.A.; Razvickas, C.V.; Schor, N. Effects of Ionizing Radiation on Progressive Experimental Renal Disease: A Hemodynamic Approach. Nephron 2001, 87, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Fritsch-Yelle, J.M.; Charles, J.B.; Jones, M.M.; Wood, M.L. Microgravity decreases heart rate and arterial pressure in humans. J. Appl. Physiol. 1996, 80, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, M.; Kamo, T.; Kamegai, M.; Baevsky, R.M.; Funtova, I.I.; Chernikova, A. Periodic structures and diurnal variation in blood pressure and heart rate in relation to microgravity on space station MIR. Biomed. Pharmacother. 2004, 58, S31–S34. [Google Scholar] [CrossRef]
- Pietrzyk, R.A.; Jones, J.A.; Sams, C.F.; Whitson, P.A. Renal stone formation among astronauts. Aviat. Space Environ. Med. 2007, 78, A9–A13. [Google Scholar] [PubMed]
- Kassemi, M.; Thompson, D. Prediction of renal crystalline size distributions in space using a PBE analytic model. 1. Effect of microgravity-induced biochemical alterations. Am. J. Physiol. Ren. Physiol. 2016, 311, F520–F530. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; MacLeod, R.J. Extracellular Calcium Sensing and Extracellular Calcium Signaling. Physiol. Rev. 2001, 81, 239–297. [Google Scholar] [CrossRef] [PubMed]
- Procino, G.; Mastrofrancesco, L.; Tamma, G.; Lasorsa, D.R.; Ranieri, M.; Stringini, G. Calcium-sensing receptor and aquaporin 2 interplay in hypercalciuria-associated renal concentrating defect in humans. An in vivo and in vitro study. PLoS ONE 2012, 7, e33145. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, V.; Leivaditis, K.; Eleftheriadis, T.; Dombros, N. The kidney in space. Int. Urol. Nephrol. 2012, 44, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Heer, M.; Shackelford, L.C.; Sibonga, J.D.; Spatz, J.; Pietrzyk, R.A. Bone metabolism and renal stone risk during International Space Station missions. Bone 2015, 81, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Zwart, S.R.; Heer, M.; Hudson, E.K.; Shackelford, L.; Morgan, J.L.L. Men and women in space: Bone loss and kidney stone risk after long-duration spaceflight. J. Bone Miner. Res. 2014, 29, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Çiftçioglu, N.; Björklund, M.; Kuorikoski, K.; Bergström, K.; Kajander, E.O. Nanobacteria: An infectious cause for kidney stone formation. Kidney Int. 1999, 56, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Çiftçioǧlu, N.; Haddad, R.S.; Golden, D.C.; Morrison, D.R.; McKay, D.S. A potential cause for kidney stone formation during space flights: Enhanced growth of nanobacteria in microgravity. Kidney Int. 2005, 67, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Ansari, H.; Akhavan Sepahi, A.; Akhavan Sepahi, M. Different Approaches to Detect Nanobacteria in Patients with Kidney Stones: An Infectious Cause or a Subset of Life? Urol. J. 2017, 14, 5001–5007. [Google Scholar] [PubMed]
- Kassemi, M.; Thompson, D. Prediction of renal crystalline size distributions in space using a PBE analytic model. 2. Effect of dietary countermeasures. Am. J. Physiol. Ren. Physiol. 2016, 311, F531–F538. [Google Scholar] [CrossRef] [PubMed]
- Zerwekh, J.E.; Odvina, C.V.; Wuermser, L.A.; Pak, C.Y.C. Reduction of Renal Stone Risk by Potassium-Magnesium Citrate During 5 Weeks of Bed Rest. J. Urol. 2007, 177, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Gáspár, S.; Niculiţe, C.; Cucu, D.; Marcu, I. Effect of calcium oxalate on renal cells as revealed by real-time measurement of extracellular oxidative burst. Biosens. Bioelectron. 2010, 25, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Boonla, C.; Wunsuwan, R.; Tungsanga, K.; Tosukhowong, P. Urinary 8-hydroxydeoxyguanosine is elevated in patients with nephrolithiasis. Urol Res. 2007, 35, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Abdelhafiz, A.H.; Ahmed, S.; Nahas, M. El. Microalbuminuria: Marker or Maker of Cardiovascular Disease. Nephron. Exp. Nephrol. 2011, 119, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Cerasola, G.; Cottone, S.; Mulè, G. The progressive pathway of microalbuminuria: From early marker of renal damage to strong cardiovascular risk predictor. J. Hypertens. 2010, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Díaz, A.G.; Pazarín-Villaseñor, L.; Yanowsky-Escatell, F.G.; Andrade-Sierra, J. Oxidative Stress in Diabetic Nephropathy with Early Chronic Kidney Disease. J. Diabetes Res. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, E.G.; Alkhalaf, A.; Landman, G.W.D.; Hateren, K.J.J.; Groenier, K.H.; Struck, J. Serum Peroxiredoxin 4: A Marker of Oxidative Stress Associated with Mortality in Type 2 Diabetes (ZODIAC-28). PLoS ONE 2014, 9, e89719. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, J.; Valentini, J.; Grotto, D.; Paniz, C.; Roehrs, M.; Brucker, N. Association Among Microalbuminuria and Oxidative Stress Biomarkers in Patients with Type 2 Diabetes. J. Investig. Med. 2011, 59, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Giner, V.; Tormos, C.; Chaves, F.J.; Saez, G.; Redon, J. Microalbuminuria and oxidative stress in essential hypertension. J. Intern. Med. 2004, 255, 588–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirillo, M.; De Santo, N.G.; Heer, M.; Norsk, P.; Elmann-Larsen, B.; Bellini, L. Low urinary albumin excretion in astronauts during space missions. Nephron 2003, 93, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Pastushkova, L.K.; Kireev, K.S.; Kononikhin, A.S.; Tiys, E.S.; Popov, I.A.; Starodubtseva, N.L. Detection of Renal Tissue and Urinary Tract Proteins in the Human Urine after Space Flight. PLoS ONE 2013, 8, e71652. [Google Scholar] [CrossRef] [PubMed]
- Larina, I.M.; Pastushkova, L.K.; Tiys, E.S.; Kireev, K.S.; Kononikhin, A.S.; Starodubtseva, N.L. Permanent proteins in the urine of healthy humans during the Mars-500 experiment. J. Bioinform. Comput. Biol. 2015, 13, 35–39. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Year | Type of Study | Study Facilities/Environment | Duration | Subject Characteristics | Methods | Main Findings | |
|---|---|---|---|---|---|---|---|
| 10Wan, S.; et al. | 2005 | In Vitro | Laboratory facility | Human breast cells | Radiation (γ-, X-rays, protons, HZE particles, 56Fe ions) | OS correlated with radiation in a dose-depended pattern. HZE particle radiation was less effective in producing OS compared to γ- and Χ-rays | |
| 19Versari, et al. | 2013 | In Vitro | Spaceflight | 10 days | Human umbilical vein endothelial cells (HUVECs) | Post-flight microarray gene analysis | 1023 significantly modulated genes. Thioredoxin was the most highly upregulated. Heat-shock protein 90 and 70 were the most downregulated. Significant increase of IL-1a and IL-1β. |
| 47Indo, et al. | 2016 | Cohort study | Spaceflight, post-flight | 6 months | Hair samples by astronauts | Quantitative PCR for mtDNA/nDNA and mtRNA/nRNA. Antioxidant related gene expression (MnSOD, CuZnSOD, Nrf2, Keap1, GPx4 and Catalase) | Significant reduction of mtDNA/nDNA ratio during flight. Significant reductions in the mtRNA/nRNA ratios in inflight and post-flight samples. All antioxidant related genes’ expression was significantly decreased. Catalase gene expression remained unchanged. |
| 69Drummer C, et al. | 1997 | Observational study | Space mission | 1 week | Male astronaut | Saliva and urine sampling on prespecified time points. | Reduction of natriuresis and diuresis throughout the flight. Urodilatin excretion correlated with sodium excretion. |
| 71Drummer C, et al. | 1997 | Observational study | Space mission | 5 days | 4 male astronauts | Urine and blood samples on prespecified time points (plus before 1.6L isotonic saline infusion). | ANP and cGMP levels did not increase after the infusion of saline (simulated plasma expansion). Renal excretion of urodilatin increased for several hours post saline infusion. |
| 72Drummer, et al. | 2000 | Observational study | Space missions (EuroMIR94 and MIR97) | 30 and 17 days | 3 astronauts | Diet monitoring. Water and sodium balance. Body mass measurement. Blood samples. | Body weight loss is attributed to reduced caloric intake during spaceflight. Microgravity provokes sodium retention through hormonal dysregulations. |
| 87Fritsch-Yelle, J.; et al. | 1996 | Observational study | Space missions | 5 to 10 days | 12 male astronauts | Blood pressure and heart rate monitoring during prespecified intervals. | Significant reduction of heart rate, diastolic blood pressure and premature ventricular contractions during spaceflight. |
| 88Shiraishi, et al. | 2004 | Observational study | Space mission (MIR) | 6 months | 4 male astronauts | Blood pressure and heart rate measurement. | Systolic blood pressure during sleep on spaceflight were increased compared to earth. The initially prolonged periodicity of blood pressure and heart rate was shortened to pre-flight values after a few months on space mission. |
ROS generation
|
| Mitochondrial dysfunction |
| Endothelial dysfunction |
| Vascular senescence |
|
Non-targeted effects (on genes’ stability, expression, replication) |
Hemodynamic changes
|
| Nephrolithiasis |
| ADH levels alteration |
| Hypercalciuria/hypocitraturia |
| Sodium handling |
| Authors | Year | Type of Study | Study Facilities/Environment | Duration | Subject Characteristics | Methods | Main Findings |
|---|---|---|---|---|---|---|---|
| 95Smith, et al. | 2015 | Retrospective analysis | Space missions | Variable (>100 days) | 23 astronauts (four women) Seven astronauts received aledronate (six men, one woman) | Blood and urine samples (pre and during spaceflight). Resistive exercise equipment. | Risk of nephrolithiasis increased during spaceflight (irrespectively to exercise intensity) in all groups. Significant increase of sclerostin during spaceflight in all groups. Urine supersaturation risk was higher compared to the general population. |
| 96Smith, et al. | 2014 | Retrospec tive analysis | Space missions | Variable (49–215 days) | 42 astronauts (33 men and nine women) | Blood and urine samples pre and post flight. Bone densitometry. Resistive exercise equipment. | No sex difference in the response of bone mineral density. Equal risk for urine supersaturation between sexes. Equal risk for stone formation between sexes. |
| 98Ciftcioglu, et al. | 2005 | In vitro study | High aspect rotation vessels (HARV)—microgravity simulation. Stationary and shaker cultures. | Non applicable | Nanobacteria cultures | Spectrophotometer analysis, SEM, TEM, EDX. | 4.6 times faster multiplication of nanobacteria in HARVs. Existence of apatite crystals on all nanobacteria cultures. Thinner layer of apatite on HARV cultures. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlakou, P.; Dounousi, E.; Roumeliotis, S.; Eleftheriadis, T.; Liakopoulos, V. Oxidative Stress and the Kidney in the Space Environment. Int. J. Mol. Sci. 2018, 19, 3176. https://doi.org/10.3390/ijms19103176
Pavlakou P, Dounousi E, Roumeliotis S, Eleftheriadis T, Liakopoulos V. Oxidative Stress and the Kidney in the Space Environment. International Journal of Molecular Sciences. 2018; 19(10):3176. https://doi.org/10.3390/ijms19103176
Chicago/Turabian StylePavlakou, Paraskevi, Evangelia Dounousi, Stefanos Roumeliotis, Theodoros Eleftheriadis, and Vassilios Liakopoulos. 2018. "Oxidative Stress and the Kidney in the Space Environment" International Journal of Molecular Sciences 19, no. 10: 3176. https://doi.org/10.3390/ijms19103176
APA StylePavlakou, P., Dounousi, E., Roumeliotis, S., Eleftheriadis, T., & Liakopoulos, V. (2018). Oxidative Stress and the Kidney in the Space Environment. International Journal of Molecular Sciences, 19(10), 3176. https://doi.org/10.3390/ijms19103176