Effects of OsCDPK1 on the Structure and Physicochemical Properties of Starch in Developing Rice Seeds


Abstract
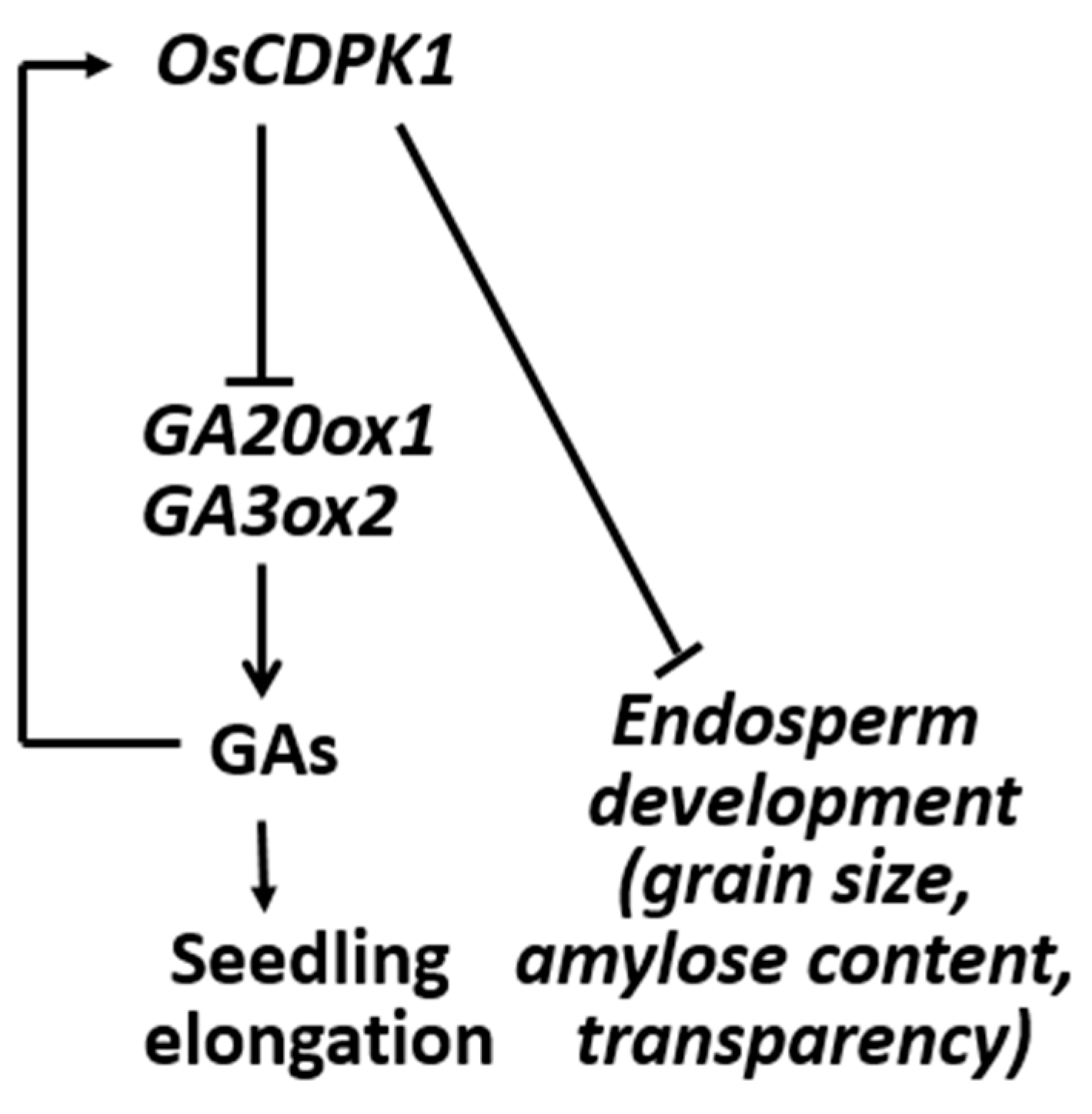
:1. Introduction
2. Results
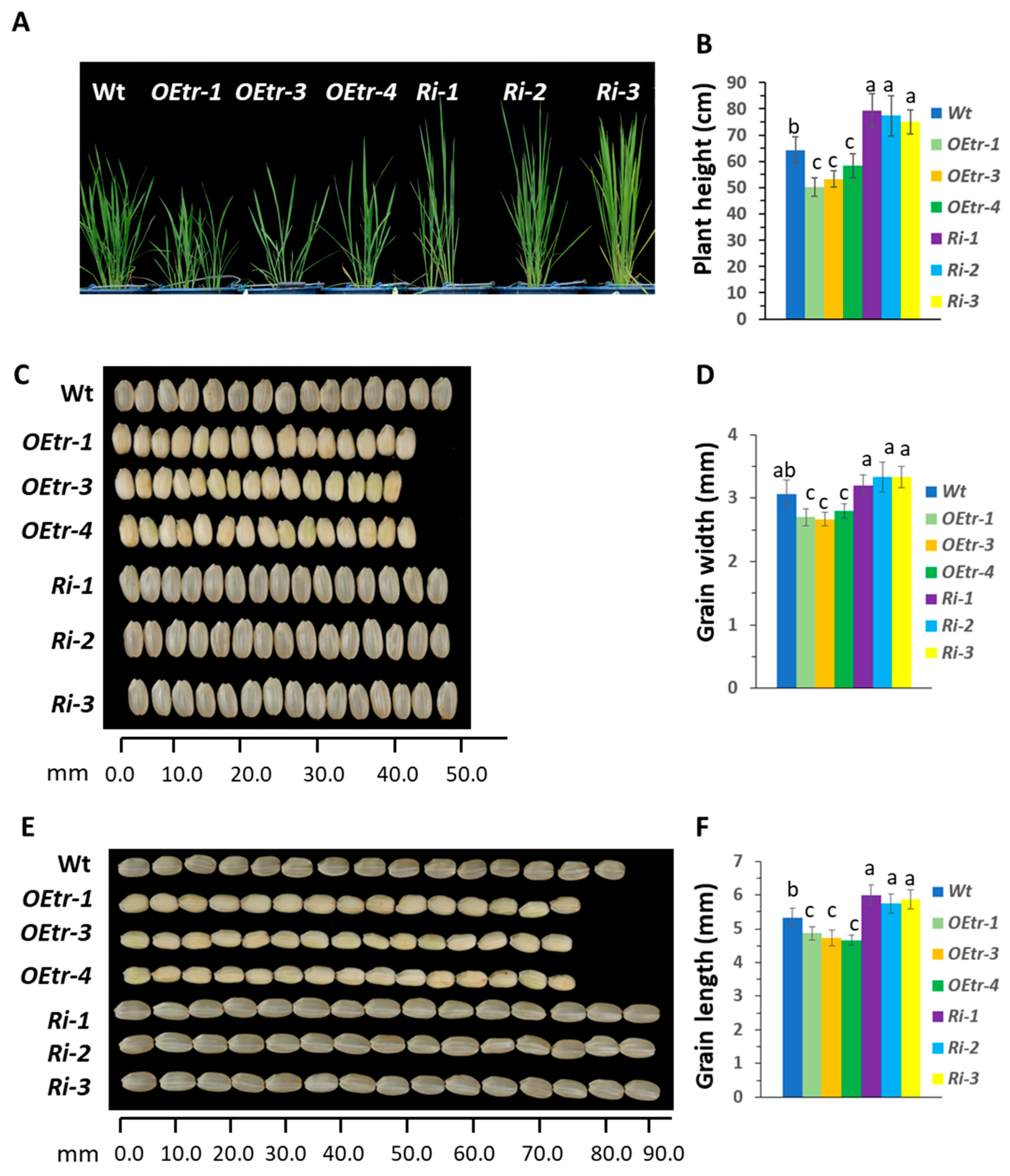
2.1. Phenotypic Changes in Transgenic Rice Plants with Overexpressing or Silenced OsCDPK1
2.2. Ectopic Overexpression and Silencing of OsCDPK1 in Transgenic Rice Plants Yielded Opaque and Transparent Endosperms, Respectively
2.3. Effect of OsCDPK1 on Starch Granule Morphology in Rice Endosperms
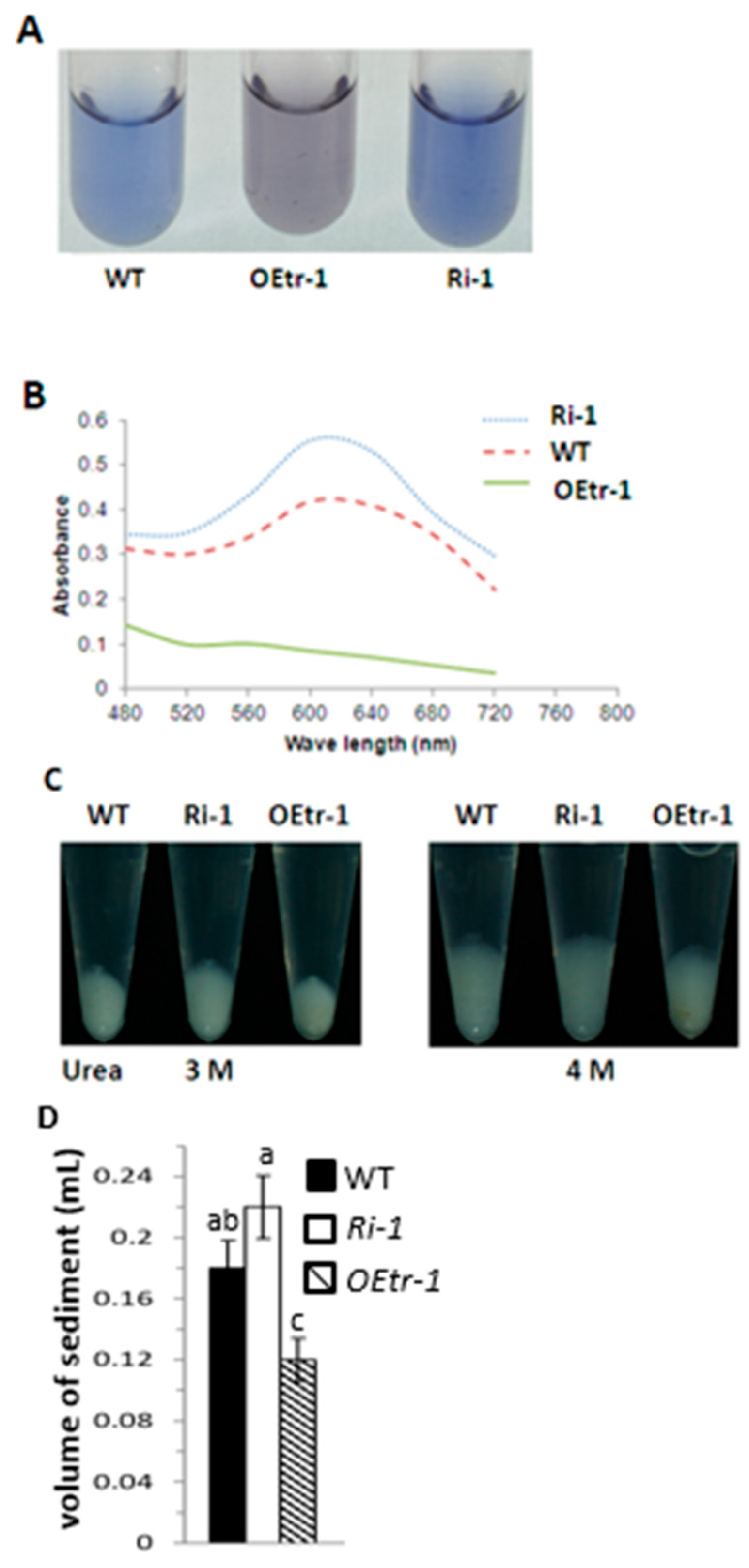
2.4. Effects of OsCDPK1 on Starch Properties and Gelatinization in the Endosperm
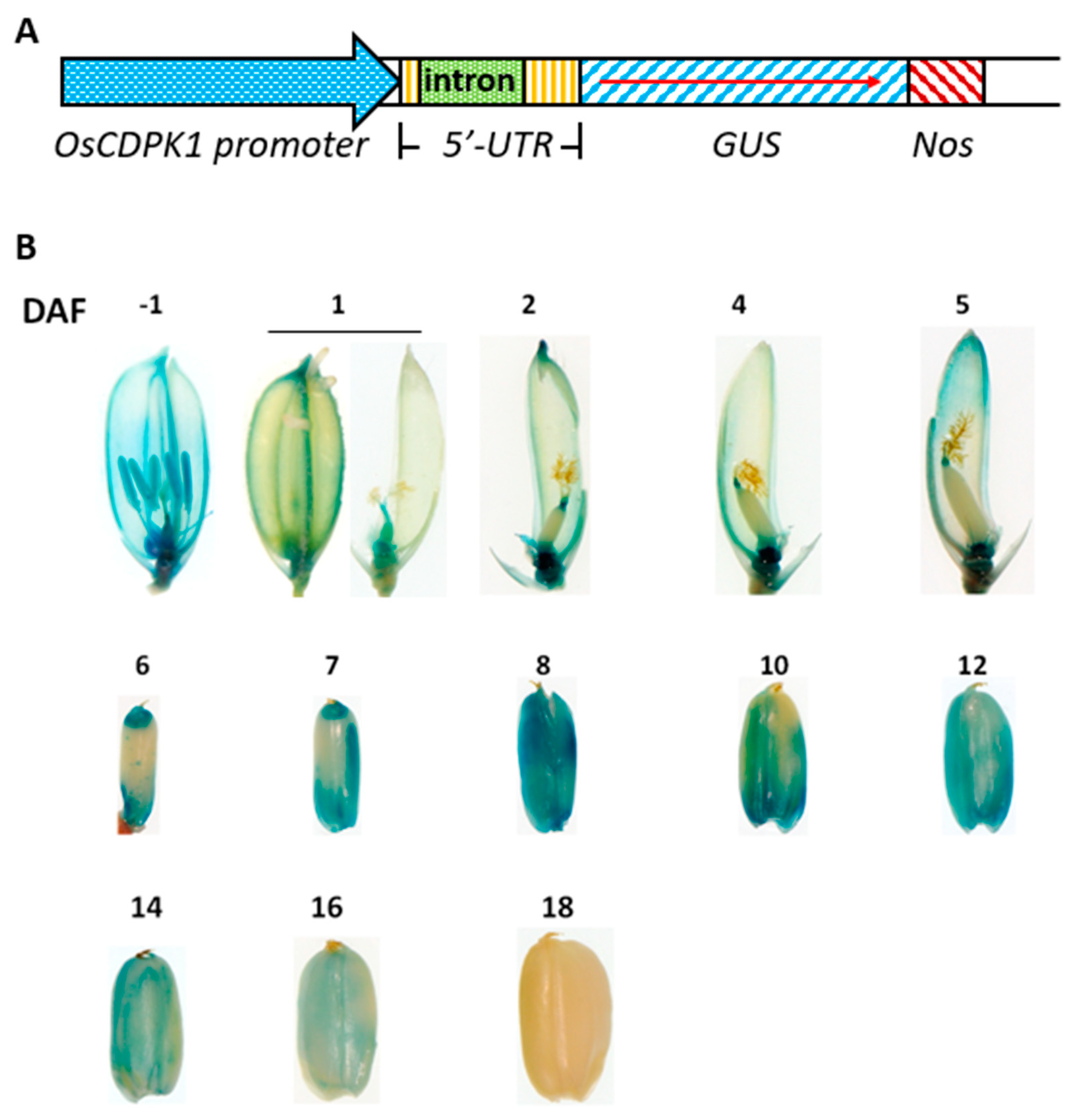
2.5. OsCDPK1 Expression Profiles in Developing Rice Seeds
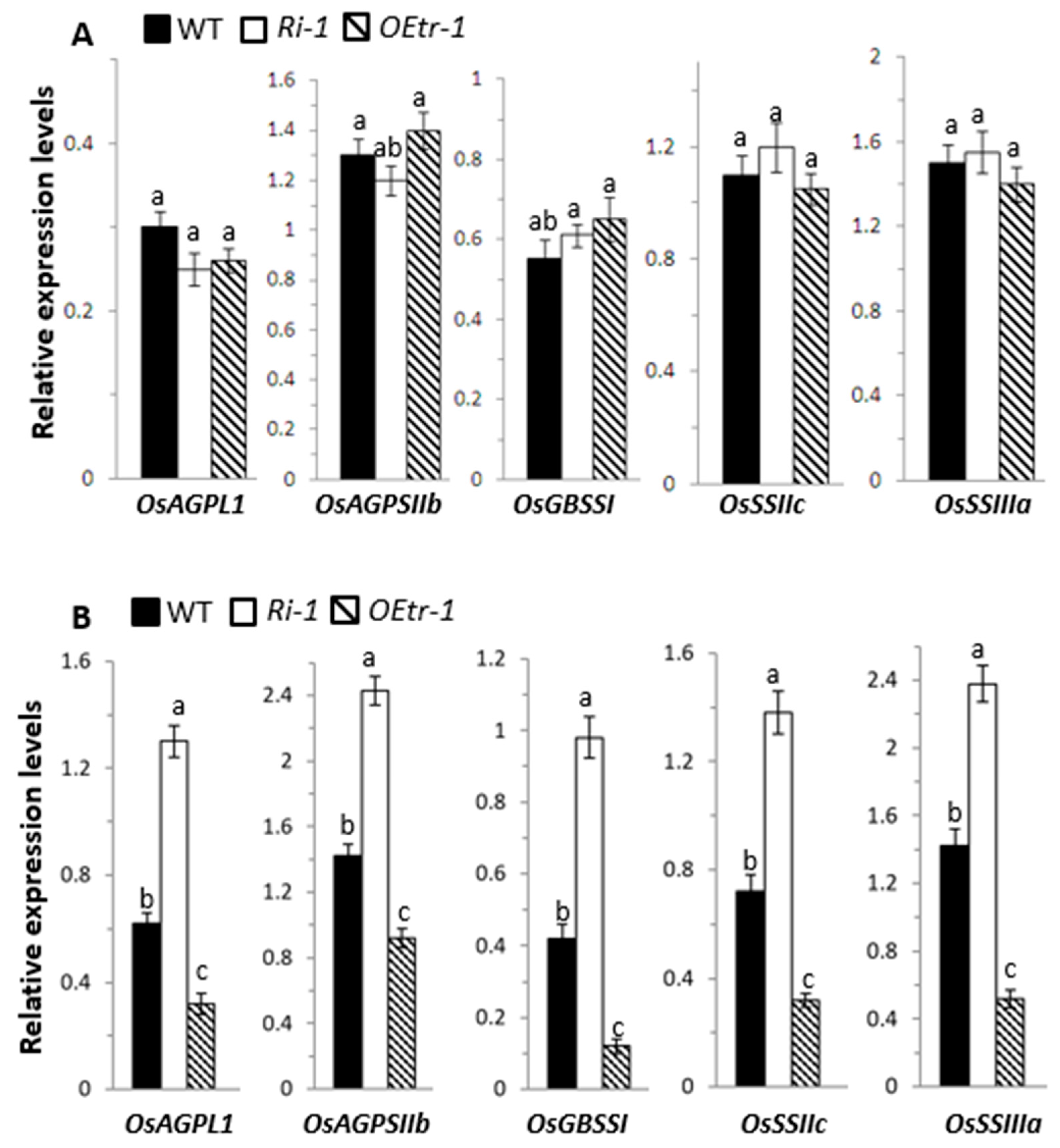
2.6. Effects of OsCDPK1 on the Levels of Starch-Biosynthesis-Related Genes in Developing Rice Seeds
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Callus Induction
4.3. Primers
4.4. Construction of OsCDPK1::GUS Expression Vectors
4.5. Plant Transformation
4.6. Histochemical Staining of GUS Activity in Developing Rice Grains
4.7. Quantitative RT-PCR
4.8. Analysis of the Gelatinization Properties of the Starch
4.9. Apparent Amylose Content Analysis
4.10. Scanning Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rabiei, B.; Valizadeh, M.; Ghareyazie, B.; Moghaddam, M.; Ali, A.J. Identification of QTLs for rice grain size and shape of Iranian cultivars using SSR markers. Euphytica 2004, 137, 325–332. [Google Scholar] [CrossRef]
- Bao, J.; Jin, L.; Xiao, P.; Shen, S.; Sun, M.; Corke, H. Starch physicochemical properties and their associations with microsatellite alleles of starch synthesizing genes in a rice RIL population. J. Agric. Food Chem. 2008, 56, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.A.; Lisle, A.J.; Martin, M. Chalky and translucent rice grains differ in starch composition and structure and cooking properties. Cereal Chem. 2000, 77, 627–632. [Google Scholar]
- Jiang, H.; Dian, W.; Wu, P. Effect of high temperature on fine structure of amylopectin in rice endosperm by reducing the activity of the starch branching enzyme. Phytochemistry 2003, 63, 53–59. [Google Scholar] [CrossRef]
- Morita, S.; Shiratsuchi, H.; Takanashi, J.; Fujita, K. Effect of high temperature on grain ripening in rice plants: Analysis of the effects of high night and high day temperatures applied to the panicle and other parts of the plant. Jpn. J. Crop Sci. 2004, 73, 77–83. [Google Scholar] [CrossRef]
- Counce, P.A.; Bryant, R.J.; Bergman, C.J.; Bautista, R.C.; Wang, Y.J.; Siebenmorgen, T.J.; Moldenhauer, K.A.; Meullenet, J.F.C. Rice milling quality, grain dimensions, and starch branching as affected by high night temperatures. Cereal Chem. 2005, 82, 645–648. [Google Scholar] [CrossRef]
- Patindol, J.; Wang, Y.J. Fine structures and physicochemical properties of starches from chalky and translucent rice kernels. J. Agric. Food Chem. 2003, 51, 2777–2784. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.F.; Xue, H.W. Coexpression analysis identifies rice starch regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator. Plant Physiol. 2010, 154, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Kimura, K.; Arai, Y.; Kawasaki, T.; Shimada, H.; Baba, T. Starch branching enzymes from immature rice seeds. J. Biochem. 1992, 112, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Denyer, K.; Martin, C. The synthesis of the starch granule. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Preiss, J. Starch branching enzymes from maize. Plant Physiol. 1985, 79, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Takeichi, T.; Kawaguchi, K.; Yamanouchi, H. Purification of two forms of starch branching enzyme (Q-enzyme) from developing rice endosperm. Physiol. Plant. 1992, 84, 329–335. [Google Scholar] [CrossRef]
- Satoh, H.; Omura, T. New endosperm mutations induced by chemical mutagens in rice, Oryza sativa L. Jpn. J. Breed. 1981, 31, 316–326. [Google Scholar] [CrossRef]
- Kaushik, R.P.; Khush, G.S. Genetic analysis of endosperm mutants in rice Oryza sativa L. Theor. Appl. Genet. 1991, 83, 146–152. [Google Scholar] [CrossRef] [PubMed]
- She, K.C.; Kusano, H.; Koizumi, K.; Yamakawa, H.; Hakatae, M.; Imamura, T.; Fukuda, M.; Naito, N.; Tsurumaki, Y.; Yaeshima, M.; et al. A novel factor FLOURY ENDOSPERM 2 is involved in regulation of rice grain size and starch quality. Plant Cell 2010, 22, 3280–3294. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Iida, S. Mutant having a low content of 16-kDa allergenic protein in rice (Oryza sativa L. ). Theor. Appl. Genet. 1993, 86, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.G.; Park, S.; Matsuoka, M.; An, G. White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene (OsPPDKB). Plant J. 2005, 42, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, N.; Yu, C.; Park, C.S.; Baik, M.Y.; Park, I.M.; Cho, M.H.; Bhoo, S.H.; An, G.; Hahn, T.R.; Jeon, J.S. Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L.). Plant Cell Rep. 2007, 26, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Y.H.; Liu, F.; Ren, Y.L.; Zhou, K.N.; Lv, J.; Zheng, M.; Zhao, S.L.; Zhang, L.; Wang, C.M.; et al. FLOURY ENDOSPERM6 encodes a CBM48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm. Plant J. 2014, 77, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ren, Y.L.; Lu, B.Y.; Yang, C.Y.; Feng, Z.M.; Liu, Z.; Chen, J.; Ma, W.W.; Wang, Y.H.; Liu, Y.S.; et al. FLOURY ENDOSPERM7 encodes a regulator of starch synthesis and amyloplast development essential for peripheral endosperm development in rice. J. Exp. Bot. 2016, 67, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14, S401–S417. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.F.; Breton, G.; Harmon, A. Decoding Ca2+ signals through plant protein kinases. Annu. Rev. Plant Biol. 2004, 55, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Mori, I.C.; Murata, Y.; Yang, Y.; Munemasa, S.; Wang, Y.F.; Andreoli, S.; Tiriac, H.; Alonso, J.M.; Harper, J.F.; Ecker, J.R.; et al. CDPKs CPK6 and CPK3 function in ABA regulation of guard cell S-type anion- and Ca2+-permeable channels and stomatal closure. PLoS Biol. 2006, 4, e327. [Google Scholar] [CrossRef] [PubMed]
- Sheen, J. Ca2+-dependent protein kinases and stress signal transduction in plants. Science 1996, 274, 1900–1902. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.A.; Romeis, T.; Jones, J.D. CDPK-mediated signalling pathways: Specificity and cross-talk. J. Exp. Bot. 2004, 55, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Romeis, T.; Ludwig, A.A.; Martin, R.; Jones, J.D.G. Calcium-dependent protein kinases play an essential role in a plant defence response. EMBO J. 2001, 20, 5556–5567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi, F.; Onodera, H.; Toki, S.; Tanaka, H.; Komatsu, S. OsCDPK13, a calcium-dependent protein kinase gene from rice, is induced by cold and gibberellin in rice leaf sheath. Plant Mol. Biol. 2004, 55, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Wan, B.; Lin, Y.; Mou, T. Expression of rice Ca2+-dependent protein kinases (CDPKs) genes under different environmental stresses. FEBS Lett. 2007, 581, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Kunieda, N.; Omura, Y.; Ibe, H.; Kawasaki, T.; Takano, M.; Sato, M.; Furuhashi, H.; Mujin, T.; Takaiwa, F.; et al. Rice SPK, a calmodulin-like domain protein kinase, is required for storage product accumulation during seed development: Phosphorylation of sucrose synthase is a possible factor. Plant Cell 2002, 14, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.L.; Huang, L.F.; Lu, C.A.; He, S.L.; Wang, C.C.; Yu, S.P.; Chen, J.; Yu, S.M. Sugar starvation- and GA-inducible calcium-dependent protein kinase 1 feedback regulates GA biosynthesis and activates a 14-3-3 protein to confer drought tolerance in rice seedlings. Plant Mol. Biol. 2013, 81, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. A simplified assay for milled-rice amylose. Cereal Sci. Today 1971, 16, 334–340. [Google Scholar]
- Cameron, D.K.; Wang, Y.J. A better understanding of factors that affect the hardness and stickiness of long-grain rice. Cereal Chem. 2005, 82, 113–119. [Google Scholar] [CrossRef]
- Li, H.; Prakash, S.; Nicholson, T.M.; Fitzgerald, M.A.; Gilbert, R.G. The importance of amylose and amylopectin fine structure for textural properties of cooked rice grains. Food Chem. 2016, 196, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohdan, T.; Francisco, P.B., Jr.; Sawada, T.; Hirose, T.; Terao, T.; Satoh, H.; Nakamura, Y. Expression profiling of genes involved in starch synthesis in sink and source organs of rice. J. Exp. Bot. 2005, 56, 3229–3244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.C.; Xu, H.; Zhu, Y.; Liu, Q.Q.; Ca, X.L. OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. J. Exp. Bot. 2013, 64, 3453–3466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wei, X.; Ren, Y.; Qiu, J.; Jiao, G.; Guo, X.; Tang, S.; Wan, J.; Hu, P. OsBT1 encodes an ADP-glucose transporter involved in starch synthesis and compound granule formation in rice endosperm. Sci. Rep. 2017, 7, 40124. [Google Scholar] [CrossRef] [PubMed]
- Sano, Y.; Maekawa, M.; Kikuchi, H. Temperature effects on the Wx protein level and amylose content in the endosperm of rice. J. Hered. 1985, 6, 221–222. [Google Scholar] [CrossRef]
- Asaoka, M.; Okuno, K.; Sugimoto, Y.; Kawakami, J.; Fuwa, H. Effect of environmental temperature during development of rice plants on some properties of endosperm starch. Starch 1984, 36, 189–193. [Google Scholar] [CrossRef]
- Lyman, N.B.; Jagadish, S.V.K.; Nalley, L.L.; Dixon, B.L.; Siebenmorgen, T. Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress. PLoS ONE 2013, 8, e72157. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Yin, X.; Struik, P.C.; Solis, C.; Xie, F.; Schmidt, R.C.; Huang, M.; Zou, Y.; Ye, C.; Jagadish, S.V.K. High day- and night-time temperatures affect grain growth dynamics in contrasting rice genotypes. J. Exp. Bot. 2017, 68, 5233–5245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakata, M.; Fukamatsu, Y.; Miyashita, T.; Hakata, M.; Kimura, R.; Nakata, Y.; Kuroda, M.; Yamaguchi, T.; Yamakawa, H. High temperature-induced expression of rice α-amylases in developing endosperm produces chalky grains. Front. Plant Sci. 2017, 8, 2089. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.C.; Wang, C.C.; Sun, C.S.; Hsu, C.; Yin, K.C.; Chu, C.Y.; Bi, F.Y. Establishment of an efficient medium for another culture of rice through comparative experiments on the nitrogen sources. Sci. Sin. 1975, 5, 659–668. [Google Scholar]
- Ho, S.L.; Tong, W.F.; Yu, S.M. Multiple mode regulation of a cysteine proteinase gene expression in rice. Plant Physiol. 2000, 122, 57–66. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Plant Height (cm) | Heading Day (day) | Growth Duration (day) | 1000-Grain (g) | Dehulled Grain (mg/grain) | Starch Content (mg/grain) | Amylose Content (%) |
|---|---|---|---|---|---|---|---|
| WT | 108.2 ± 2.5 | 89.3 ± 2.4 | 115.2 ± 3.3 | 23.2 ± 0.6 | 21.4 ± 0.3 | 15.7 ± 0.4 (73.4%) a | 23.2 ± 0.3 |
| OEtr-1 | 99.5 ± 3.5 * | 92.1 ± 2.3 | 117.6 ± 2.7 | 18.2 ± 0.3 ** | 16.8 ± 0.2 ** | 10.5 ± 0.1 ** (62.5%) a | 14.3 ± 0.3 * |
| OEtr-3 | 102.6 ± 2.6 * | 92.4 ± 3.4 * | 119.1 ± 3.8 | 18.6 ± 0.1 ** | 17.3 ± 0.4 ** | 11.8 ± 0.3 ** (68.2%) a | 15.3 ± 0.2 * |
| OEtr-4 | 103. 6 ± 3.1 * | 91.1 ± 2.7 * | 119.4 ± 3.3 | 18.1 ± 0.3 ** | 16.6 ± 0.2 ** | 10.3 ± 0.1 ** (62.0%)a | 15.6 ± 0.4 * |
| Ri-1 | 115.4 ± 4.6 * | 83.6 ± 2.5 ** | 106.6 ± 3.1 * | 25.6 ± 0.3 * | 23.9 ± 0.3 * | 18.7 ± 0.2 * (78.2%)a | 25.8 ± 0.1 * |
| Ri-2 | 118.7 ± 3.8 * | 82.7 ± 2.8 ** | 103.6 ± 2.1 * | 25.3 ± 0.2 * | 23.5 ± 0.2 * | 18.5 ± 0.2 * (78.7%) a | 26.6 ± 0.5 * |
| Ri-3 | 113.6 ± 4.3 * | 83.2 ± 3.4 ** | 104.3 ± 3.6 * | 26.2 ± 0.5 * | 24.4 ± 0.2 * | 18.9 ± 0.6 * (77.5%) a | 26.2 ± 0.2 * |
| Plant Species | Wild Type | OEtr-1 | OEtr-3 | OEtr-4 | Ri-1 | Ri-2 | Ri-3 |
|---|---|---|---|---|---|---|---|
| The ratios of chalky grains in LT (%) | 16.2 ± 2.19 | 100 | 100 | 100 | 6.0 ± 0.61 | 6.7 ± 0.56 | 7.1 ± 0. 61 |
| The ratios of chalky grains in HT (%) | 63.6 ± 5.19 | 100 | 100 | 100 | 50.1 ± 3.61 | 51.2 ± 4.56 | 45.1 ± 3.21 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.-Z.; Kuo, C.-H.; Chen, B.-H.; Chen, M.-K.; Lin, C.-S.; Ho, S.-L. Effects of OsCDPK1 on the Structure and Physicochemical Properties of Starch in Developing Rice Seeds. Int. J. Mol. Sci. 2018, 19, 3247. https://doi.org/10.3390/ijms19103247
Jiang J-Z, Kuo C-H, Chen B-H, Chen M-K, Lin C-S, Ho S-L. Effects of OsCDPK1 on the Structure and Physicochemical Properties of Starch in Developing Rice Seeds. International Journal of Molecular Sciences. 2018; 19(10):3247. https://doi.org/10.3390/ijms19103247
Chicago/Turabian StyleJiang, Jian-Zhi, Chun-Hsiang Kuo, Bo-Hong Chen, Mao-Kei Chen, Choun-Sea Lin, and Shin-Lon Ho. 2018. "Effects of OsCDPK1 on the Structure and Physicochemical Properties of Starch in Developing Rice Seeds" International Journal of Molecular Sciences 19, no. 10: 3247. https://doi.org/10.3390/ijms19103247
APA StyleJiang, J.-Z., Kuo, C.-H., Chen, B.-H., Chen, M.-K., Lin, C.-S., & Ho, S.-L. (2018). Effects of OsCDPK1 on the Structure and Physicochemical Properties of Starch in Developing Rice Seeds. International Journal of Molecular Sciences, 19(10), 3247. https://doi.org/10.3390/ijms19103247