Optimized Clump Culture Methods for Adult Human Multipotent Neural Cells

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
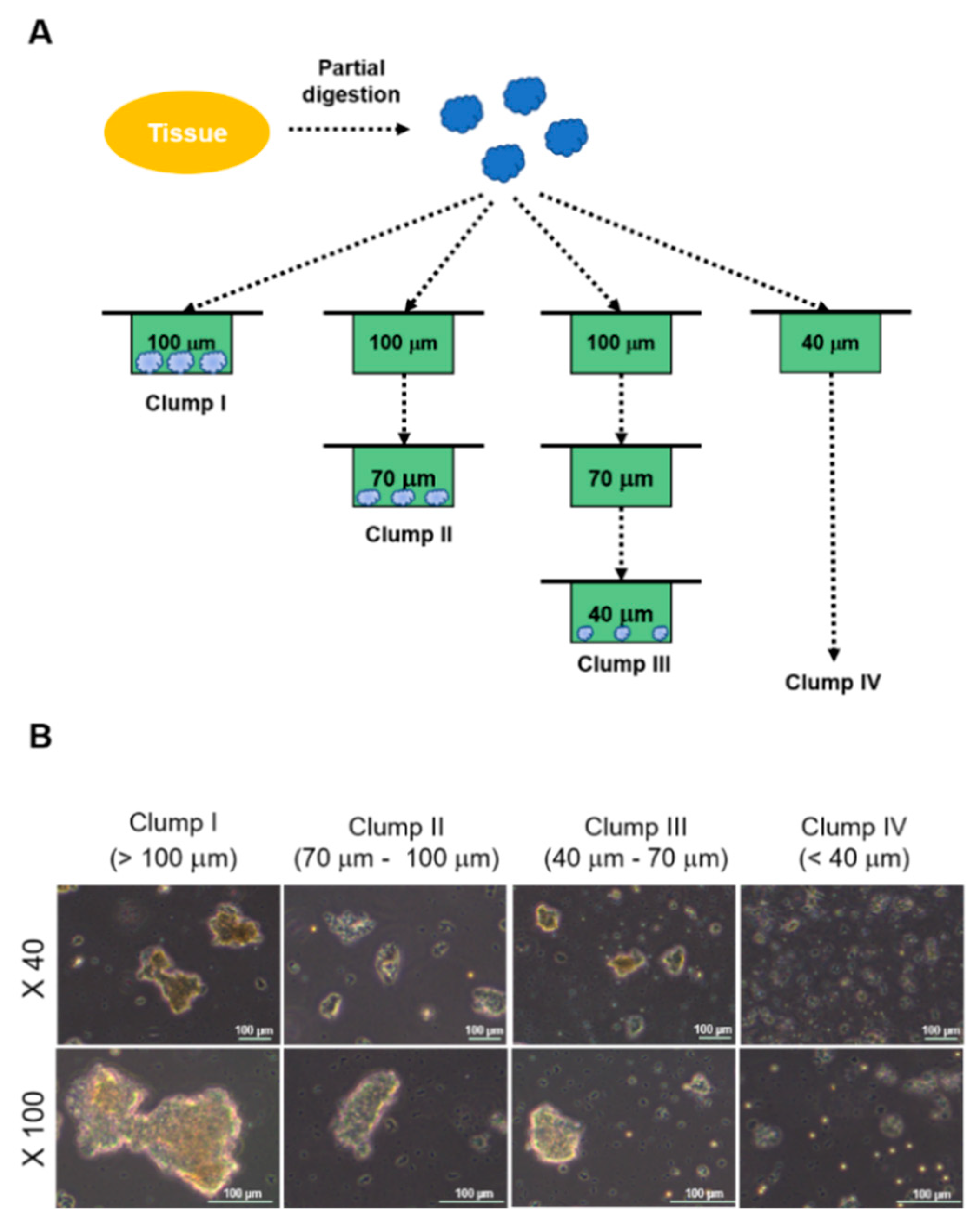
2.1. Experimental Design and Morphologies of Clumps
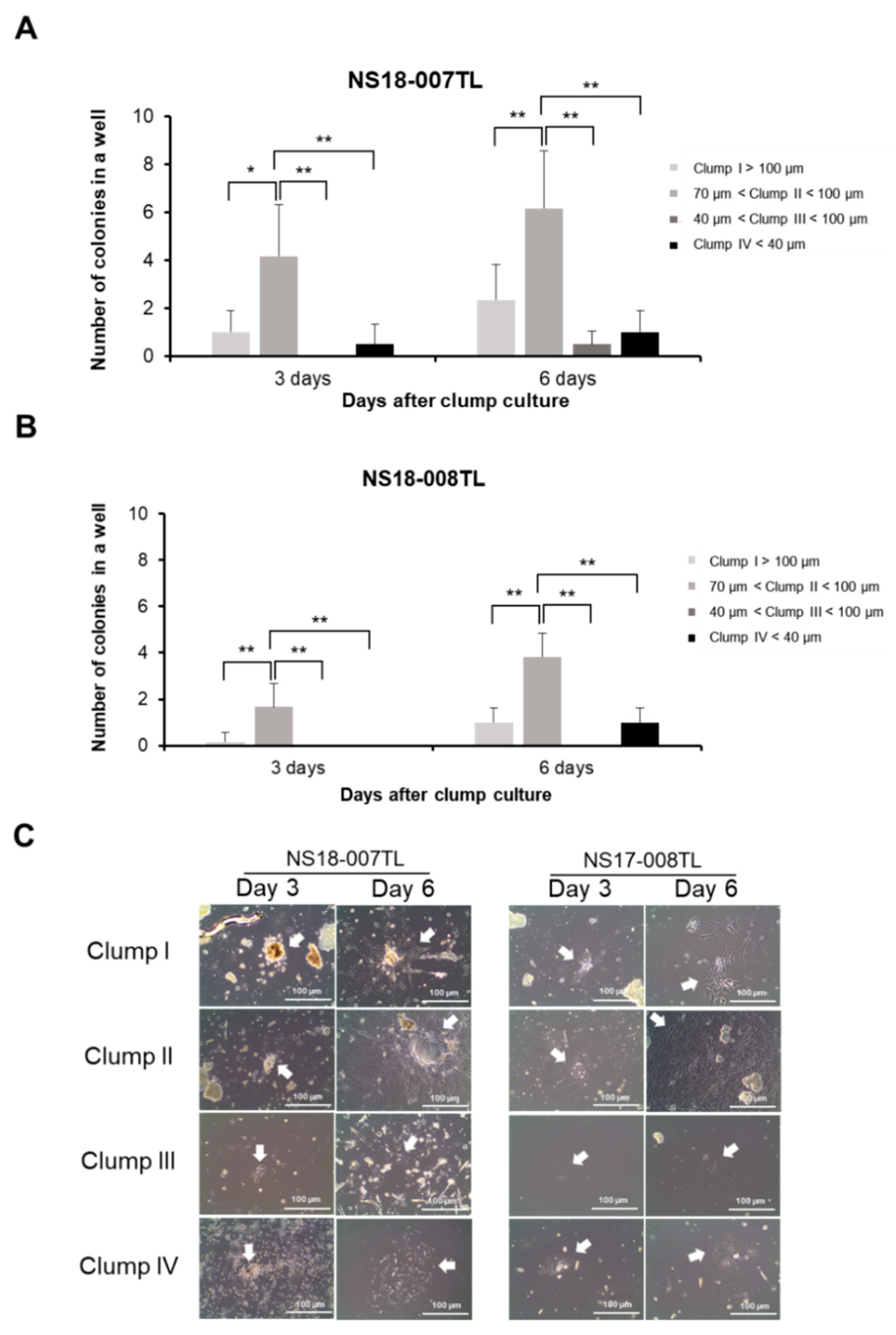
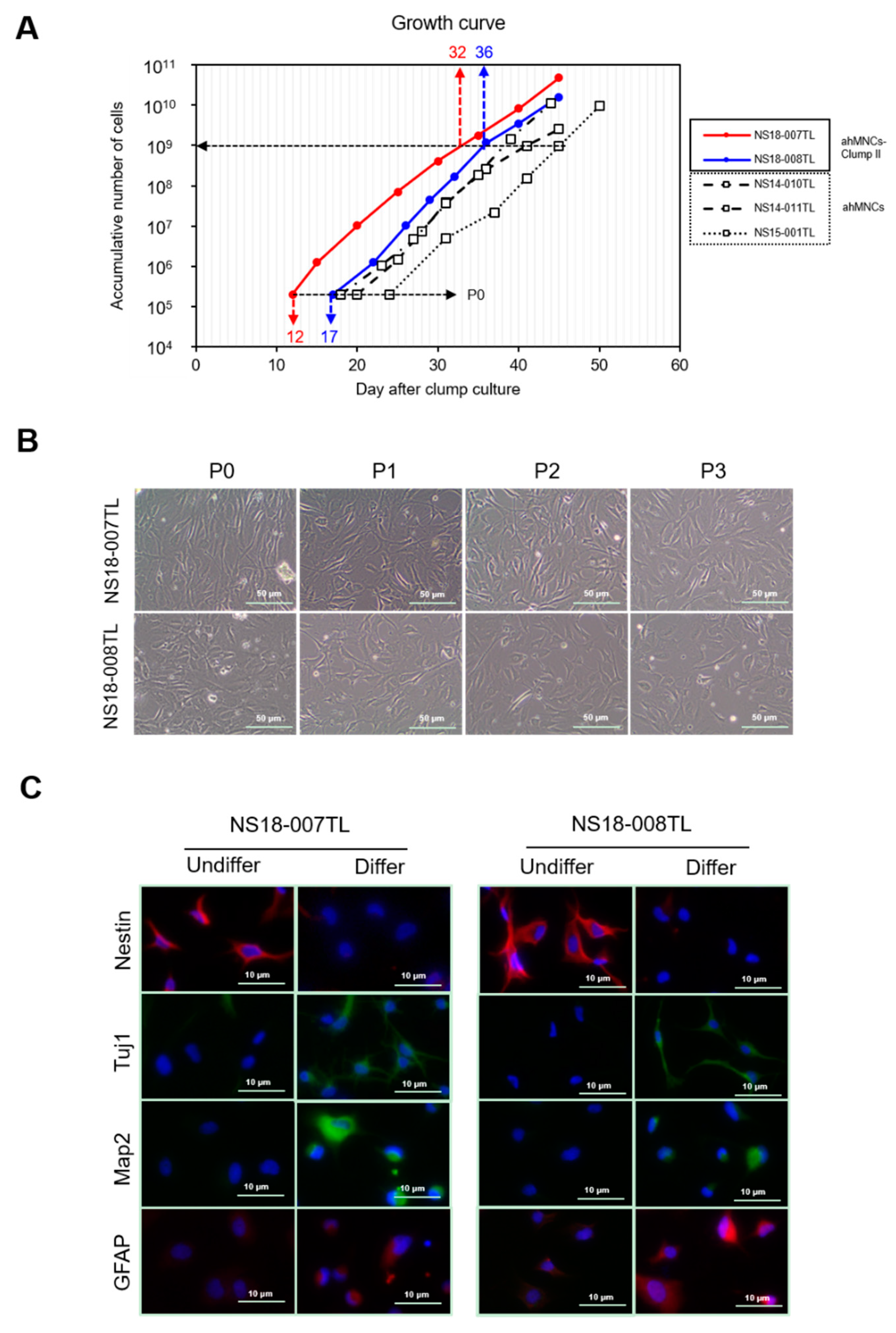
2.2. Optimal Clump Size to Isolate and Expand ahMNCs
2.3. Characterization of ahMNCs-Clump II
2.4. Angiogenic Potential of ahMNCs-Clump II
3. Discussion
4. Materials and Methods
4.1. Study Approval
4.2. Clump Culture
4.3. In Vitro Differentiation
4.4. Immunocytochemistry
4.5. In Vivo Matrigel Plug Assay
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lindvall, O.; Kokaia, Z. Stem cells in human neurodegenerative disorders—Time for clinical translation? J. Clin. Investig. 2010, 120, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.S.; Sakowski, S.A.; Hur, J.; Feldman, E.L. Stem cell technology for neurodegenerative diseases. Ann. Neurol. 2011, 70, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Curtis, E.; Martin, J.R.; Gabel, B.; Sidhu, N.; Rzesiewicz, T.K.; Mandeville, R.; Van Gorp, S.; Leerink, M.; Tadokoro, T.; Marsala, S.; et al. A First-in-Human, Phase I Study of Neural Stem Cell Transplantation for Chronic Spinal Cord Injury. Cell Stem Cell 2018, 22, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Levi, A.D.; Anderson, K.D.; Okonkwo, D.O.; Park, P.; Bryce, T.; Kurpad, S.N.; Aarabi, B.; Hsieh, J.; Gant, K. Clinical Outcomes from a Multi-Center Study of Human Neural Stem Cell Transplantation in Chronic Cervical Spinal Cord Injury. J. Neurotrauma 2018. [Google Scholar] [CrossRef] [PubMed]
- Feldman, E.L.; Boulis, N.M.; Hur, J.; Johe, K.; Rutkove, S.B.; Federici, T.; Polak, M.; Bordeau, J.; Sakowski, S.A.; Glass, J.D. Intraspinal neural stem cell transplantation in amyotrophic lateral sclerosis: Phase 1 trial outcomes. Ann. Neurol. 2014, 75, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalladka, D.; Sinden, J.; Pollock, K.; Haig, C.; McLean, J.; Smith, W.; McConnachie, A.; Santosh, C.; Bath, P.M.; Dunn, L.; et al. Human neural stem cells in patients with chronic ischaemic stroke (PISCES): A phase 1, first-in-man study. Lancet 2016, 388, 787–796. [Google Scholar] [CrossRef]
- Madhavan, L.; Collier, T.J. A synergistic approach for neural repair: Cell transplantation and induction of endogenous precursor cell activity. Neuropharmacology 2010, 58, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Deng, W.; Gage, F.H. Mechanisms and functional implications of adult neurogenesis. Cell 2008, 132, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Ostenfeld, T.; Joly, E.; Tai, Y.T.; Peters, A.; Caldwell, M.; Jauniaux, E.; Svendsen, C.N. Regional specification of rodent and human neurospheres. Brain Res. Dev. Brain Res. 2002, 134, 43–55. [Google Scholar] [CrossRef]
- Schwartz, P.H.; Bryant, P.J.; Fuja, T.J.; Su, H.; O’Dowd, D.K.; Klassen, H. Isolation and characterization of neural progenitor cells from post-mortem human cortex. J. Neurosci. Res. 2003, 74, 838–851. [Google Scholar] [CrossRef] [PubMed]
- Piao, J.H.; Odeberg, J.; Samuelsson, E.B.; Kjaeldgaard, A.; Falci, S.; Seiger, A.; Sundstrom, E.; Akesson, E. Cellular composition of long-term human spinal cord- and forebrain-derived neurosphere cultures. J. Neurosci. Res. 2006, 84, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Vescovi, A.L.; Parati, E.A.; Gritti, A.; Poulin, P.; Ferrario, M.; Wanke, E.; Frolichsthal-Schoeller, P.; Cova, L.; Arcellana-Panlilio, M.; Colombo, A.; et al. Isolation and cloning of multipotential stem cells from the embryonic human CNS and establishment of transplantable human neural stem cell lines by epigenetic stimulation. Exp. Neurol. 1999, 156, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.S.; Nakano, T.; Keyoung, H.M.; Windrem, M.; Rashbaum, W.K.; Alonso, M.L.; Kang, J.; Peng, W.; Carpenter, M.K.; Lin, J.; et al. Telomerase immortalization of neuronally restricted progenitor cells derived from the human fetal spinal cord. Nat. Biotechnol. 2004, 22, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Quinn, S.M.; Walters, W.M.; Vescovi, A.L.; Whittemore, S.R. Lineage restriction of neuroepithelial precursor cells from fetal human spinal cord. J. Neurosci. Res. 1999, 57, 590–602. [Google Scholar] [CrossRef]
- Barami, K.; Zhao, J.; Diaz, F.G.; Lyman, W.D. Comparison of neural precursor cell fate in second trimester human brain and spinal cord. Neurol. Res. 2001, 23, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, M.A.; He, X.; Wilkie, N.; Pollack, S.; Marshall, G.; Wafford, K.A.; Svendsen, C.N. Growth factors regulate the survival and fate of cells derived from human neurospheres. Nat. Biotechnol. 2001, 19, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.K.; Cui, X.; Hu, Z.Y.; Jackson, J.; Sherman, S.; Seiger, A.; Wahlberg, L.U. In vitro expansion of a multipotent population of human neural progenitor cells. Exp. Neurol. 1999, 158, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, C.N.; ter Borg, M.G.; Armstrong, R.J.; Rosser, A.E.; Chandran, S.; Ostenfeld, T.; Caldwell, M.A. A new method for the rapid and long term growth of human neural precursor cells. J. Neurosci. Methods 1998, 85, 141–152. [Google Scholar] [CrossRef]
- Akesson, E.; Piao, J.H.; Samuelsson, E.B.; Holmberg, L.; Kjaeldgaard, A.; Falci, S.; Sundstrom, E.; Seiger, A. Long-term culture and neuronal survival after intraspinal transplantation of human spinal cord-derived neurospheres. Physiol. Behav. 2007, 92, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Joo, K.M.; Kang, B.G.; Yeon, J.Y.; Cho, Y.J.; An, J.Y.; Song, H.S.; Won, J.H.; Kim, S.J.; Hong, S.C.; Nam, D.H. Experimental and clinical factors influencing long-term stable in vitro expansion of multipotent neural cells from human adult temporal lobes. Exp. Neurol. 2013, 240, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Pyeon, H.J.; Nam, H.; Won, J.S.; Hwang, J.Y.; Lee, K.A.; Yeon, J.Y.; Hong, S.C.; Nam, D.H.; Lee, K.; et al. Significant therapeutic effects of adult human multipotent neural cells on spinal cord injury. Stem Cell Res. 2018, 31, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Nam, H.; Jeong da, E.; Kim, S.S.; Song, H.J.; Pyeon, H.J.; Kang, K.; Hong, S.C.; Nam, D.H.; Joo, K.M. Sensitive Tumorigenic Potential Evaluation of Adult Human Multipotent Neural Cells Immortalized by hTERT Gene Transduction. PLoS ONE 2016, 11, e0158639. [Google Scholar] [CrossRef] [PubMed]
- Moe, M.C.; Westerlund, U.; Varghese, M.; Berg-Johnsen, J.; Svensson, M.; Langmoen, I.A. Development of neuronal networks from single stem cells harvested from the adult human brain. Neurosurgery 2005, 56, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Doetsch, F. A niche for adult neural stem cells. Curr. Opin. Genet. Dev. 2003, 13, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Doetsch, F.; Caille, I.; Lim, D.A.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 1999, 97, 703–716. [Google Scholar] [CrossRef]
- Lois, C.; Alvarez-Buylla, A. Proliferating subventricular zone cells in the adult mammalian forebrain can differentiate into neurons and glia. Proc. Natl. Acad. Sci. USA 1993, 90, 2074–2077. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.D.; Schwartz, P.H.; Taupin, P.; Kaspar, B.; Stein, S.A.; Gage, F.H. Cell culture. Progenitor cells from human brain after death. Nature 2001, 411, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.C.; Roy, N.S.; Keyoung, H.M.; Goodman, R.R.; McKhann, G.; Jiang, L.; Kang, J.; Nedergaard, M.; Goldman, S.A. Identification and isolation of multipotential neural progenitor cells from the subcortical white matter of the adult human brain. Nat. Med. 2003, 9, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Moe, M.C.; Varghese, M.; Danilov, A.I.; Westerlund, U.; Ramm-Pettersen, J.; Brundin, L.; Svensson, M.; Berg-Johnsen, J.; Langmoen, I.A. Multipotent progenitor cells from the adult human brain: Neurophysiological differentiation to mature neurons. Brain 2005, 128, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Wang, X.; Xie, L.; Mao, X.O.; Zhu, W.; Wang, Y.; Shen, J.; Mao, Y.; Banwait, S.; Greenberg, D.A. Evidence for stroke-induced neurogenesis in the human brain. Proc. Natl. Acad. Sci. USA 2006, 103, 13198–13202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsenijevic, Y.; Villemure, J.G.; Brunet, J.F.; Bloch, J.J.; Deglon, N.; Kostic, C.; Zurn, A.; Aebischer, P. Isolation of multipotent neural precursors residing in the cortex of the adult human brain. Exp. Neurol. 2001, 170, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Ayuso-Sacido, A.; Roy, N.S.; Schwartz, T.H.; Greenfield, J.P.; Boockvar, J.A. Long-term expansion of adult human brain subventricular zone precursors. Neurosurgery 2008, 62, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, B.A.; Weiss, S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C.B.; Svensson, M.; Wallstedt, L.; Janson, A.M.; Frisen, J. Neural stem cells in the adult human brain. Exp. Cell Res. 1999, 253, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Kukekov, V.G.; Laywell, E.D.; Suslov, O.; Davies, K.; Scheffler, B.; Thomas, L.B.; O’Brien, T.F.; Kusakabe, M.; Steindler, D.A. Multipotent stem/progenitor cells with similar properties arise from two neurogenic regions of adult human brain. Exp. Neurol. 1999, 156, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Westerlund, U.; Moe, M.C.; Varghese, M.; Berg-Johnsen, J.; Ohlsson, M.; Langmoen, I.A.; Svensson, M. Stem cells from the adult human brain develop into functional neurons in culture. Exp. Cell Res. 2003, 289, 378–383. [Google Scholar] [CrossRef]
- Westerlund, U.; Svensson, M.; Moe, M.C.; Varghese, M.; Gustavsson, B.; Wallstedt, L.; Berg-Johnsen, J.; Langmoen, I.A. Endoscopically harvested stem cells: A putative method in future autotransplantation. Neurosurgery 2005, 57, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Walton, N.M.; Sutter, B.M.; Chen, H.X.; Chang, L.J.; Roper, S.N.; Scheffler, B.; Steindler, D.A. Derivation and large-scale expansion of multipotent astroglial neural progenitors from adult human brain. Development 2006, 133, 3671–3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, M.; Olstorn, H.; Berg-Johnsen, J.; Moe, M.C.; Murrell, W.; Langmoen, I.A. Isolation of human multipotent neural progenitors from adult filum terminale. Stem Cells Dev. 2009, 18, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Lojewski, X.; Srimasorn, S.; Rauh, J.; Francke, S.; Wobus, M.; Taylor, V.; Arauzo-Bravo, M.J.; Hallmeyer-Elgner, S.; Kirsch, M.; Schwarz, S.; et al. Perivascular Mesenchymal Stem Cells From the Adult Human Brain Harbor No Instrinsic Neuroectodermal but High Mesodermal Differentiation Potential. Stem Cells Transl. Med. 2015, 4, 1223–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozen, I.; Boix, J.; Paul, G. Perivascular mesenchymal stem cells in the adult human brain: A future target for neuroregeneration? Clin. Transl. Med. 2012, 1, 30. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.; Ozen, I.; Christophersen, N.S.; Reinbothe, T.; Bengzon, J.; Visse, E.; Jansson, K.; Dannaeus, K.; Henriques-Oliveira, C.; Roybon, L.; et al. The adult human brain harbors multipotent perivascular mesenchymal stem cells. PLoS ONE 2012, 7, e35577. [Google Scholar] [CrossRef] [PubMed]
- Maslov, A.Y.; Barone, T.A.; Plunkett, R.J.; Pruitt, S.C. Neural stem cell detection, characterization, and age-related changes in the subventricular zone of mice. J. Neurosci. 2004, 24, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Simian, M.; Bissell, M.J. Organoids: A historical perspective of thinking in three dimensions. J. Cell Biol. 2017, 216, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Melero-Martin, J.M.; De Obaldia, M.E.; Kang, S.Y.; Khan, Z.A.; Yuan, L.; Oettgen, P.; Bischoff, J. Engineering robust and functional vascular networks in vivo with human adult and cord blood-derived progenitor cells. Circ. Res. 2008, 103, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Melero-Martin, J.M.; Khan, Z.A.; Picard, A.; Wu, X.; Paruchuri, S.; Bischoff, J. In vivo vasculogenic potential of human blood-derived endothelial progenitor cells. Blood 2007, 109, 4761–4768. [Google Scholar] [CrossRef] [PubMed] [Green Version]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeon, J.Y.; Hwang, J.-Y.; Lee, H.W.; Pyeon, H.-J.; Won, J.-S.; Noh, Y.-J.; Nam, H.; Joo, K.M. Optimized Clump Culture Methods for Adult Human Multipotent Neural Cells. Int. J. Mol. Sci. 2018, 19, 3380. https://doi.org/10.3390/ijms19113380
Yeon JY, Hwang J-Y, Lee HW, Pyeon H-J, Won J-S, Noh Y-J, Nam H, Joo KM. Optimized Clump Culture Methods for Adult Human Multipotent Neural Cells. International Journal of Molecular Sciences. 2018; 19(11):3380. https://doi.org/10.3390/ijms19113380
Chicago/Turabian StyleYeon, Je Young, Ji-Yoon Hwang, Hye Won Lee, Hee-Jang Pyeon, Jeong-Seob Won, Yoo-Jung Noh, Hyun Nam, and Kyeung Min Joo. 2018. "Optimized Clump Culture Methods for Adult Human Multipotent Neural Cells" International Journal of Molecular Sciences 19, no. 11: 3380. https://doi.org/10.3390/ijms19113380
APA StyleYeon, J. Y., Hwang, J. -Y., Lee, H. W., Pyeon, H. -J., Won, J. -S., Noh, Y. -J., Nam, H., & Joo, K. M. (2018). Optimized Clump Culture Methods for Adult Human Multipotent Neural Cells. International Journal of Molecular Sciences, 19(11), 3380. https://doi.org/10.3390/ijms19113380