Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis


Abstract
1. Introduction
2. Results
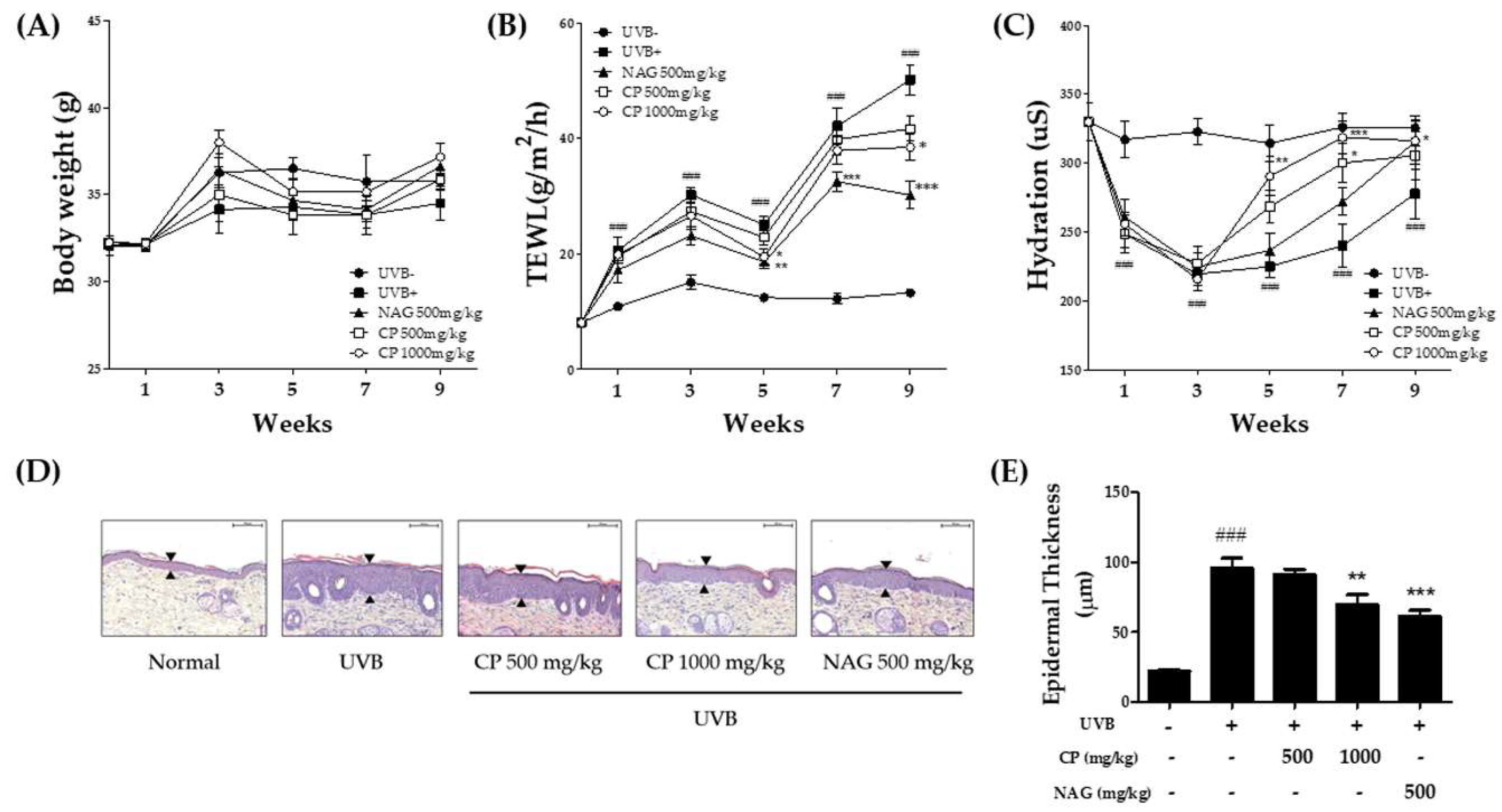
2.1. Changes in Transepidermal Water Loss, Skin Hydration, and Epidermal Thickness in the Skin of Hairless Mice Treated with Collagen Peptide
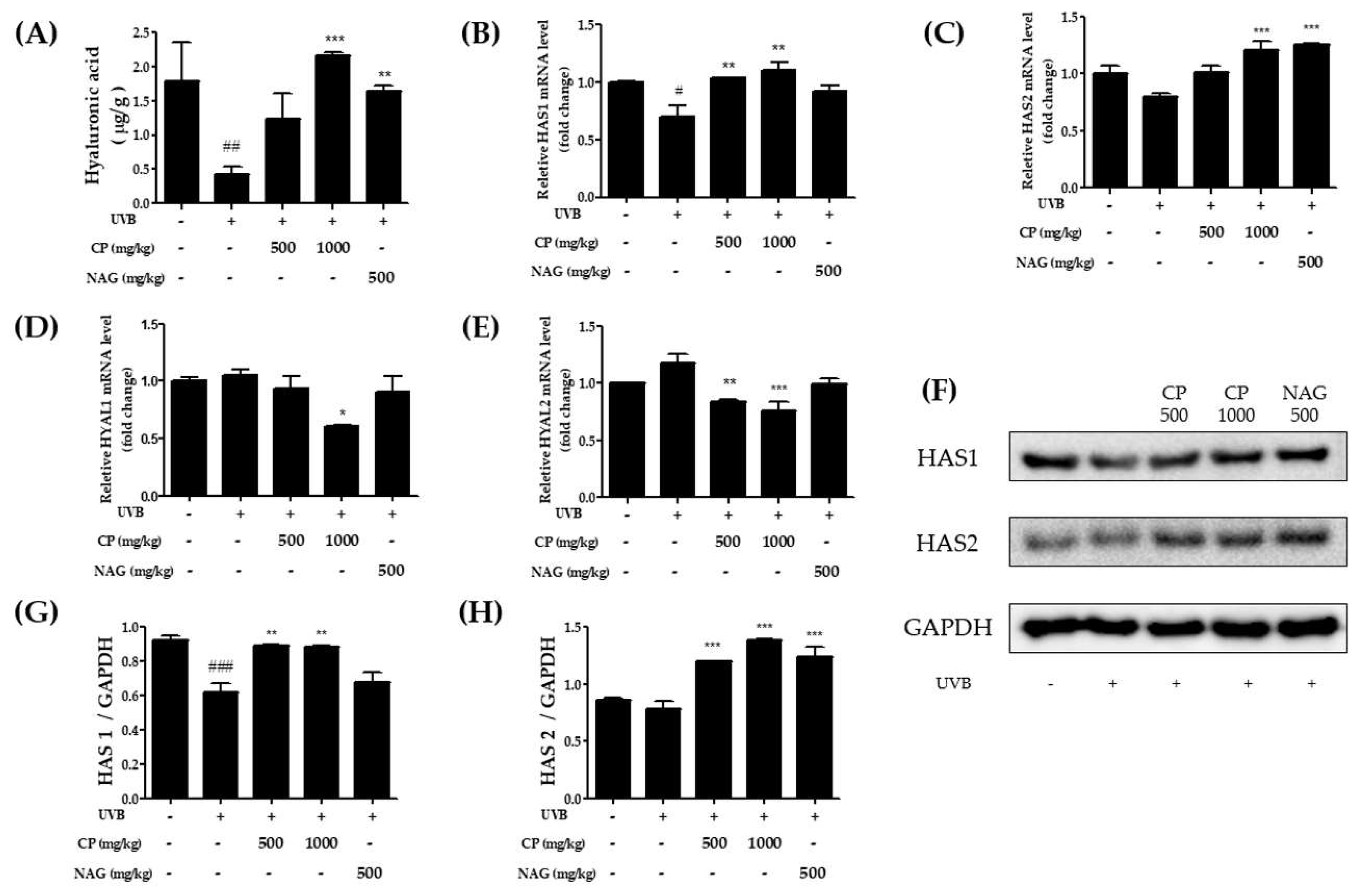
2.2. Hyaluronic Acid Production and HAS and HYAL mRNA and Protein Expression in the Skin Tissue of UV-Irradiated Hairless Mice Treated with Collagen Peptide
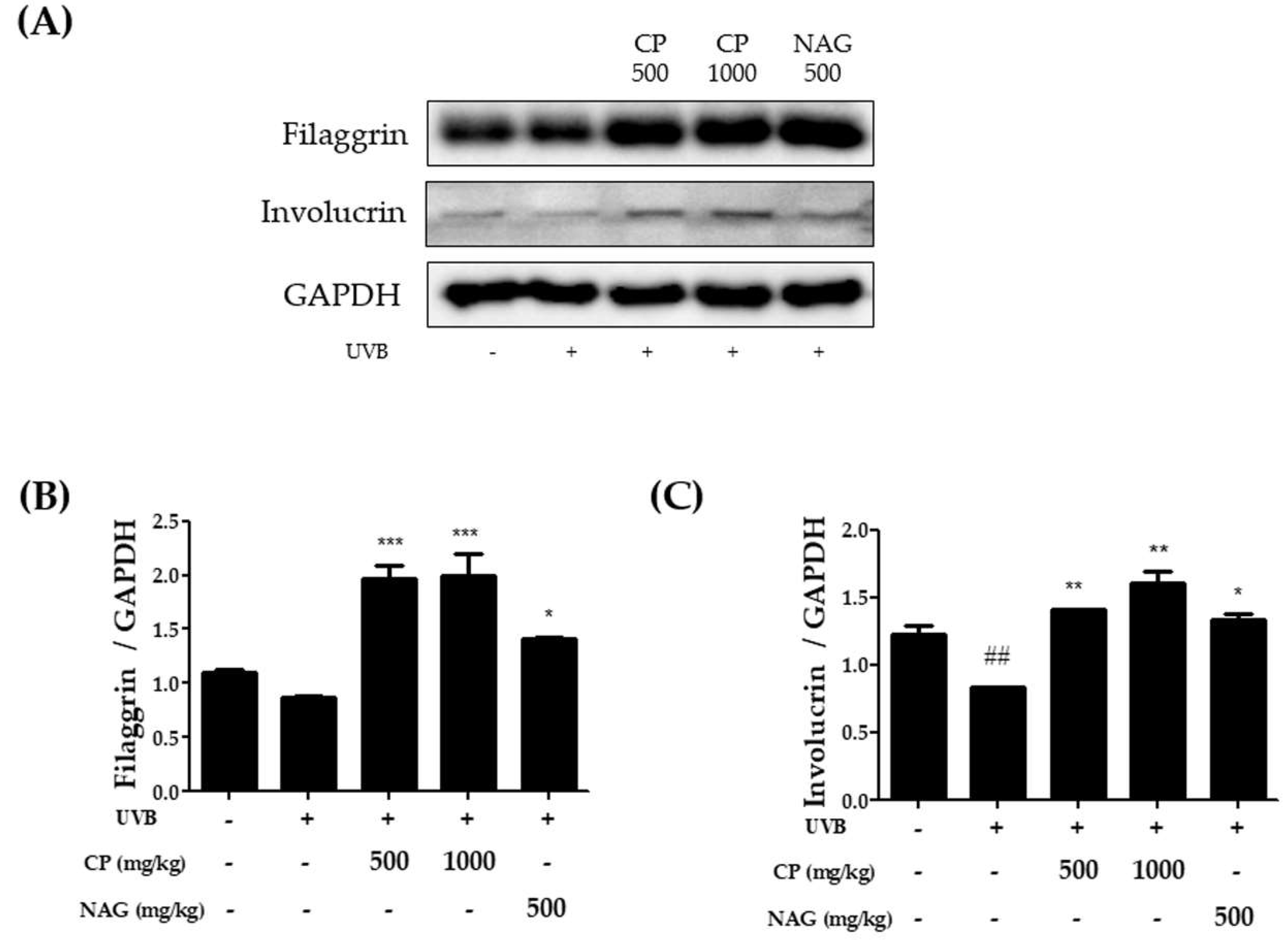
2.3. Expression of Skin Barrier Function-Related Proteins in the Dorsal Skin of Hairless Mice Treated with Collagen Peptide
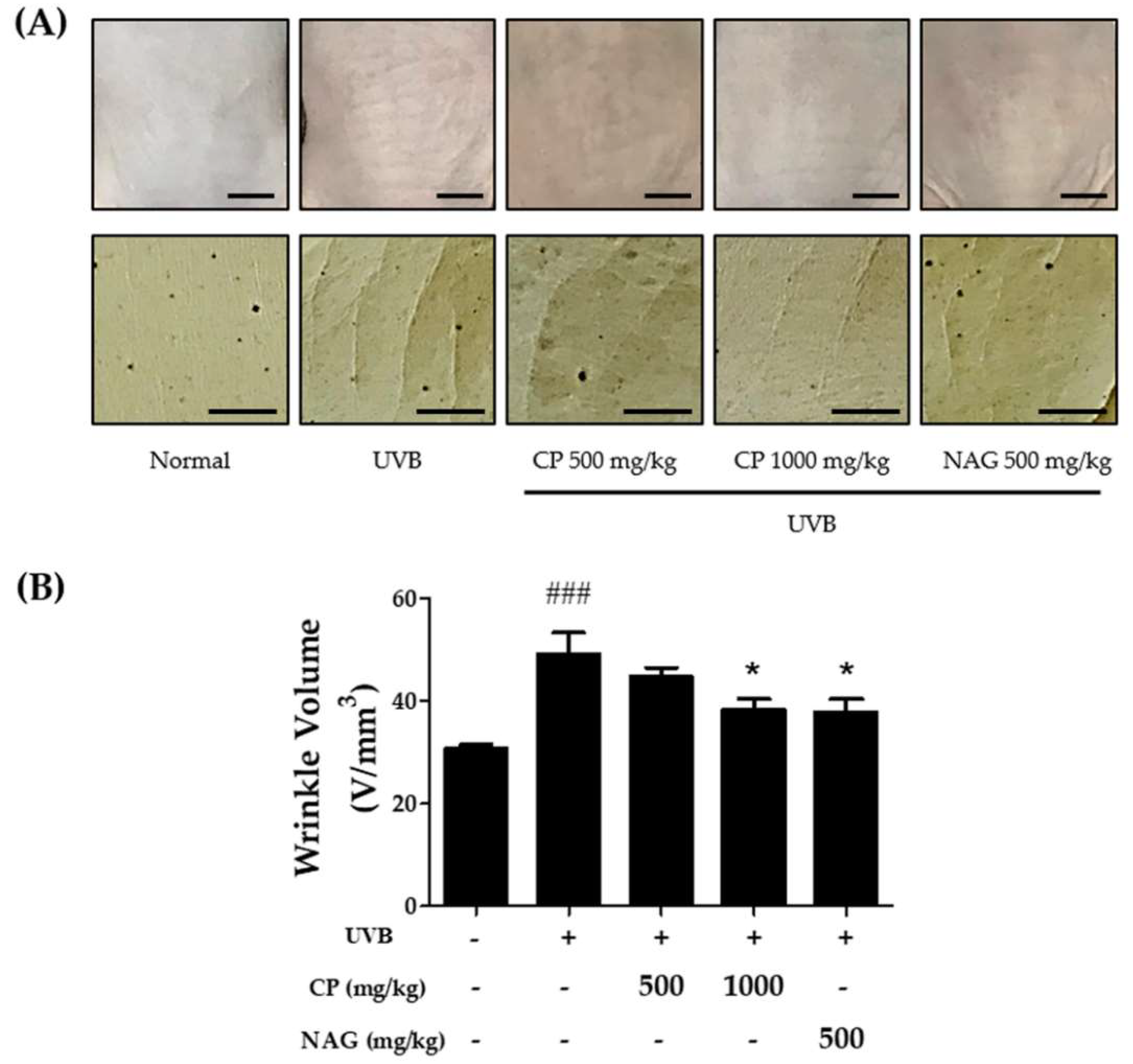
2.4. Changes in Wrinkle Formation in the Dorsal Skin of Hairless Mice Treated with Collagen Peptide
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. UVB Radiation
4.3. Treatment
4.4. Evaluation of Skin Barrier Function
4.5. Wrinkle Measurement
4.6. Western Blot Analysis
4.7. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
4.8. Histological Skin Analysis
4.9. Extraction and Quantification of Hyaluronic Acid
4.10. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV Radiation and the Skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.F.; Chen, Y.Q.; Kopp, J.B.; Fisher, L.; Brown, D.B.; Hahn, P.J.; Robey, F.A.; Lakkakorpi, J.; Uitto, J. Long-term sun exposure alters the collagen of the papillary dermis. Comparison of sun-protected and photoaged skin by northern analysis, immunohistochemical staining, and confocal laser scanning microscopy. J. Am. Acad. Dermatol. 1996, 34, 209–218. [Google Scholar] [CrossRef]
- Imokawa, G. Recent advances in characterizing biological mechanisms underlying UV-induced wrinkles: A pivotal role of fibrobrast-derived elastase. Arch. Dermatol. Res. 2008, 300, S7–S20. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.R.; Oh, C.T.; Choi, E.J.; Kim, S.R.; Jang, Y.J.; Ko, E.J.; Yoo, K.H.; Kim, B.J. Conditioned medium from human bone marrow-derived mesenchymal stem cells promotes skin moisturization and effacement of wrinkles in UVB-irradiated SKH-1 hairless mice. Photodermatol. Photoimmunol. Photomed. 2016, 32, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Boelsma, E.; van de Vijver, L.P.; Goldbohm, R.A.; Klopping-Ketelaars, I.A.; Hendriks, H.F.; Roza, L. Human skin condition and its associations with nutrient concentrations in serum and diet. Am. J. Clin. Nutr. 2003, 77, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Berillis, P. Marine collagen: Extraction and applications. In Research Trends in Biochemistry, Molecular Biology and Microbiology; 2015; pp. 1–13. Available online: http://www.smgebooks.com/research-trends-biochemistry-molecular-biology-microbiology/chapters/RTBMBM-15-02.pdf (accessed on 5 April 2015).
- Gómez-Guillén, M.; Giménez, B.; López-Caballero, M.a.; Montero, M. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocol. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Spandana, V.; Poonam, T. Extraction and Determination of Collagen Peptide and Its Clinical Importance from Tilapia Fish Scales (Oreochromis niloticus). 2011. Available online: http://agris.fao.org/agris-search/search.do?recordID=AV2012074699 (accessed on 10 November 2018).
- Ichikawa, S.; Morifuji, M.; Ohara, H.; Matsumoto, H.; Takeuchi, Y.; Sato, K. Hydroxyproline-containing dipeptides and tripeptides quantified at high concentration in human blood after oral administration of gelatin hydrolysate. Int. J. Food Sci. Nutr. 2010, 61, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Hasegawa, T.; Taguchi, Y.; Morimatsu, F.; Sato, K.; Nakamura, Y.; Higashi, A.; Kido, Y.; Nakabo, Y.; Ohtsuki, K. Identification of food-derived collagen peptides in human blood after oral ingestion of gelatin hydrolysates. J. Agric. Food Chem. 2005, 53, 6531–6536. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Segger, D.; Degwert, J.; Schunck, M.; Zague, V.; Oesser, S. Oral supplementation of specific collagen peptides has beneficial effects on human skin physiology: A double-blind, placebo-controlled study. Skin Pharmacol. Physiol. 2014, 27, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Oesser, S.; Adam, M.; Babel, W.; Seifert, J. Oral administration of 14C labeled gelatin hydrolysate leads to an accumulation of radioactivity in cartilage of mice (C57/BL). J. Nutr. 1999, 129, 1891–1895. [Google Scholar] [CrossRef] [PubMed]
- Shigemura, Y.; Iwai, K.; Morimatsu, F.; Iwamoto, T.; Mori, T.; Oda, C.; Taira, T.; Park, E.Y.; Nakamura, Y.; Sato, K. Effect of Prolyl-hydroxyproline (Pro-Hyp), a food-derived collagen peptide in human blood, on growth of fibroblasts from mouse skin. J. Agric. Food Chem. 2009, 57, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yang, P.; Zhou, C.; Li, S.; Hong, P. Marine collagen peptides from the skin of Nile Tilapia (Oreochromis niloticus): Characterization and wound healing evaluation. Marine Drugs 2017, 15, 102. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Lee, J.-H.; Bae, I.-H.; Seo, D.-B.; Lee, S.-J. Beneficial effect of a collagen peptide supplement on the epidermal skin barrier. Korean J. Food Sci. Technol. 2011, 43, 458–463. [Google Scholar] [CrossRef]
- Zague, V.; de Freitas, V.; da Costa Rosa, M.; de Castro, G.A.; Jaeger, R.G.; Machado-Santelli, G.M. Collagen hydrolysate intake increases skin collagen expression and suppresses matrix metalloproteinase 2 activity. J. Med. Food 2011, 14, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Puizina-Ivic, N. Skin aging. Acta Dermatovenerol Alp Pannonica Adriat 2008, 17, 47–54. [Google Scholar] [PubMed]
- Berneburg, M.; Plettenberg, H.; Krutmann, J. Photoaging of human skin. Photodermatol. Photoimmunol. Photomed. 2000, 16, 239–244. [Google Scholar] [CrossRef]
- Verdier-Sevrain, S.; Bonte, F. Skin hydration: A review on its molecular mechanisms. J. Cosmet. Dermatol. 2007, 6, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Wiest, L.; Kerscher, M. Native hyaluronic acid in dermatology—Results of an expert meeting. J. German. Soc. Dermatol. 2008, 6, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Int. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef]
- Sakai, S.; Yasuda, R.; Sayo, T.; Ishikawa, O.; Inoue, S. Hyaluronan exists in the normal stratum corneum. J. Investig. Dermatol. 2000, 114, 1184–1187. [Google Scholar] [CrossRef] [PubMed]
- Itano, N.; Kimata, K. Mammalian hyaluronan synthases. IUBMB Life 2002, 54, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Csoka, A.B.; Frost, G.I.; Stern, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biol. 2001, 20, 499–508. [Google Scholar] [CrossRef]
- Dai, G.; Freudenberger, T.; Zipper, P.; Melchior, A.; Grether-Beck, S.; Rabausch, B.; de Groot, J.; Twarock, S.; Hanenberg, H.; Homey, B.; et al. Chronic ultraviolet B irradiation causes loss of hyaluronic acid from mouse dermis because of down-regulation of hyaluronic acid synthases. Am. J. Pathol. 2007, 171, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.V. Molecular basis for stratum corneum maturation and moisturization. Br. J. Dermatol. 2014, 171, 19–28. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.A.; Uitto, J. The filaggrin story: Novel insights into skin-barrier function and disease. Trends Mol. Med. 2008, 14, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Nishifuji, K.; Yoon, J.S. The stratum corneum: The rampart of the mammalian body. Vet. Dermatol. 2013, 24, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, J.E.; Suk, S.; Kwon, O.W.; Park, G.; Lim, T.G.; Seo, S.G.; Kim, J.R.; Kim, D.E.; Lee, M.; et al. A fermented barley and soybean formula enhances skin hydration. J. Clin. Biochem. Nutr. 2015, 57, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Del Bino, S.; Vioux, C.; Rossio-Pasquier, P.; Jomard, A.; Demarchez, M.; Asselineau, D.; Bernerd, F. Ultraviolet B induces hyperproliferation and modification of epidermal differentiation in normal human skin grafted on to nude mice. Br. J. Dermatol. 2004, 150, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, S.N.A.; Roswandi, N.L.; Waqas, M.; Habib, H.; Hussain, F.; Khan, S.; Sohail, M.; Ramli, N.A.; Thu, H.E.; Hussain, Z. Hyaluronic acid, a promising skin rejuvenating biomedicine: A review of recent updates and pre-clinical and clinical investigations on cosmetic and nutricosmetic effects. Int. J. Biol. Macromol. 2018, 120, 1682–1695. [Google Scholar] [CrossRef] [PubMed]
- Croce, M.A.; Dyne, K.; Boraldi, F.; Quaglino, D., Jr.; Cetta, G.; Tiozzo, R.; Pasquali Ronchetti, I. Hyaluronan affects protein and collagen synthesis by in vitro human skin fibroblasts. Tissue Cell 2001, 33, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Matahira, Y. Oral N-acetylglucosamine supplementation improves skin conditions of female volunteers: Clinical evaluation by a microscopic three-dimensional skin surface analyzer. J. Appl. Cosmetol. 2002, 20, 143–152. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | UVB | Sample Dose |
|---|---|---|
| Normal | − | − |
| UVB | + | − |
| CP 500 | + | 500 mg/kg |
| CP 1000 | + | 1000 mg/kg |
| NAG | + | 500 mg/kg |
| Gene | Sequence |
|---|---|
| HAS1 | forward: 5′-CTATGCTACCAAGTATACCTCG-3′ |
| reverse: 5′-TCTCGGAAGTAAGATTTGGAC-3′ | |
| HAS2 | forward: 5′-GAGCACCAAGGTTCTGCTTC-3′ |
| reverse: 5′-CTCTCCATACGGCGAGAGTC-3′ | |
| Hyal 1 | forward: 5′-TCATCGTGAACGTGACCAGT-3′ |
| reverse: 5′-GAGAGCCTCAGGATAACTTGGATG-3′ | |
| Hyal 2 | forward: 5′-GCAGGACTAGGTCCCATCATC-3′ |
| reverse: 5′-TTCCATGCTACCACAAAGGGT-3′ | |
| GAPDH | forward: 5′-CCATGGAGAAGGCTGGGG-3′ |
| reverse: 5′-CAAAGTTGTCATGGATGACC-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, M.C.; Yumnam, S.; Kim, S.Y. Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. Int. J. Mol. Sci. 2018, 19, 3551. https://doi.org/10.3390/ijms19113551
Kang MC, Yumnam S, Kim SY. Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. International Journal of Molecular Sciences. 2018; 19(11):3551. https://doi.org/10.3390/ijms19113551
Chicago/Turabian StyleKang, Min Cheol, Silvia Yumnam, and Sun Yeou Kim. 2018. "Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis" International Journal of Molecular Sciences 19, no. 11: 3551. https://doi.org/10.3390/ijms19113551
APA StyleKang, M. C., Yumnam, S., & Kim, S. Y. (2018). Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. International Journal of Molecular Sciences, 19(11), 3551. https://doi.org/10.3390/ijms19113551