Inadequate Dietary Phosphorus Levels Cause Skeletal Anomalies and Alter Osteocalcin Gene Expression in Zebrafish

 ,
,
Abstract
:1. Introduction
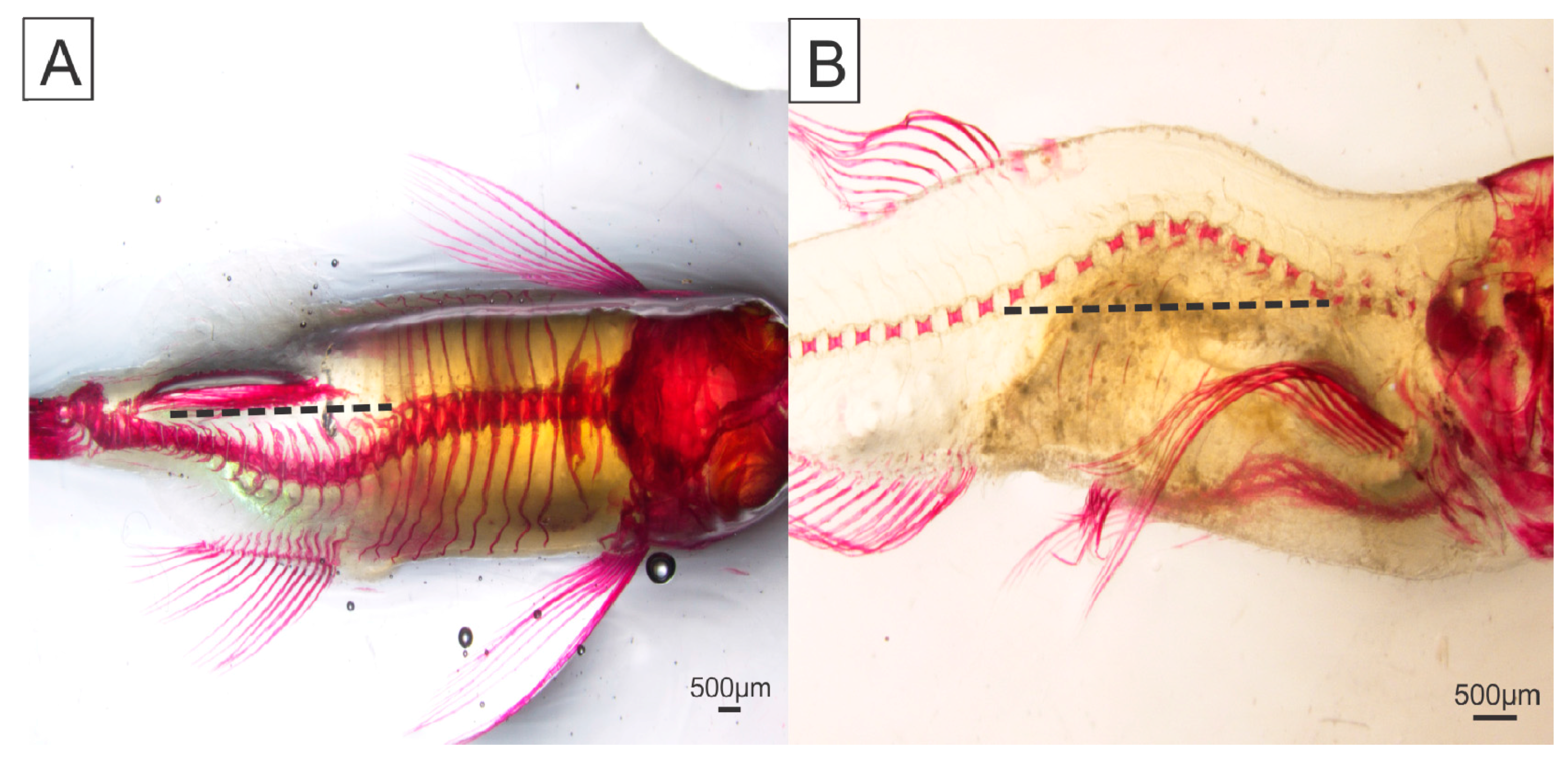
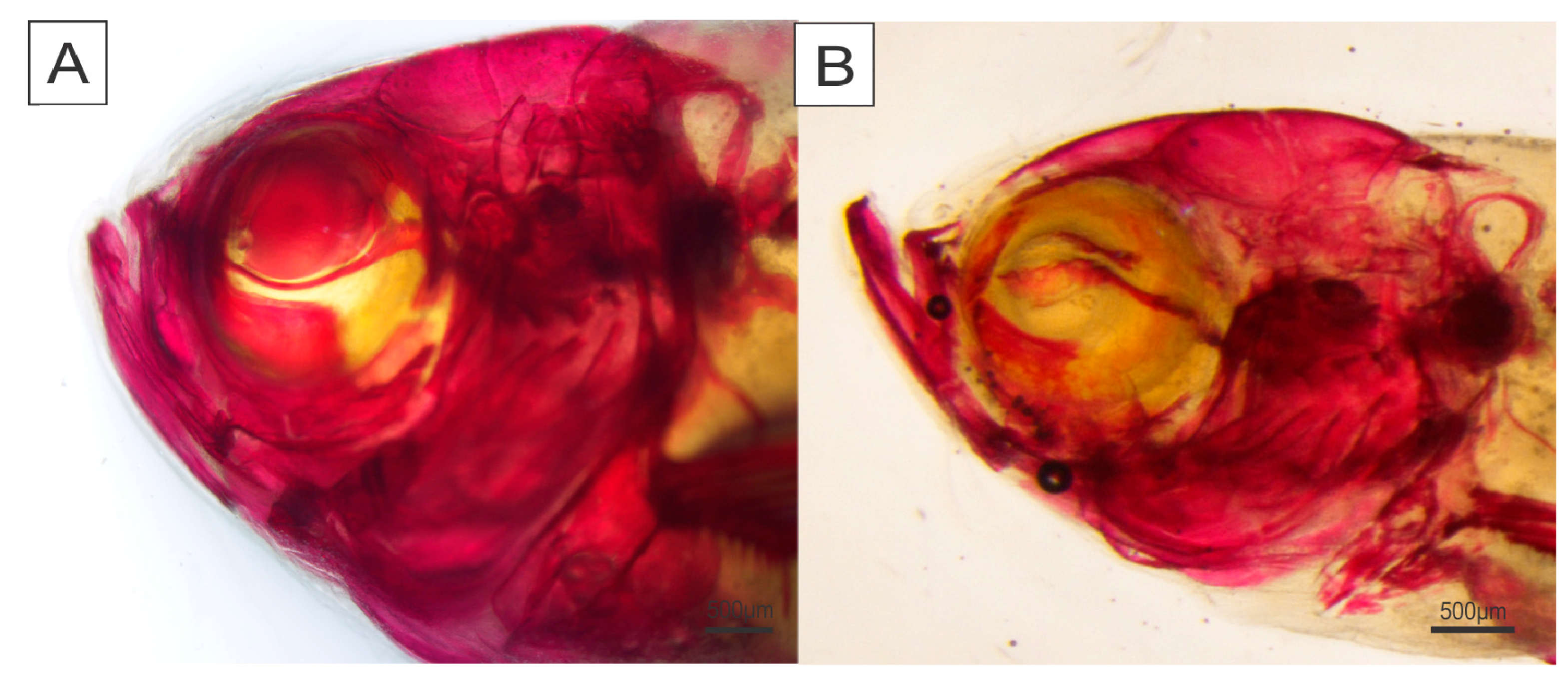
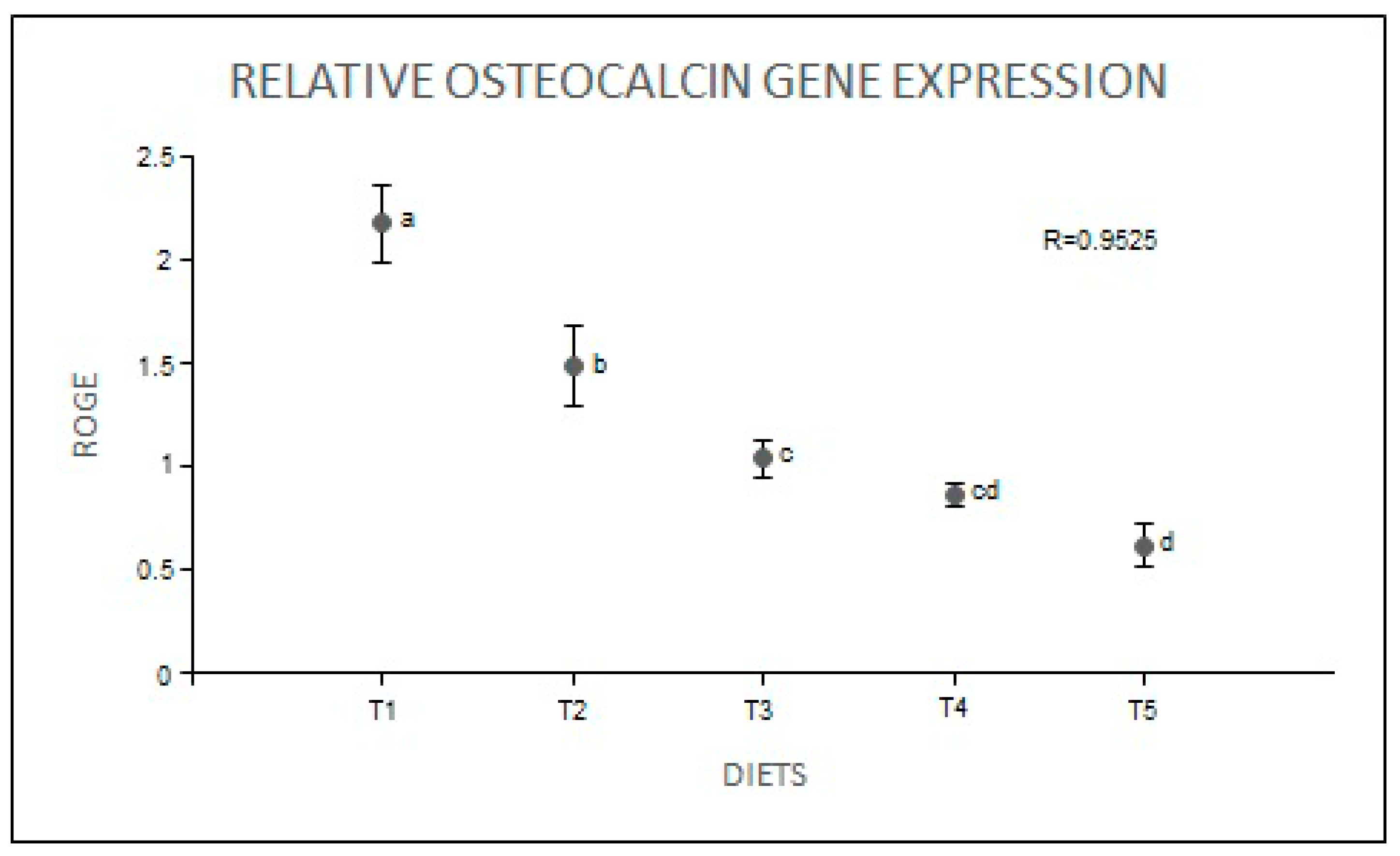
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Animal Production
4.2. Nutrition Experiment
4.3. Diets
4.4. Diaphanization
4.5. Skeletal Deformities
4.6. Gene Expression
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jobling, M. National Research Council (NRC): Nutrient requirements of fish and shrimp. Aquac. Int. 2011, 20, 601–602. [Google Scholar] [CrossRef]
- Sapir-Koren, R.; Livshits, G. Bone Mineralization and Regulation of Phosphate Homeostasis. IBMS BoneKEy 2011, 8, 286–300. [Google Scholar] [CrossRef]
- Grabowski, P. Physiology of bone. Endocr. Dev. 2009, 16, 32–48. [Google Scholar] [PubMed]
- Lall, S.P. The Minerals. In Fish Nutrition, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 259–308. [Google Scholar] [CrossRef]
- Leprévost, A.; Sire, J.Y. Architecture, mineralization and development of the axial skeleton in Acipenseriformes, and occurrences of axial anomalies in rearing conditions; can current knowledge in teleost fish help? J. Appl. Ichthyol. 2014, 30, 767–776. [Google Scholar] [CrossRef]
- Bird, N.C.; Mabee, P.M. Developmental morphology of the axial skeleton of the zebrafish, Danio rerio (Ostariophysi: Cyprinidae). Dev. Dyn. 2003, 228, 337–357. [Google Scholar] [CrossRef] [PubMed]
- Boglione, C.; Gisbert, E.; Gavaia, P.; Witten, P.E.; Moren, M.; Fontagné, S.; Koumoundouros, G. Skeletal anomalies in reared European fish larvae and juveniles. Part 2: Main typologies, occurrences and causative factors. Rev. Aquac. 2013, 5. [Google Scholar] [CrossRef]
- Fjelldal, P.G.; Hansen, T.J.; Lock, E.-J.; Wargelius, A.; Fraser, T.W.K.; Sambraus, F.; El-Mowafi, A.; Albrektsen, S.; Waagbø, R.; Ørnsrud, R. Increased dietary phosphorous prevents vertebral deformities in triploid Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2016, 22, 72–90. [Google Scholar] [CrossRef]
- Witten, P.E.; Owen, M.A.; Fontanillas, R.; Soenens, M.; McGurk, C.; Obach, A. A primary phosphorus-deficient skeletal phenotype in juvenile Atlantic salmon Salmo salar: The uncoupling of bone formation and mineralization. J. Fish Biol. 2016, 88, 690–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensimon-Brito, A.; Cardeira, J.; Dionísio, G.; Huysseune, A.; Cancela, M.L.; Witten, P.E. Revisiting in vivo staining with alizarin red S—A valuable approach to analyze zebrafish skeletal mineralization during development and regeneration. BMC Dev. Biol. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreri, F.; Nicolais, C.; Boglione, C.; Bertolini, B. Skeletal characterization of wild and reared zebrafish: Anomalies and meristic characters. J. Fish Biol. 2000, 56, 1115–1128. [Google Scholar] [CrossRef]
- Geurtzen, K.; Knopf, F.; Wehner, D.; Huitema, L.F.A.; Leonie, F.A.; Schulte-Merker, S.; Weidinger, G. Mature osteoblasts dedifferentiate in response to traumatic bone injury in the zebrafish fin and skull. Dev. 2014, 14, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Suttie, J.W.; John, W. Vitamin K-Dependent Proteins and Their Metabolic Roles; Elsevier: Amsterdam, The Netherlands, 1990; ISBN 0444015663. [Google Scholar]
- Harada, S.; Rodan, G. Control of osteoblast function and regulation of bone mass. Nature 2003, 423, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Olson, E. Bone and muscle endocrine functions: Unexpected paradigms of inter-organ communication. Cell 2016, 164, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Oury, F. Regulation of male fertility by the bone-derived hormone osteocalcin. Mol. Cell. Endocrinol. 2014, 382, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Neve, A.; Corrado, A.; Cantatore, F.P. Osteocalcin: Skeletal and extra-skeletal effects. J. Cell. Physiol. 2013, 228, 1149–1153. [Google Scholar] [CrossRef] [PubMed]
- Mera, P.; Laue, K.; Ferron, M.; Confavreux, C.; Wei, J.; Galán-Díez, M.; Lacampagne, A.; Mitchell, S.J.; Mattison, J.A.; Chen, Y.; et al. Osteocalcin signaling in myofibers is necessary and sufficient for optimum adaptation to exercise. Cell Metab. 2016, 23, 1078–1092. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, S.K.; Waite, J.H.; Nishimoto, M.; Kriwacki, R.W. Structure, activity, and distribution of fish osteocalcin. J. Biol. Chem. 2003, 278, 11843–11848. [Google Scholar] [CrossRef] [PubMed]
- Ponomareva, N.; Poprygina, T.; Lesovoi, M.; Sokolov, Y.V.; Agapov, B.L. Crystal structure and composition of hydroxyapatite biocomposites prepared at excess of calcium ions. Russ. J. Gen. Chem. 2009, 79, 186–190. [Google Scholar] [CrossRef]
- Dahm, R.; Geisler, R. Learning from small fry: The zebrafish as a genetic model organism for aquaculture fish species. Mar. Biotechnol. 2006, 8, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Wegrzyn, D.; Ortubay, S. Salmonidos en Patagonia; Mar del Plata: Bariloche, Argentina, 2009. [Google Scholar]
- Biga, P.R.; Goetz, F.W. Zebrafish and giant danio as models for muscle growth: Determinate vs. indeterminate growth as determined by morphometric analysis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1327–R1337. [Google Scholar] [CrossRef] [PubMed]
- Johnston, I.; Macqueen, D.; Watabe, S. Molecular Biotechnology of Development and Growth in Fish Muscle; Terrapub: Tokyo, Japan, 2008. [Google Scholar]
- Ulloa, P.E.; Iturra, P.; Neira, R.; Araneda, C. Zebrafish as a model organism for nutrition and growth: Towards comparative studies of nutritional genomics applied to aquacultured fishes. Rev. Fish Biol. Fish. 2011, 21, 649–666. [Google Scholar] [CrossRef]
- Bruneel, B.; Witten, P.E. Power and challenges of using zebrafish as a model for skeletal tissue imaging. Connect. Tissue Res. 2015, 56, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.A.; Powell, M.; D’Abramo, L.R. Fundamental approaches to the study of zebrafish nutrition. ILAR J. 2012, 53, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.A.; Lawrence, C.; Powell, M.; D’Abramo, L.R. The Vital Relationship between Nutrition and Health in Zebrafish. Zebrafish 2016, 13, S72–S76. [Google Scholar] [CrossRef] [PubMed]
- Borlongan, I.G.; Satoh, S. Dietary phosphorus requirement of juvenile milkfish, Chanos chanos (Forsskal). Aquac. Res. 2001, 32, 26–32. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Pimentel-Rodrigues, A. Phosphorus requirement of European sea bass (Dicentrarchus labrax L.) juveniles. Aquac. Res. 2004, 35, 636–642. [Google Scholar] [CrossRef]
- Kamler, E.; Wolnicki, J.; Kamiński, R.; Sikorska, J. Fatty acid composition, growth and morphological deformities in juvenile cyprinid, Scardinius erythrophthalmus fed formulated diet supplemented with natural food. Aquaculture 2008, 278, 69–76. [Google Scholar] [CrossRef]
- Tocher, D.R.; Bendiksen, E.Â.; Campbell, P.J.; Bell, J.G. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef]
- Kaushik, S.J. Mineral nutrition. In Nutrition and Feeding of Fish and Crustaceans; Springer: London, UK, 2001; pp. 169–181. ISBN 1852332417. [Google Scholar]
- Yao, W.; Lv, Y.; Gong, X.; Wu, J.; Bao, B. Different ossification patterns of intermuscular bones in fish with different swimming modes. Biol. Open 2015, 4, 1727–1732. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.; Reid, S.W.; Ternent, H.; Manchester, N.J.; Roberts, R.J.; Stone, D.A.; Hardy, R.W. The aetiology of spinal deformity in Atlantic salmon, Salmo salar L.: Influence of different commercial diets on the incidence and severity of the preclinical condition in salmon parr under two contrasting husbandry regimes. J. Fish Dis. 2007, 30, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.; Hammond, G.; Roberts, R.J.; Manchester, N.J. Spinal deformation in commercially cultured Atlantic salmon, Salmo salar L.: A clinical and radiological study. J. Fish Dis. 2007, 30, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.K.; Lall, S.P. Dietary phosphorus requirement of juvenile haddock (Melanogrammus aeglefinus L.). Aquaculture 2003, 221, 451–468. [Google Scholar] [CrossRef]
- Yang, S.; Lin, T.S.; Liu, F.; Liou, C. Influence of dietary phosphorus levels on growth, metabolic response and body composition of juvenile silver perch (Bidyanus bidyanus). Aquaculture 2006, 253, 592–601. [Google Scholar] [CrossRef]
- Pittman, K.; Skiftesvik, A.B.; Harboe, T. Effect of temperature on growth rates and organogenesis in the larvae of halibut (Hippoglossus hippoglossus L.). Rapp. P.-v. Reun. Cons. Int. Explor. Mer 1989, 191, 423–430. [Google Scholar]
- Panserat, S.; Kaushik, S.J. Regulation of gene expression by nutritional factors in fish. Aquac. Res. 2010, 41, 751–762. [Google Scholar] [CrossRef]
- Shih, Y.V.; Hwang, Y.; Phadke, A.; Kang, H.; Hwang, N.S.; Caro, E.J.; Nguyen, S.; Siu, M.; Theodorakis, E.A.; Gianneschi, N.C.; et al. Calcium phosphate-bearing matrices induce osteogenic differentiation of stem cells through adenosine signaling. Proc. Natl. Acad. Sci. USA 2013, 111, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Gundberg, C.M.; Lian, J.B.; Booth, S.L. Vitamin K-Dependent Carboxylation of Osteocalcin: Friend or Foe? Adv. Nutr. 2012, 3, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Young, M.F. Bone matrix proteins: Their function, regulation, and relationship to osteoporosis. Osteoporos. Int. 2003, 14, S35–S42. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. A Guide for the Laboratory Use of Zebrafish (Danio Rerio); University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Do Carmo E Sá, M.V.; Pezzato, L.E.; Barros, M.M.; De Magalhes Padilha, P. Relative bioavailability of zinc in supplemental inorganic and organic sources for Nile tilapia Oreochromis niloticus fingerlings. Aquac. Nutr. 2005, 11, 273–281. [Google Scholar] [CrossRef]
- Markzent, Z. Spectrophotometric Determination of Elements; Ellis Howood: Chichester, UK, 2002; ISBN 0003-2670. [Google Scholar]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists International, 20th ed.; AOAC International: Washington, DC, USA, 2016; ISBN 0-935584-42-0. [Google Scholar]
- Potthoff, T. Clearing and Staining Techniques. In Ontogeny and Systematics of Fishes; American Society of Ichthyologists and Herpetologists: Lawrence, KS, USA, 1984; pp. 35–37. ISBN 0031208000233. [Google Scholar]
- Tang, R.; Dodd, A.; Lai, D.; McNabb, W.C.; Love, D.R. Validation of zebrafish (Danio rerio) reference genes for quantitative real-time RT-PCR normalization. Acta Biochim. Biophys. Sin. 2007, 39, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 16–21. [Google Scholar] [CrossRef]
- Ayres, M.; Ayres, M.; Ayres, D.; Santos, A. BIOESTAT 2.0–Aplicações Estatísticas nas Áreas das Ciências Biológicas e Médicas; Sociedade Civil Mamirauá: Brasília, Brazil, 2003. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anomalies * | Diets | |||||
|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T5 | T6 | |
| Absence of parallelism between neural and hemal spines | 51.5 a | 52.0 a | 43.0 a | 39.0 ab | 5.5 bc | 0.0 c |
| Partial neural spine fusion | 10.0 a | 9.0 a | 6.0 a | 10.0 a | 3.0 c | 0.0 c |
| Partial hemal spine fusion | 10.0 a | 10.0 a | 8.5 a | 9.5 a | 3.0 c | 0.0 c |
| Absence of vertebral body fusion fusion | 35.0 ab | 70.0 a | 61.5 a | 16.0 b | 6.0 bc | 0.0 c |
| Normal intervertebral spacing | 20.0 c | 56.5 b | 54.0 b | 100.0 a | 100.0 a | 100.0 a |
| Vertebral compression | 10.0 bc | 32.5 a | 21.0 a | 14.0 b | 6.0 bc | 0.0 c |
| Ingredients (%) | Diets | |||||
|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T5 | T6 | |
| Albumin | 34.74 | 34.76 | 34.79 | 34.81 | 34.83 | 34.85 |
| Poultry Viscera Meal | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Gelatin | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 |
| Starch | 38.95 | 36.01 | 33.11 | 30.21 | 27.31 | 24.42 |
| Soy oil | 2.64 | 3.51 | 4.36 | 5.21 | 6.07 | 6.92 |
| Cellulose | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Dicalcium phosphate a | 0.00 | 2.05 | 4.07 | 6.10 | 8.12 | 10.14 |
| BHT b | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| NaCl | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Premix c | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Vitamin C d | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Chemical Composition * | ||||||
| Crude Protein % | 40.13 | 40.07 | 40.00 | 40.08 | 40.01 | 40.10 |
| Crude Lipid % | 4.53 | 5.39 | 6.24 | 7.09 | 7.93 | 8.78 |
| Total Phosphorus (%) | 0.34 | 0.67 | 0.93 | 1.26 | 1.55 | 1.81 |
| Ca/P ratio (%) | 2.03 | 2.25 | 2.32 | 2.34 | 2.37 | 2.38 |
| Energy (kcal−1) | 3.493 | 3.494 | 3.506 | 3.506 | 3.508 | 3.493 |
| Deformities | Scale | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| Number of Vertebrae (V) or Spines (S) Affected | ||||||
| Scoliosis (V) | 1–2 | 3–4 | 5–6 | 7–8 | 9–10 | ≥11 |
| Lordosis (V) | 1–2 | 3–4 | 5–6 | 7–8 | 9–10 | ≥11 |
| Anchylosis (S) | 1–2 | 3–4 | 5–6 | 7–8 | 9–10 | ≥11 |
| Caudal fin anomaly (V) | 1 | 2 | 3 | 4 | 5 | ≥6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, J.M.; Sartori, M.M.P.; Nascimento, N.F.d.; Kadri, S.M.; Ribolla, P.E.M.; Pinhal, D.; Pezzato, L.E. Inadequate Dietary Phosphorus Levels Cause Skeletal Anomalies and Alter Osteocalcin Gene Expression in Zebrafish. Int. J. Mol. Sci. 2018, 19, 364. https://doi.org/10.3390/ijms19020364
Costa JM, Sartori MMP, Nascimento NFd, Kadri SM, Ribolla PEM, Pinhal D, Pezzato LE. Inadequate Dietary Phosphorus Levels Cause Skeletal Anomalies and Alter Osteocalcin Gene Expression in Zebrafish. International Journal of Molecular Sciences. 2018; 19(2):364. https://doi.org/10.3390/ijms19020364
Chicago/Turabian StyleCosta, Juliana M., Maria M. P. Sartori, Nivaldo F. do Nascimento, Samir M. Kadri, Paulo E. M. Ribolla, Danillo Pinhal, and Luiz E. Pezzato. 2018. "Inadequate Dietary Phosphorus Levels Cause Skeletal Anomalies and Alter Osteocalcin Gene Expression in Zebrafish" International Journal of Molecular Sciences 19, no. 2: 364. https://doi.org/10.3390/ijms19020364
APA StyleCosta, J. M., Sartori, M. M. P., Nascimento, N. F. d., Kadri, S. M., Ribolla, P. E. M., Pinhal, D., & Pezzato, L. E. (2018). Inadequate Dietary Phosphorus Levels Cause Skeletal Anomalies and Alter Osteocalcin Gene Expression in Zebrafish. International Journal of Molecular Sciences, 19(2), 364. https://doi.org/10.3390/ijms19020364