Exploring the History of Chloroplast Capture in Arabis Using Whole Chloroplast Genome Sequencing



Abstract
:1. Introduction
2. Results
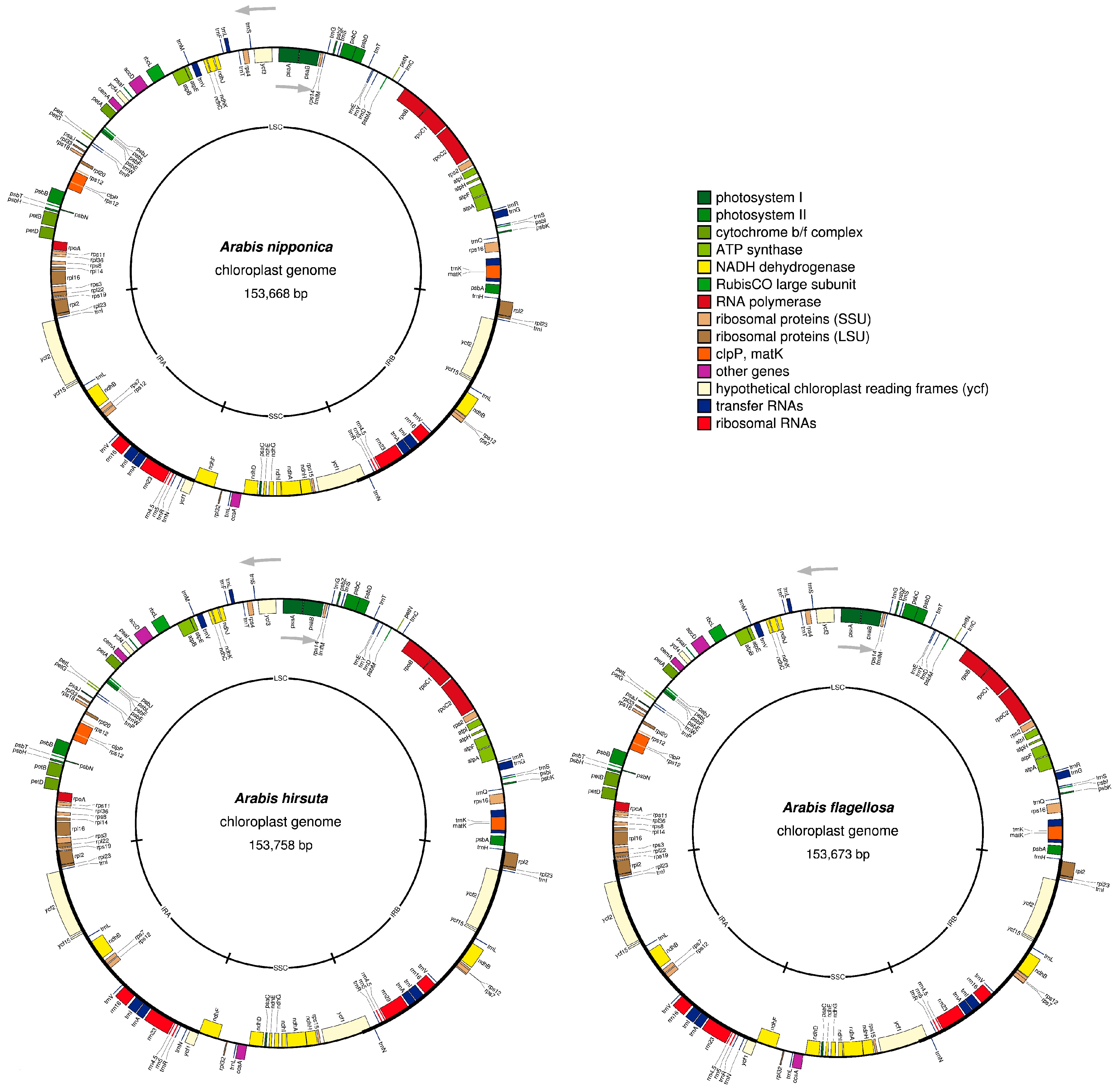
2.1. Chloroplast Genome Structure of Arabis Species
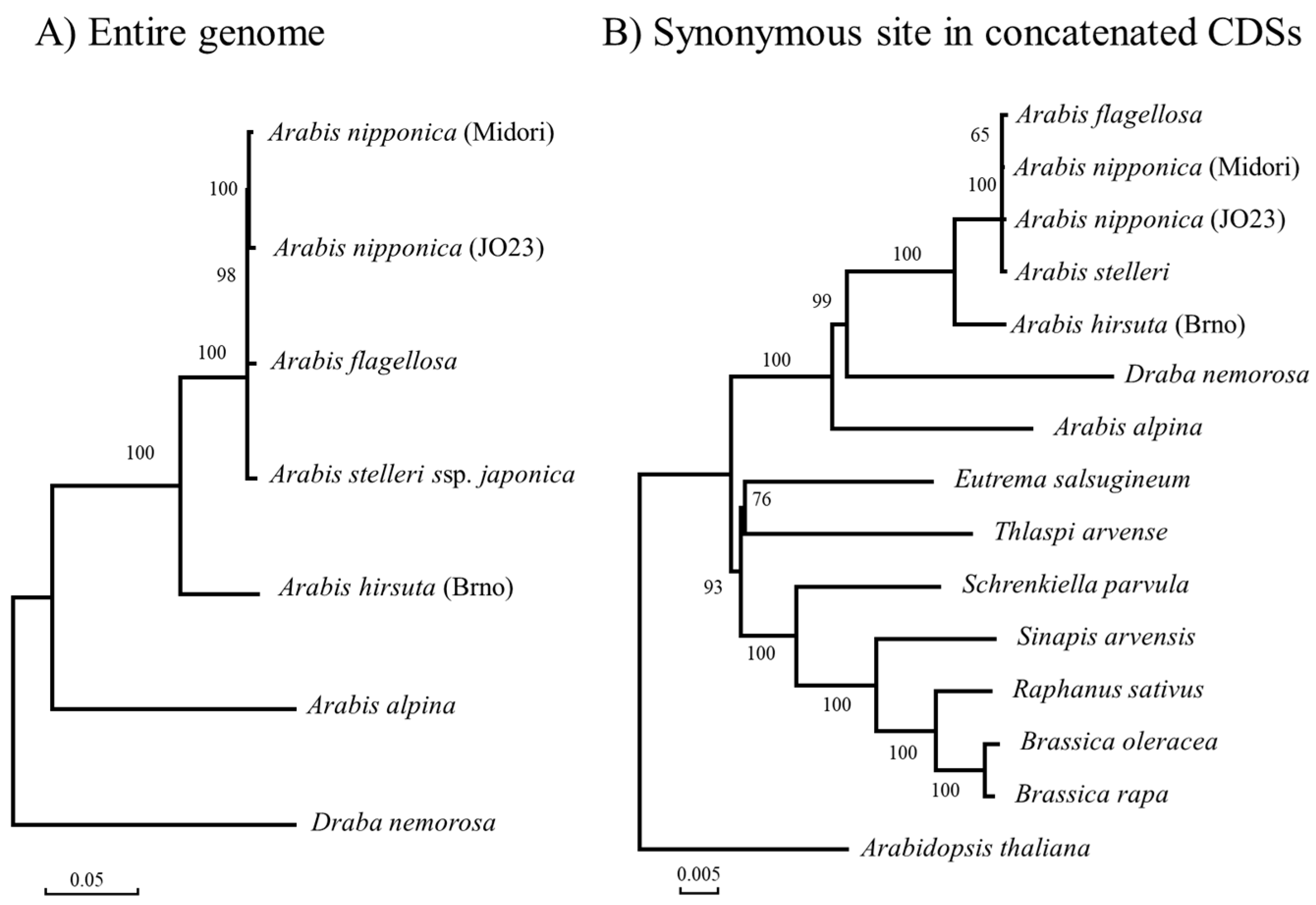
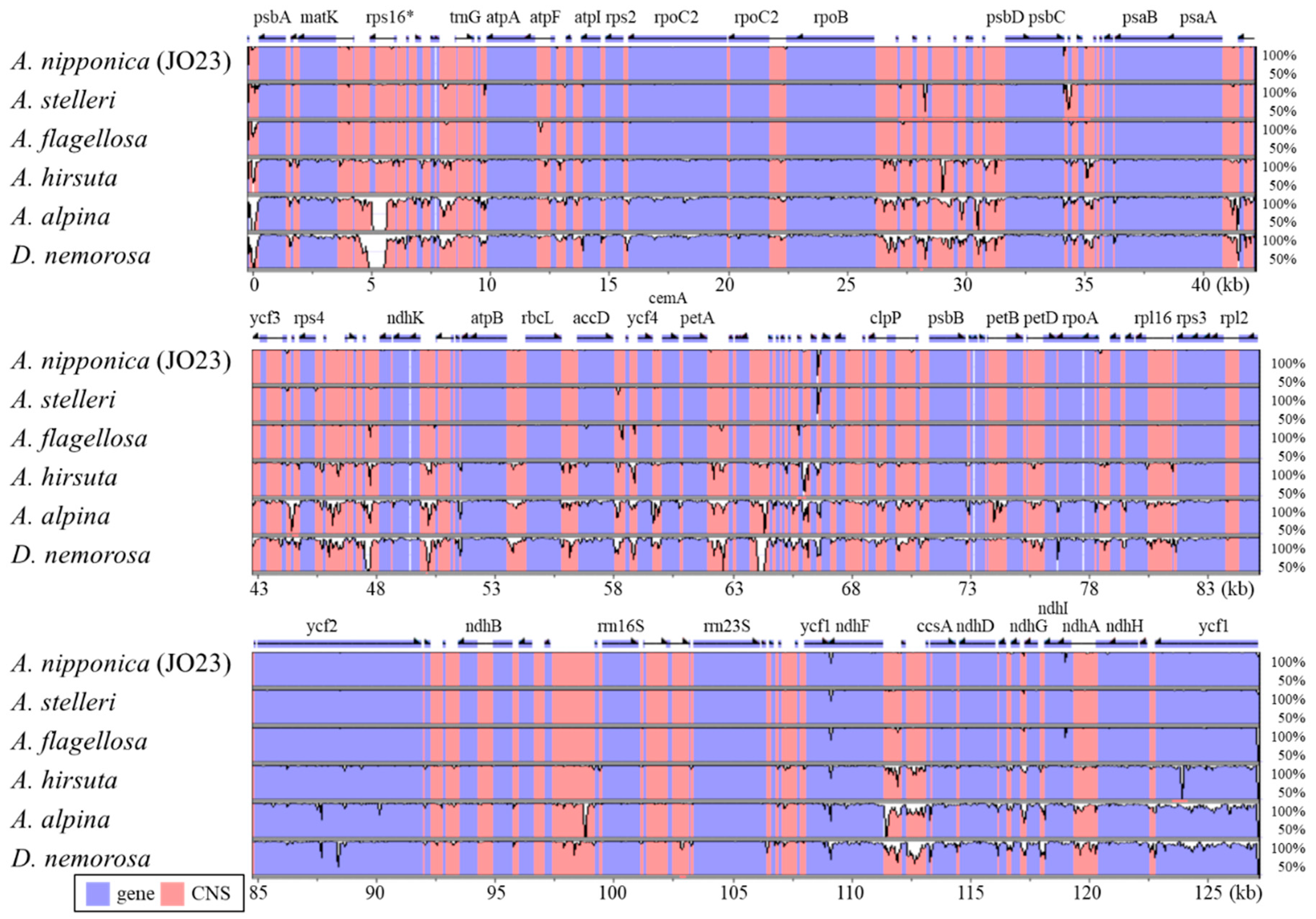
2.2. Chloroplast Genome Divergence
2.3. Distribution of Simple Sequence Repeats in the Chloroplast Genomes
2.4. Mitochondrial Genome Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. DNA Isolation, NGS Sequencing, and Genome Assembly
4.3. Molecular Evolutionary Analyses
4.4. Mapping NGS Reads to Mitochondrial Genome Sequences
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Al-Shehbaz, I.A. Transfer of most North American species of Arabis to Boechera (Brassicaceae). Novon 2003, 13, 381–391. [Google Scholar] [CrossRef]
- O’Kane, S.L.; Al-Shehbaz, I.A. Phylogenetic Position and Generic Limits of Arabidopsis (Brassicaceae) Based on Sequences of Nuclear Ribosomal DNA. Ann. Mo. Bot. Gard. 2003, 90, 603–612. [Google Scholar] [CrossRef]
- Al-Shehbaz, I.A.; Beilstein, M.A.; Kellogg, E.A. Systematics and phylogeny of the Brassicaceae (Cruciferae): An overview. Plant Syst. Evol. 2006, 259, 89–120. [Google Scholar] [CrossRef]
- Al-Shehbaz, I.A.; German, D.A.; Karl, R.; Ingrid, J.T.; Koch, M.A. Nomenclatural adjustments in the tribe Arabideae (Brassicaceae). Plant Div. Evol. 2011, 129, 71–76. [Google Scholar] [CrossRef]
- Koch, M.A.; Karl, R.; German, D.A.; Al-Shehbaz, I.A. Systematics, taxonomy and biogeography of three new Asian genera of Brassicaceae tribe Arabideae: An ancient distribution circle around the Asian high mountains. Taxon 2012, 61, 955–969. [Google Scholar]
- Kiefer, M.; Schmickl, R.; German, D.A.; Mandáková, T.; Lysak, M.A.; Al-Shehbaz, I.A.; Franzke, A.; Mummenhoff, K.; Stamatakis, A.; Koch, M.A. BrassiBase: Introduction to a novel knowledge database on Brassicaceae evolution. Plant Cell Physiol. 2014, 55, e3. [Google Scholar] [CrossRef] [PubMed]
- Ansell, S.W.; Grundmann, M.; Russell, S.J.; Schneider, H.; Vogel, J.C. Genetic discontinuity, breeding-system change and population history of Arabis alpina in the Italian Peninsula and adjacent Alps. Mol Ecol. 2008, 17, 2245–2257. [Google Scholar] [CrossRef] [PubMed]
- Bergonzi, S.; Albani, M.C.; ver Loren van Themaat, E.; Nordström, K.J.; Wang, R.; Schneeberger, K.; Moerland, P.D.; Coupland, G. Mechanisms of age-dependent response to winter temperature in perennial flowering of Arabis alpina. Science 2013, 340, 1094–1097. [Google Scholar] [CrossRef] [PubMed]
- Karl, R.; Koch, M.A. A world-wide perspective on crucifer speciation and evolution: Phylogenetics, biogeography and trait evolution in tribe Arabideae. Ann. Bot. 2013, 112, 983–1001. [Google Scholar] [CrossRef] [PubMed]
- Toräng, P.; Vikström, L.; Wunder, J.; Wötzel, S.; Coupland, G.; Ågren, J. Evolution of the selfing syndrome: Anther orientation and herkogamy together determine reproductive assurance in a self-compatible plant. Evolution 2017, 71, 2206–2218. [Google Scholar] [CrossRef] [PubMed]
- Heidel, A.J.; Kiefer, C.; Coupland, G.; Rose, L.E. Pinpointing genes underlying annual/perennial transitions with comparative genomics. BMC Genom. 2016, 17, 921. [Google Scholar] [CrossRef] [PubMed]
- Willing, E.M.; Rawat, V.; Mandáková, T.; Maumus, F.; James, G.V.; Nordström, K.J.; Becker, C.; Warthmann, N.; Chica, C.; Szarzynska, B.; et al. Genome expansion of Arabis alpina linked with retrotransposition and reduced symmetric DNA methylation. Nat. Plants 2015, 1, 14023. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.B.; Accinelli, G.G.; Hartwig, B.; Kiefer, C.; Baker, D.; Severing, E.; Willing, E.M.; Piednoel, M.; Woetzel, S.; Madrid-Herrero, E.; et al. Improving and correcting the contiguity of long-read genome assemblies of three plant species using optical mapping and chromosome conformation capture data. Genome Res. 2017, 27, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.A.; Karl, R.; Kiefer, C.; Al-Shehbaz, I.A. Colonizing the American continent: Systematics of the genus Arabis in North America (Brassicaceae). Am. J. Bot. 2010, 97, 1040–1057. [Google Scholar] [CrossRef] [PubMed]
- Karl, R.; Kiefer, C.; Ansell, S.W.; Koch, M.A. Systematics and evolution of Arctic-Alpine Arabis alpina (Brassicaceae) and its closest relatives in the eastern Mediterranean. Am. J. Bot. 2012, 99, 778–794. [Google Scholar] [CrossRef] [PubMed]
- Karl, R.; Koch, M.A. Phylogenetic signatures of adaptation: The Arabis hirsuta species aggregate (Brassicaceae) revisited. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 247–264. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Soltis, D.E. Phylogenetic consequences of cytoplasmic gene flow in plants. Evoluti. Trends Plants 1991, 5, 65–84. [Google Scholar]
- Soltis, D.E.; Kuzoff, R.K. Discordance between nuclear and chloroplast phylogenies in the Heuchera group (Saxifragaceae). Evolution 1995, 49, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Ruhfel, B.R.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E.; Burleigh, J.G. From algae to angiosperms-inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes. BMC Evol. Biol. 2014, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, N.; Wolf, E.M.; Lysak, M.A.; Koch, M.A. A Time-calibrated road map of brassicaceae species radiation and evolutionary history. Plant Cell 2015, 27, 2770–2784. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, J.; Hao, G.; Zhang, L.; Mao, K.; Wang, X.; Zhang, D.; Ma, T.; Hu, Q.; Al-Shehbaz, I.A.; Koch, M.A. Plastome phylogeny and early diversification of Brassicaceae. BMC Genom. 2017, 18, 176. [Google Scholar] [CrossRef] [PubMed]
- Mandáková, T.; Hloušková, P.; German, D.A.; Lysak, M.A. Monophyletic origin and evolution of the largest crucifer genomes. Plant Physiol. 2017, 174, 2062–2071. [Google Scholar] [CrossRef] [PubMed]
- Raman, G.; Park, V.; Kwak, M.; Lee, B.; Park, S. Characterization of the complete chloroplast genome of Arabis stellari and comparisons with related species. PLoS ONE 2017, 12, e0183197. [Google Scholar] [CrossRef] [PubMed]
- Jordon-Thaden, I.; Hase, I.; Al-Shehbaz, I.A.; Koch, M.A. Molecular phylogeny and systematics of the genus Draba (Brassicaceae) and identification of its most closely related genera. Mol. Phylogenet. Evol. 2010, 55, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Melodelima, C.; Lobréaux, S. Complete Arabis alpina chloroplast genome sequence and insight into its polymorphism. Meta Gene 2013, 1, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Acosta, M.C.; Premoli, A.C. Evidence of chloroplast capture in South American Nothofagus (subgenus Nothofagus, Nothofagaceae). Mol. Phylogenet. Evol. 2010, 54, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Dorado, O.; Rieseberg, L.H.; Arias, D.M. Chloroplast DNA introgression in southern California sunflowers. Evolution 1992, 46, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Fehrer, J.; Gemeinholzer, B.; Chrtek, J.; Bräutigam, S. Incongruent plastid and nuclear DNA phylogenies reveal ancient intergeneric hybridization in Pilosella hawkweeds (Hieracium, Cichorieae, Asteraceae). Mol. Phylogenet. Evol. 2007, 42, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Gurushidze, M.; Fritsch, R.M.; Blattner, F.R. Species-level phylogeny of Allium subgenus Melanocrommyum: Incomplete lineage sorting, hybridization and trnF gene duplication. Taxon 2010, 59, 829–840. [Google Scholar]
- Liston, A.; Kadereit, J.W. Chloroplast DNA evidence for introgression and long distance dispersal in the desert annual Senecio flavus (Asteraceae). Plant Syst. Evol. 1995, 197, 33–41. [Google Scholar] [CrossRef]
- Mir, C.; Jarne, P.; Sarda, V.; Bonin, A.; Lumaret, R. Contrasting nuclear and cytoplasmic exchanges between phylogenetically distant oak species (Quercus suber L. and Q. ilex L.) in Southern France: Inferring crosses and dynamics. Plant Biol. 2009, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, Y.; Fujii, N.; Wakabayashi, M.; Kawakita, A.; Ito, M.; Watanabe, M.; Murakami, N.; Kato, M. Nonuniform concerted evolution and chloroplast capture: Heterogeneity of observed introgression patterns in three molecular data partition phylogenies of Asian Mitella (Saxifragaceae). Mol. Biol. Evol. 2005, 22, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H.; Choi, H.C.; Ham, F. Differential cytoplasmic versus nuclear introgression in Helianthus. J. Hered. 1991, 82, 489–493. [Google Scholar] [CrossRef]
- Schilling, E.E.; Panero, J.K. Phylogenetic reticulation in subtribe Helianthinae. Am. J. Bot. 1996, 83, 939–948. [Google Scholar] [CrossRef]
- Wolfe, A.D.; Elisens, W.J. Evidence of chloroplast capture and pollen-mediated gene flow in Penstemon sect. Peltanthera (Scrophulariaceae). Syst. Bot. 1995, 20, 395–412. [Google Scholar] [CrossRef]
- Yi, T.S.; Jin, G.H.; Wen, J. Chloroplast capture and intra-and inter-continental biogeographic diversification in the Asian–New World disjunct plant genus Osmorhiza (Apiaceae). Mol. Phylogenet. Evol. 2015, 85, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.W.; Olmstead, R.G. A species-level phylogenetic study of the Verbena complex (Verbenaceae) indicates two independent intergeneric chloroplast transfers. Mol. Phylogenet. Evol. 2008, 48, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C. Molecular Markers, Natural History and Evolution, 2nd ed.; Sinauer: Sunderland, MA, USA, 2004. [Google Scholar]
- Rieseberg, L.H.; Wendel, J. Introgression and its consequences in plants. In Hybrid Zones and the Evolutionary Process; Harrison, R., Ed.; Oxford University Press: New York, NY, USA, 1993; pp. 70–109. [Google Scholar]
- Martinsen, G.D.; Whitham, T.G.; Turek, R.J.; Keim, P. Hybrid populations selectively filter gene introgression between species. Evolution 2001, 55, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H. The role of hybridization in evolution: Old wine in new skins. Am. J. Bot. 1995, 82, 944–953. [Google Scholar] [CrossRef]
- Tsitrone, A.; Kirkpatrick, M.; Levin, D.A. A model for chloroplast capture. Evolution 2003, 57, 1776–1782. [Google Scholar] [CrossRef] [PubMed]
- Okegawa, Y.; Motohashi, K. Chloroplastic thioredoxin m functions as a major regulator of Calvin cycle enzymes during photosynthesis in vivo. Plant J. 2015, 84, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW)—A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Seol, Y.J.; Kim, K.; Kang, S.H.; Perumal, S.; Lee, J.; Kim, C.K. The complete chloroplast genome of two Brassica species, Brassica nigra and B. oleracea. Mitochondrial DNA Part A 2017, 28, 167–168. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Hao, G.; Wang, X.; Bi, H.; Li, Y.; Guo, X.; Ma, T. The complete chloroplast genome of Schrenkiella parvula (Brassicaceae). Mitochondrial DNA Part A 2016, 27, 3527–3528. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.M.; Chung, W.H.; Mun, J.H.; Kim, N.; Yu, H.J. De novo assembly and characterization of the complete chloroplast genome of radish (Raphanus sativus L.). Gene 2014, 551, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Nakamura, Y.; Kaneko, T.; Asamizu, E.; Tabata, S. Complete structure of the chloroplast genome of Arabidopsis thaliana. DNA Res. 1999, 6, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal Alignment: Finding rearrangements during alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chu, P.; Yang, Q.; Chang, S.; Chen, J.; Hu, M.; Guan, R. Complete mitochondrial genome of Eruca sativa Mill. (Garden rocket). PLoS ONE 2014, 9, e105748. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Nucleotide Length (bp) | GC Contents (%) | NCBI # | Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Entire | LSC | SSC | IR | Entire | LSC | SSC | IR | ||||
| Draba nemorosa | JO21 | 153289 | 82457 | 18126 | 26353 | 36.47 | 34.27 | 29.3 | 42.39 | AP009373 (NC009272) | |
| Arabis alpina | 152866 | 82338 | 17938 | 26933 | 36.45 | 34.21 | 29.31 | 42.39 | HF934132 (NC023367) | [25] | |
| Arabis hirsuta | Brno | 153758 | 82710 | 18156 | 26446 | 36.4 | 34.15 | 29.16 | 42.41 | LC361350 | this study |
| Arabis flagellosa | Kifune | 153673 | 82775 | 18052 | 26423 | 36.4 | 34.13 | 29.22 | 42.41 | LC361351 | this study |
| Arabis stelleri | 153683 | 82807 | 18030 | 26423 | 36.39 | 34.11 | 29.22 | 42.42 | KY126841 | [23] | |
| Arabis nipponica | JO23 | 153689 | 82811 | 18036 | 26421 | 36.4 | 34.1 | 29.31 | 42.42 | AP009369 (NC009268) | |
| Arabis nipponica | Midori | 153668 | 82772 | 18052 | 26422 | 36.39 | 34.1 | 29.24 | 42.42 | LC361349 | this study |
| Compared Species | # of Differences | Divergence (%: Ks with JC Correction) | ||
|---|---|---|---|---|
| Draba nemorosa | vs. | Arabis alpina | 4475 | 2.976 |
| Draba nemorosa | vs. | Arabis hirsuta | 4219 | 2.801 |
| Draba nemorosa | vs. | Arabis flagellosa | 4262 | 2.765 |
| Draba nemorosa | vs. | Arabis stelleri | 4171 | 2.771 |
| Draba nemorosa | vs. | Arabis nipponica (JO23) | 4150 | 2.757 |
| Draba nemorosa | vs. | Arabis nipponica (Midori) | 4131 | 2.745 |
| Arabis alpina | vs. | Arabis hirsuta | 3566 | 2.366 |
| Arabis alpina | vs. | Arabis flagellosa | 3571 | 2.371 |
| Arabis alpina | vs. | Arabis stelleri | 3565 | 2.366 |
| Arabis alpina | vs. | Arabis nipponica (JO23) | 3564 | 2.366 |
| Arabis alpina | vs. | Arabis nipponica (Midori) | 3547 | 2.355 |
| Arabis hirsuta | vs. | Arabis flagellosa | 1245 | 0.815 |
| Arabis hirsuta | vs. | Arabis stelleri | 1253 | 0.82 |
| Arabis hirsuta | vs. | Arabis nipponica (JO23) | 1234 | 0.808 |
| Arabis hirsuta | vs. | Arabis nipponica (Midori) | 1214 | 0.795 |
| Arabis flagellosa | vs. | Arabis stelleri | 132 | 0.086 |
| Arabis flagellosa | vs. | Arabis nipponica (JO23) | 111 | 0.072 |
| Arabis flagellosa | vs. | Arabis nipponica (Midori) | 86 | 0.056 |
| Arabis stelleri | vs. | Arabis nipponica (JO23) | 130 | 0.085 |
| Arabis stelleri | vs. | Arabis nipponica (Midori) | 104 | 0.068 |
| Arabis nipponica (JO23) | vs. | Arabis nipponica (Midori) | 55 | 0.036 |
| Position in A. nipponica (Midori) Genome | UNIT | A. nipponica | A. stelleri | A. flagellosa | A. hirsuta | A. alpina | |||
|---|---|---|---|---|---|---|---|---|---|
| from | to | Midori | JO23 | ||||||
| 287 | 318 | AT | 16 | 15 | 15 | 12 | 13 with 2 mutations | 29 bp with several mutations | |
| 1922 | 1932 | A | 11 | 11 | 9 | 12 | 11 | 9 | |
| 3929 | 3938 | T | 10 | 9 | 10 | 9 | 7 | 7 | |
| 4258 | 4270 | T | 13 | 18 | 18 | 17 | 13 | 13 | |
| 7713 | 7727 | T | 15 | 15 | 15 | 15 | 12 | 11 | |
| 7729 | 7738 | A | 10 | 10 | 10 | 9 | 10 | 10 | |
| 8203 | 8216 | TA | 7 | 6 | 7 | 7 | 6 | 6 | |
| 8273 | 8282 | TA | 5 | 5 | 5 | 5 | 5 | 6 | |
| 8289 | 8302 | AT | 7 | 7 | 6 | 7 | 8 | 6 | |
| 8321 | 8330 | TA | 5 | 5 | 4 | 5 | 5 | deletion | |
| 9677 | 9690 | T | 14 | 14 | T4GT10 | 15 | 14 | 14 | |
| 9982 | 9991 | TA | 5 | 5 | 5 | 5 | 5 | 5 | |
| 11,660 | 11,669 | A | 10 | 10 | 9 | 10 | 10 | 7 | |
| 12,406 | 12,414 | T | 9 | 9 | 10 | 10 | T3AT6 | T3AT6 | |
| 13,010 | 13,018 | T | 9 | 9 | 10 | 9 | T7AT2 | T10AT2 | |
| 13,810 | 13,821 | ATT | 4 | 4 | 4 | 4 | 4 | ATTATATTCTT | |
| 14,101 | 14,110 | A | 10 | 10 | 14 | 10 | 12 | 10 | |
| 18,027 | 18,037 | T | CDS | 11 | 11 | 11 | 11 | 11 | 11 |
| 19,398 | 19,408 | TA | 5 | 5 | 5 | 5 | 5 | 5 | |
| 22,549 | 22,558 | T | 10 | 10 | 11 | 10 | 9 | 15 | |
| 25,777 | 25,786 | T | CDS | 10 | 10 | 10 | 10 | 10 | 10 |
| 27,601 | 27,611 | G | 11 | 11 | 11 | 15 | 12 | 9 | |
| 28,808 | 28,817 | T | 10 | 10 | 9 | 10 | 10 | T5CT3G2 | |
| 30,293 | 30,310 | A | 18 | 17 | 17 | 18 | 12 | A4CA5 | |
| 30,737 | 30,751 | T | 15 | 15 | 14 | 15 | 15 | 5 | |
| 30,830 | 30,839 | A | 10 | 9 | 11 | 10 | 8 | 6 | |
| 30,918 | 30,929 | TA | 6 | 6 | 6 | 6 | 6 | 4 | |
| 31,260 | 31,269 | AT | 5 | 5 | 5 | 5 | 5 | 3 | |
| 35,309 | 35,316 | G | 8 | 11 | 10 | 11 | 7 | 10 | |
| 35,516 | 35,525 | AT | 5 | 5 | 5 | 5 | 5 | 5 | |
| 35,538 | 35,555 | AT | 9 | 9 | 9 | 9 | 3 | deletion | |
| 41,508 | 41,522 | T | 15 | 13 | 11 | 13 | 18nt | 101nt | |
| 41,768 | 41,778 | A | 11 | 12 | 12 | 11 | A12GA4 | 11 | |
| 43,656 | 43,665 | A | 10 | 10 | 10 | 11 | A8TA2 | A9TA2TA2 | |
| 43,887 | 43,895 | T | 9 | 15 | T4AT4AT4 | T4AT4 | 4 | 4 | |
| 45,038 | 45,046 | T | 9 | 9 | 9 | 10 | 8 | 7 | |
| 45,771 | 45,788 | T | 18 | 18 | 18 | 18 | 13 | 9 | |
| 46,034 | 46,057 | A | 24 | 24 | 24 | 24 | 16 | 11 | |
| 46,116 | 46,133 | AT | 9 | 9 | 9 | 8 | 9 | 7 | |
| 46,135 | 46,144 | TA | 5 | 5 | 5 | 5 | 5 | 3 | |
| 46,782 | 46,791 | T | 10 | 11 | 10 | 11 | 14 | 10 | |
| 47,368 | 47,378 | A | 11 | 11 | 12 | 12 | 10 | 13 | |
| 47,586 | 47,595 | T | 10 | 10 | 10 | 10 | 13 | TCT8 | |
| 47,624 | 47,633 | A | 10 | 10 | 11 | 11 | 8 | 7 | |
| 49,061 | 49,070 | T | 10 | 10 | 11 | 10 | 8 | T3AT10 | |
| 49,631 | 49,640 | T | 10 | 10 | 10 | 10 | 8 | 8 | |
| 50,329 | 50,340 | A | 12 | 12 | 12 | 12 | 11 | 11 | |
| 51,202 | 51,211 | TA | 5 | 5 | 5 | 5 | 19nt | deletion | |
| 51,215 | 51,230 | T | 16 | 17 | 17 | 16 | 13 | 13 | |
| 53,088 | 53,097 | T | CDS | 10 | 10 | 10 | 10 | 10 | 12 |
| 53,592 | 53,601 | C | 10 | 11 | 9 | 12 | 9 | 9 | |
| 55,477 | 55,490 | T | 14 | 14 | 14 | 14 | complement A11 | complement A13 | |
| 55,891 | 55,906 | T | 16 | 16 | 16 | 16 | 13 | 15 | |
| 56,476 | 56,485 | T | 10 | 10 | 10 | 10 | 10 | A2T8 | |
| 58,301 | 58,310 | T | 10 | 10 | 10 | 10 | 10 | 6 | |
| 59,338 | 59,348 | T | 11 | 10 | 9 | 11 | 11 | 4 | |
| 61,731 | 61,739 | C | 9 | 13 | 9 | 12 | 8 | C3AC3 | |
| 62,108 | 62,117 | TA | 5 | 5 | 5 | 5 | 6 | 4 | |
| 62,161 | 62,182 | T | 22 | 22 | 22 | 31 | 2nt shorter | 2nt shorter | |
| 62,202 | 62,210 | A | 9 | 10 | 9 | 9 | A5TA3 | A5TA3 | |
| 63,523 | 63,538 | T | 16 | 15 | 15 | T5GT10 | 16 | 7 | |
| 64,629 | 64,639 | T | 11 | 11 | 11 | 11 | 11 | T6GT3G | |
| 65,636 | 65,645 | C | 10 | 13 | 11 | 13 | 8 | C2TCTGC7 | |
| 66,253 | 66,262 | AT | 5 | 5 | 5 | 5 | 4 | 7 | |
| 66,851 | 66,864 | A | 14 | 14 | 14 | 19 | 17 | 12 | |
| 68,965 | 68,977 | T | 13 | 13 | 13 | 13 | 11 | 11 | |
| 69,965 | 69,975 | T | 11 | 11 | 12 | 11 | 11 | 8 | |
| 75,328 | 75,340 | A | 13 | 14 | 14 | 13 | 19 | 14 | |
| 76,614 | 76,626 | T | 13 | 13 | 13 | 13 | 13 | 13 | |
| 78,154 | 78,162 | TTG | 3 | 5 | 3 | 3 | 4 | 2 | |
| 80,484 | 80,493 | A | 10 | 11 | 10 | 10 | 10 | 9 | |
| 81,019 | 81,035 | T | 17 | 17 | 17 | 17 | 17 | 17 | |
| 81,178 | 81,191 | T | 14 | 14 | 14 | 14 | 18 | 8 | |
| 82,568 | 82,578 | A | 11 | 10 | 9 | 10 | 9 | 10 | |
| 83,489 | 83,498 | TA | 5 | 5 | 5 | 5 | 5 | 4 | |
| 93,127 | 93,136 | TA | 5 | 5 | 5 | 5 | 5 | 4 | |
| 97,975 | 97,984 | A | 10 | 10 | 10 | 10 | 12 | 9 | |
| 98,781 | 98,791 | T | 11 | 11 | 11 | 11 | 10 | 14 | |
| 107,287 | 107,295 | AT | 5 | 5 | 5 | 5 | 5 | 7 | |
| 107,313 | 107,324 | T | 12 | 11 | 13 | 13 | T2(AT)4T7 | 14 | |
| 111,481 | 111,490 | TA | 5 | 5 | 5 | 5 | TA2TGTA | 4 | |
| 111,589 | 111,598 | AT | 5 | 5 | 5 | 5 | 5 | 10 | |
| 111,665 | 111,672 | T | 8 | 8 | 10 | 8 | 7 | 10 | |
| 111,801 | 111,810 | A | A7CA2 | A7CA2 | 10 | A7CA2 | A7CA2 | A7TAC | |
| 112,472 | 112,481 | A | 10 | 10 | 10 | 10 | 11 | 10 | |
| 116,836 | 116,845 | T | 10 | 9 | 10 | 11 | T7AT3 | 10 | |
| 123,173 | 123,184 | T | 12 | 12 | 12 | 12 | 12 | 12 | |
| 123,285 | 123,383 | T | 10 | 10 | 10 | 10 | 10 | 10 | |
| 123,884 | 123,893 | T | 10 | 10 | 10 | 10 | 10 | 10 | |
| 123,975 | 123,987 | A | 13 | 13 | 13 | 13 | 13 | 13 | |
| 124,356 | 124,365 | TA | 5 | 5 | 5 | 5 | 5 | 5 | |
| 124,874 | 124,886 | T | 13 | 13 | 13 | 13 | 13 | 13 | |
| 125,029 | 125,041 | A | 13 | 13 | 13 | 13 | 13 | 13 | |
| 126,052 | 125,385 | T | 15 | 15 | 15 | 15 | 15 | 17 | |
| 126,087 | 126,097 | T | 11 | 11 | 11 | 11 | 11 | 11 | |
| 126,117 | 126,128 | A | 12 | 12 | 12 | 12 | 12 | 12 | |
| 126,952 | 126,962 | T | 11 | 11 | 11 | 11 | T8CT2 | T8CT2 | |
| 127,241 | 127,252 | A | 12 | 12 | 12 | 12 | 6 | 6 | |
| Number of Mapped Reads | |||||
|---|---|---|---|---|---|
| 5 and More | 4 and More | 3 and More | 2 and More | ||
| Number of variable sites | Total | 29 | 46 | 74 | 129 |
| Specific to | A. nipponica | 1 | 1 | 4 | 12 |
| A. flagellosa | 0 | 0 | 0 | 3 | |
| A. hirsuta | 14 | 25 | 35 | 62 | |
| Shared with | A. flagellosa and A. nipponica | 14 | 19 | 31 | 46 |
| A. nipponica and A. hirsuta | 0 | 0 | 1 | 1 | |
| A. flagellosa and A. hirsuta | 0 | 0 | 1 | 1 | |
| other type | 0 | 1 | 2 | 4 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawabe, A.; Nukii, H.; Furihata, H.Y. Exploring the History of Chloroplast Capture in Arabis Using Whole Chloroplast Genome Sequencing. Int. J. Mol. Sci. 2018, 19, 602. https://doi.org/10.3390/ijms19020602
Kawabe A, Nukii H, Furihata HY. Exploring the History of Chloroplast Capture in Arabis Using Whole Chloroplast Genome Sequencing. International Journal of Molecular Sciences. 2018; 19(2):602. https://doi.org/10.3390/ijms19020602
Chicago/Turabian StyleKawabe, Akira, Hiroaki Nukii, and Hazuka Y. Furihata. 2018. "Exploring the History of Chloroplast Capture in Arabis Using Whole Chloroplast Genome Sequencing" International Journal of Molecular Sciences 19, no. 2: 602. https://doi.org/10.3390/ijms19020602
APA StyleKawabe, A., Nukii, H., & Furihata, H. Y. (2018). Exploring the History of Chloroplast Capture in Arabis Using Whole Chloroplast Genome Sequencing. International Journal of Molecular Sciences, 19(2), 602. https://doi.org/10.3390/ijms19020602