Nitrate Reductases Are Relocalized to the Nucleus by AtSIZ1 and Their Levels Are Negatively Regulated by COP1 and Ammonium

 ,
, 
Abstract
1. Introduction
2. Results
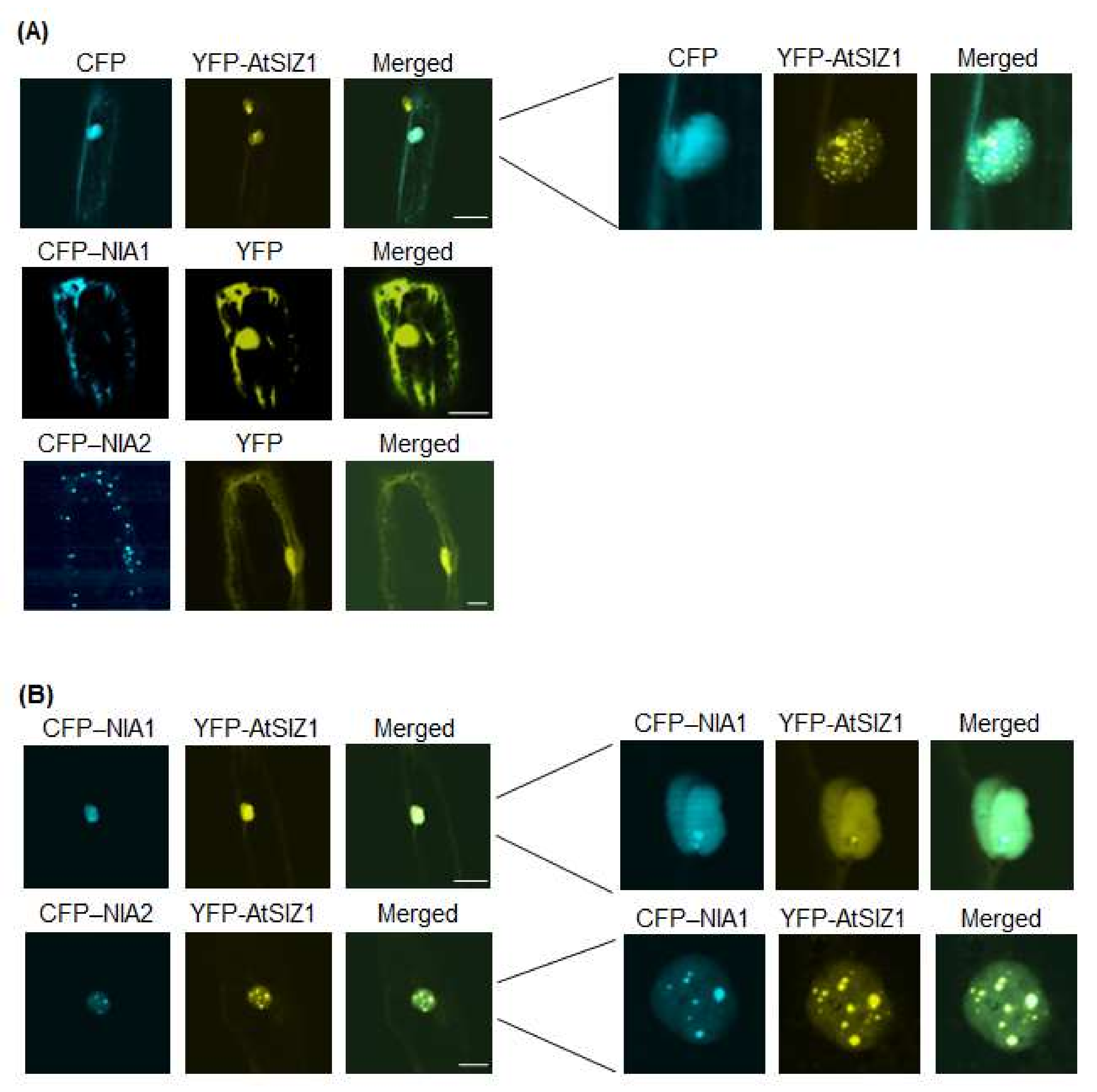
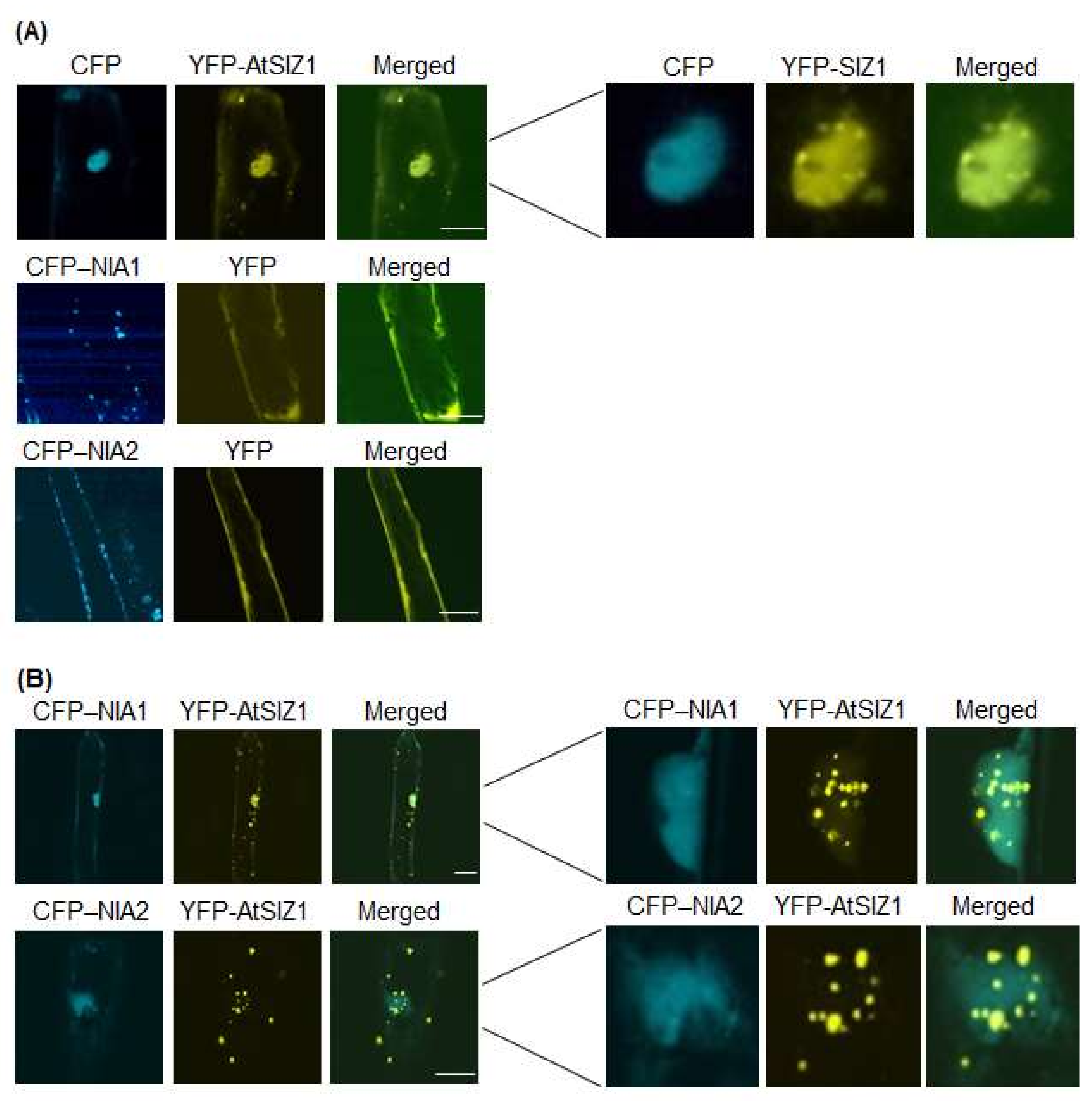
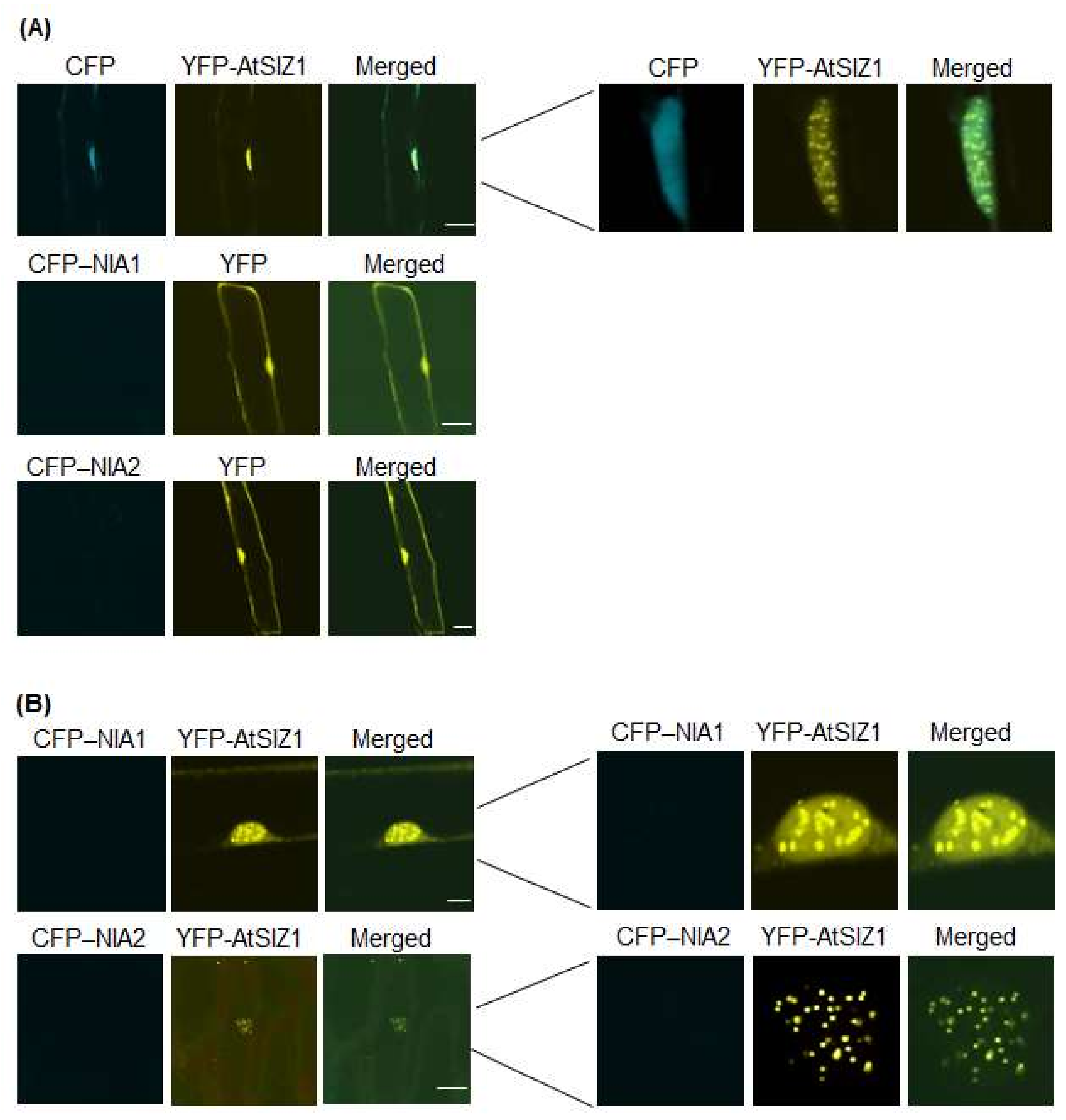
2.1. NR Proteins and AtSIZ1 Localize to the Nucleus
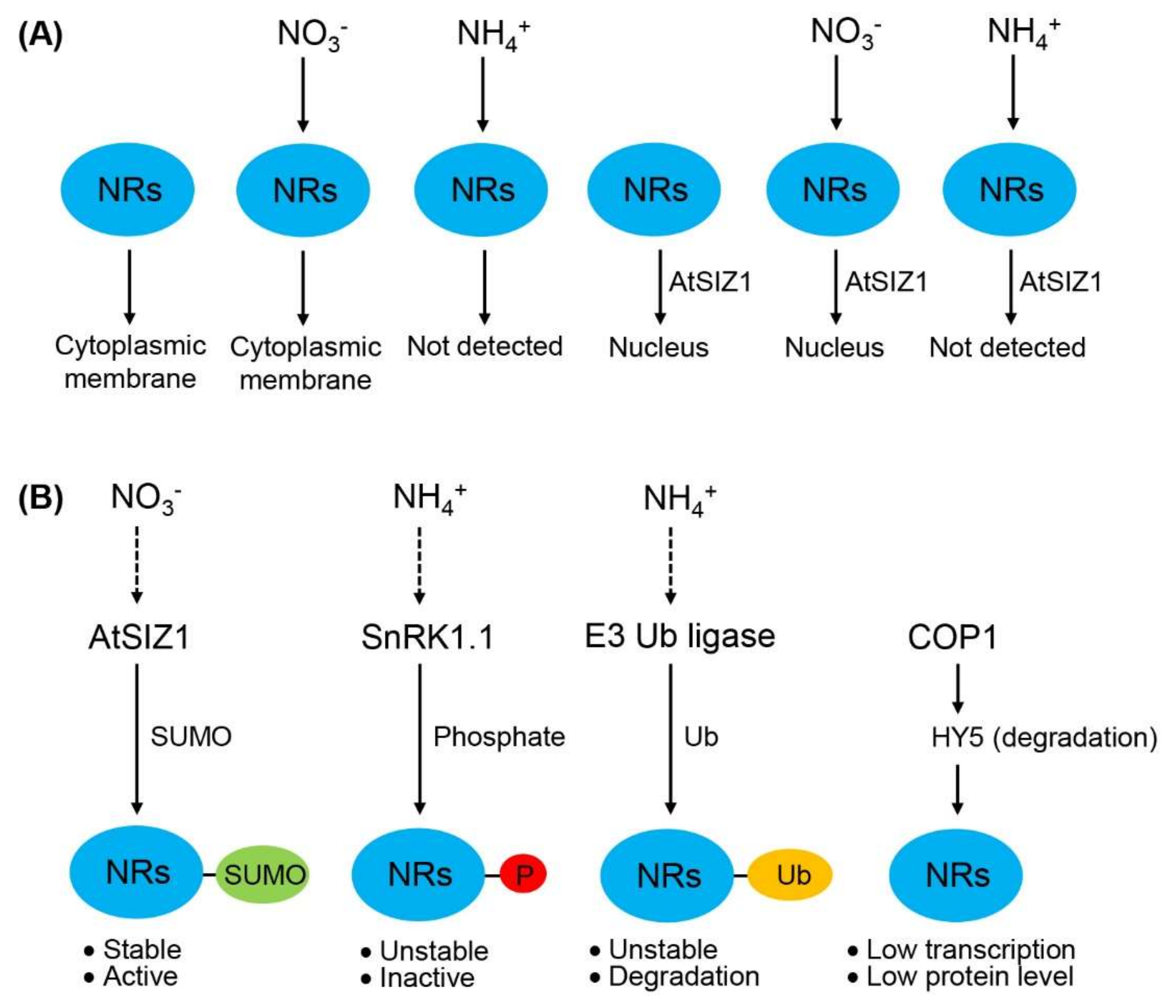
2.2. Nuclear Localization of NR Proteins Is Not Affected by Nitrate
2.3. NIA1 and NIA2 Are Not Detected in Ammonium-Treated Cells
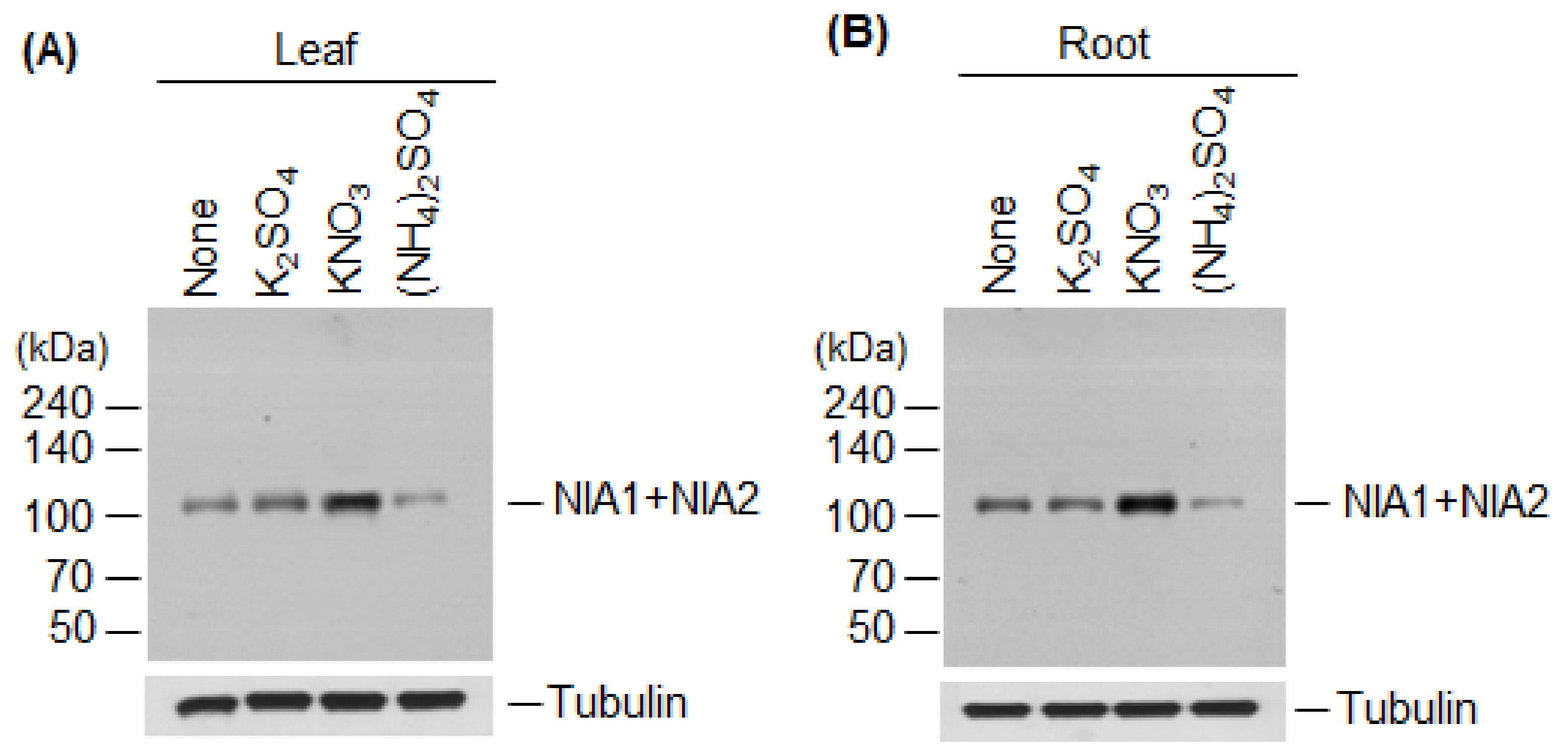
2.4. NIA1 and NIA2 Levels Are Regulated by Nitrate and Ammonium Sources
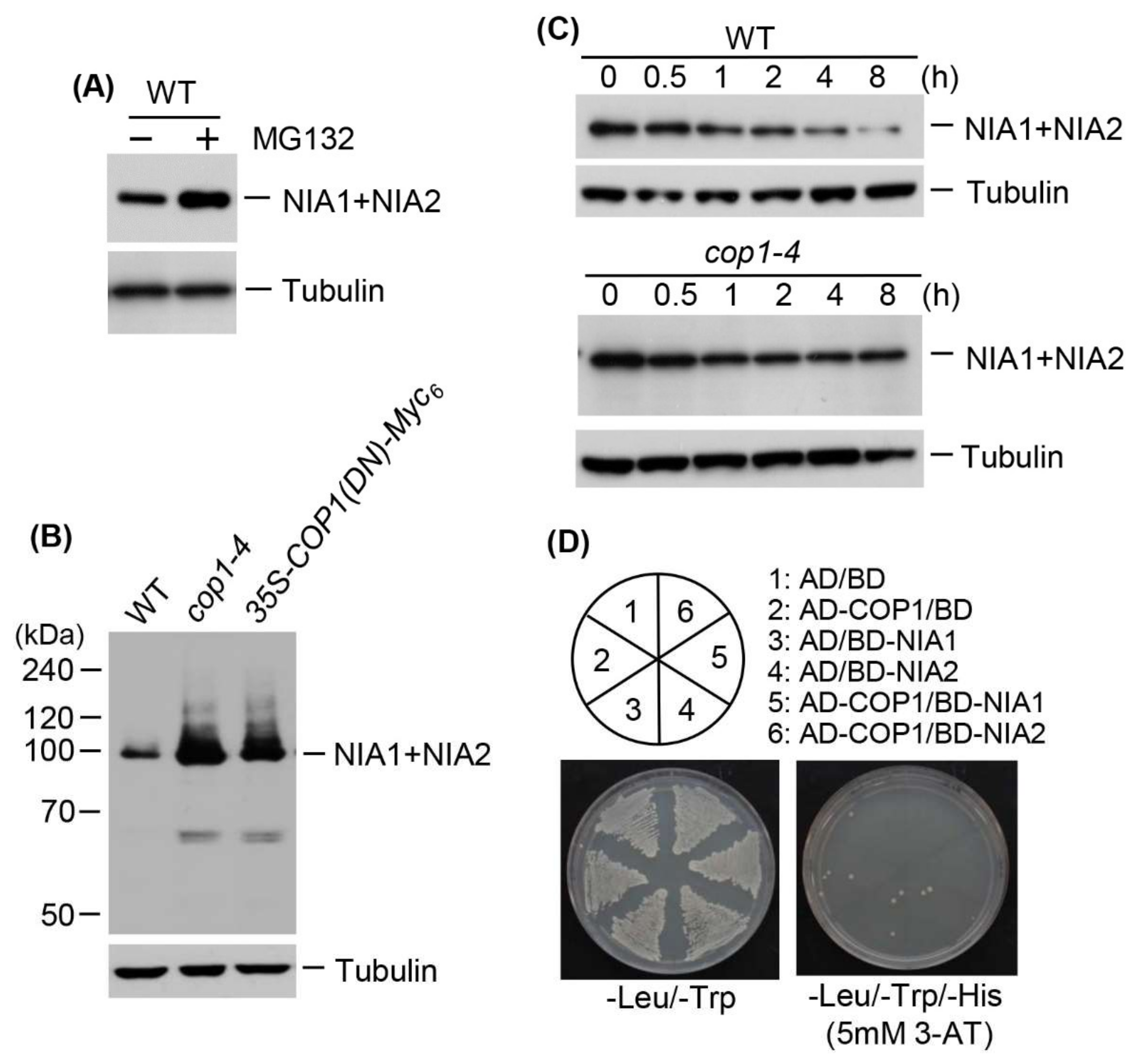
2.5. NR Proteins Accumulate in cop1-4 and DN-COP1-Overexpressing Plants
3. Discussion
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Subcellular Localization
4.3. Quantitative RT-PCR
4.4. Examination of NR Protein Levels
4.5. Cell-Free Degradation Assay
4.6. Yeast Two-Hybrid Experiments
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Campbell, W.H.; Kinghorn, K.R. Functional domains of assimilatory nitrate reductases and nitrite reductases. Trends Biochem. Sci. 1990, 15, 315–319. [Google Scholar] [CrossRef]
- Solomonson, L.P.; Barber, M.J. Assimilatory nitrate reductase-functional properties and regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 225–253. [Google Scholar] [CrossRef]
- Tanaka, T.; Ida, S.; Irifune, K.; Oeda, K.; Morikawa, H. Nucleotide sequence of a gene for nitrite reductase from Arabidopsis thaliana. DNA Seq. 1994, 5, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Seligman, K.; Saviani, E.E.; Oliveira, H.C.; Pinto-Maglio, C.A.; Salgado, I. Floral transition and nitric oxide emission during flower development in Arabidopsis thaliana is affected in nitrate reductase-deficient plants. Plant Cell Physiol. 2008, 49, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.K. A new RING for SUMO: Wrestling transcriptional responses into nuclear bodies with PIAS family E3 SUMO ligases. Genes Dev. 2001, 15, 3053–3058. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, K.A.; Henley, J.M. Mechanisms, regulation and consequences of protein sumoylation. Biochem. J. 2010, 428, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Dikic, I. Atypical ubiquitin chains: New molecular signals. Protein modifications: Beyond the usual suspects’ review series. EMBO Rep. 2008, 9, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.J.; Barrett-Wilt, G.A.; Hua, Z.; Vierstra, R.D. Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 16512–16517. [Google Scholar] [CrossRef] [PubMed]
- Elrouby, N.; Coupland, G. Proteome-wide screens for small ubiquitin-like modifier (SUMO) substrates identify Arabidopsis proteins implicated in diverse biological processes. Proc. Natl. Acad. Sci. USA 2010, 107, 17415–17420. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Hasegawa, P.M. Sumoylation and other ubiquitin-like post-translational modifications in plants. Trends Cell Biol. 2010, 20, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Park, B.S.; Song, J.T.; Seo, H.S. FLC-mediated flowering repression is positively regulated by sumoylation. J. Exp. Bot. 2014, 65, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.S.; Son, G.H.; Kim, S.I.; Song, J.T.; Seo, H.S. Arabidopsis HIGH PLOIDY2 sumoylates and stabilizes Fowering Locus C through its E3 ligase activity. Front. Plant Sci. 2016, 7, 530. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Han, Y.J.; Kim, S.I.; Song, J.T.; Seo, H.S. Arabidopsis CMT3 activity is positively regulated by AtSIZ1-mediated sumoylation. Plant Sci. 2015, 239, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Park, B.S.; Kim, D.Y.; Yeu, S.Y.; Song, S.I.; Song, J.T.; Seo, H.S. E3 SUMO ligase AtSIZ1 positively regulates SLY1-mediated GA signalling and plant development. Biochem. J. 2015, 469, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Kwak, J.S.; Song, J.T.; Seo, H.S. The E3 SUMO ligase AtSIZ1 functions in seed germination in Arabidopsis. Physiol. Plant. 2016, 158, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Rus, A.; Sharkhuu, A.; Yokoi, S.; Karthikeyan, A.S.; Raghothama, K.G.; Baek, D.; Koo, Y.D.; Jin, J.B.; Bressan, R.A.; et al. The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency responses. Proc. Natl. Acad. Sci. USA 2005, 102, 7760–7765. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Song, J.T.; Seo, H.S. Arabidopsis nitrate reductase activity is stimulated by the E3 SUMO ligase AtSIZ1. Nat. Commun. 2011, 2, 400. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.B.; Jin, Y.H.; Lee, J.; Miura, K.; Yoo, C.Y.; Kim, W.Y.; Van Oosten, M.; Hyun, Y.; Somers, D.E.; Lee, I.; et al. The SUMO E3 ligase, AtS1Z1, regulates flowering by controlling a salicylic acid-mediated floral promotion pathway and through affects on FLC chromatin structure. Plant J. 2008, 53, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Yoo, C.Y.; Miura, K.; Jin, J.B.; Lee, J.; Park, H.C.; Salt, D.E.; Yun, D.J.; Bressan, R.A.; Hasegawa, P.M. SIZ1 small ubiquitin-like modifier E3 ligase facilitates basal thermotolerance in Arabidopsis independent of salicylic acid. Plant Physiol. 2006, 142, 1548–1558. [Google Scholar] [CrossRef] [PubMed]
- Catala, R.; Ouyang, J.; Abreu, I.A.; Hu, Y.; Seo, H.; Zhang, X.; Chua, N.H. The Arabidopsis E3 SUMO ligase SIZ1 regulates plant growth and drought responses. Plant Cell 2007, 19, 2952–2966. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Jin, J.B.; Lee, J.; Yoo, C.Y.; Stirm, V.; Miura, T.; Ashworth, E.N.; Bressan, R.A.; Yun, D.J.; Hasegawa, P.M. SIZ1-mediated sumoylation of ICE1 controls CBF3/DREB1A expression and freezing tolerance in Arabidopsis. Plant Cell 2007, 19, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Sato, A.; Ohta, M.; Furukawa, J. Increased tolerance to salt stress in the phosphate-accumulating Arabidopsis mutants siz1 and pho2. Planta 2011, 234, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Nozawa, R. Overexpression of SIZ1 enhances tolerance to cold and salt stresses and attenuates response to abscisic acid in Arabidopsis thaliana. Plant Biotechnol. 2014, 31, 167–172. [Google Scholar] [CrossRef][Green Version]
- Kim, J.Y.; Jang, I.C.; Seo, H.S. COP1 Controls abiotic stress responses by modulating AtSIZ1 function through its E3 ubiquitin ligase activity. Front. Plant Sci. 2016, 7, 1182. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Song, J.T.; Seo, H.S. Post-translational modifications of Arabidopsis E3 SUMO ligase AtSIZ1 are controlled by environmental conditions. FEBS Open Bio 2017, 7, 1622–1634. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Lee, J.; Miura, T.; Hasegawa, P.M. SIZ1 controls cell growth and plant development in Arabidopsis through salicylic acid. Plant Cell Physiol. 2010, 51, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.M.; Brendle-Behnisch, E. Rapid modulation of spinach leaf nitrate reductase activity by photosynthesis I. Modulation in vivo by CO2 availability. Plant Physiol. 1991, 96, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.L.; Huber, S.C.; Campbell, W.H.; Redinbaugh, M.G. Reversible light/dark modulation of spinach leaf nitrate reductase activity involves protein phosphorylation. Arch. Biochem. Biophys. 1992, 296, 58–65. [Google Scholar] [CrossRef]
- Huber, J.L.; Redinbaugh, M.G.; Huber, S.C.; Campbell, W.H. Regulation of maize leaf nitrate reductase activity involves both gene expression and protein phosphorylation. Plant Physiol. 1994, 106, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Weiner, H.; Kaiser, W.M. 14-3-3 proteins control proteolysis of nitrate reductase in spinach leaves. FEBS Lett. 1994, 455, 75–78. [Google Scholar] [CrossRef]
- Jossier, M.; Bouly, J.P.; Meimoun, P.; Arjmand, A.; Lessard, P.; Hawley, S.; Grahame Hardie, D.; Thomas, M. SnRK1 (SNF1-related kinase 1) has a central role in sugar and ABA signalling in Arabidopsis thaliana. Plant J. 2009, 59, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M. Nitrate: Nutrient and Signal for Plant Growth. Plant Cell 1995, 7, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Saracco, S.A.; Miller, M.J.; Kurepa, J.; Vierstra, R.D. Genetic analysis of SUMOylation in Arabidopsis: Conjugation of SUMO1 and SUMO2 to nuclear proteins is essential. Plant Physiol. 2007, 145, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Konishi, M.; Yanagisawa, S. The regulatory region controlling the nitrate-responsive expression of a nitrate reductase gene, NIA1, in Arabidopsis. Plant Cell Physiol. 2011, 52, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kwon, Y.J.; Kim, S.I.; Kim, D.Y.; Song, J.T.; Seo, H.S. Ammonium inhibits chromomethylase 3-mediated methylation of the Arabidopsis nitrate reductase gene NIA2. Front. Plant Sci. 2016, 6, 1161. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.-W.; Caspar, T.; Quail, P.H. cop1: A regulatory locus involved in light-controlled development and gene expression in Arabidopsis. Genes Dev. 1991, 5, 1172–1182. [Google Scholar] [CrossRef] [PubMed]
- Jonassen, E.M.; Sandsmark, B.A.; Lillo, C. Unique status of NIA2 in nitrate assimilation: NIA2 expression is promoted by HY5/HYH and inhibited by PIF4. Plant Signal. Behav. 2009, 4, 1084–1086. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Song, J.T.; Seo, H.S. COP1 regulates plant growth and development in response to light at the post-translational level. J. Exp. Bot. 2017, 68, 4737–4748. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S.; Watanabe, E.; Tokutomi, S.; Nagatani, A.; Chua, N.H. Photoreceptor ubiquitination by COP1 E3 ligase desensitizes phytochrome A signaling. Genes Dev. 2004, 18, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, K.C.; Campbell, W.H. Immunogold localization of nitrate reductase in maize leaves. Plant Physiol. 1988, 88, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Małolepsza, U. Nitric oxide production in plants. Postepy Biochem. 2007, 53, 263–271. [Google Scholar] [PubMed]
- Stacey, M.G.; Kopp, O.R.; Kim, T.H.; von Arnim, A.G. Modular domain structure of Arabidopsis COP1. Reconstitution of activity by fragment complementation and mutational analysis of a nuclear localization signal in planta. Plant Physiol. 2000, 124, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Bauer, N.C.; Doetsch, P.W.; Corbett, A.H. Mechanisms regulating protein localization. Traffic 2015, 16, 1039–1061. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Romão, M.J. Structural and mechanistic insights on nitrate reductases. Protein Sci. 2015, 24, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Cheong, M.S.; Park, H.C.; Hong, M.J.; Lee, J.; Choi, W.; Jin, J.B.; Bohnert, H.J.; Lee, S.Y.; Bressan, R.A.; Yun, D.J. Specific domain structures control abscisic acid-, salicylic acid-, and stress-mediated SIZ1 phenotypes. Plant Physiol. 2009, 151, 1930–1942. [Google Scholar] [CrossRef] [PubMed]
- Park, H.C.; Kim, H.; Koo, S.C.; Park, H.J.; Cheong, M.S.; Hong, H.; Baek, D.; Chung, W.S.; Kim, D.H.; Bressan, R.A.; et al. Functional characterization of the SIZ/PIAS-type SUMO E3 ligases, OsSIZ1 and OsSIZ2 in rice. Plant Cell Environ. 2010, 33, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Thangasamy, S.; Guo, C.L.; Chuang, M.H.; Lai, M.H.; Chen, J.; Jauh, G.Y. Rice SIZ1, a SUMO E3 ligase, controls spikelet fertility through regulation of anther dehiscence. New Phytol. 2011, 189, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Lambeck, I.C.; Fischer-Schrader, K.; Niks, D.; Roeper, J.; Chi, J.C.; Hille, R.; Schwarz, G. Molecular mechanism of 14-3-3 protein-mediated inhibition of plant nitrate reductase. J. Biol. Chem. 2012, 287, 4562–4571. [Google Scholar] [CrossRef] [PubMed]
- Osterlund, M.T.; Hardtke, C.S.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Sullivan, J.A.; Wang, H.; Yang, J.; Shen, Y.; Rubio, V.; Ma, L.; Hoecker, U.; Deng, X.W. The COP1-SPA1 interaction defines a critical step in phytochrome A-mediated regulation of HY5 activity. Genes Dev. 2003, 17, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Huang, X.; Gusmaroli, G.; Terzaghi, W.; Lau, O.S.; Yanagawa, Y.; Zhang, Y.; Li, J.; Lee, J.H.; Zhu, D.; et al. Arabidopsis CULLIN4-damaged DNA binding protein 1 interacts with CONSTITUTIVELY PHOTOMORPHOGENIC1-SUPPRESSOR OF PHYA complexes to regulate photomorphogenesis and flowering time. Plant Cell 2010, 22, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Avaze’ri, C.; Turner, R.J.; Pommier, J.; Weiner, J.H.; Giordano, G.; Verme’glio, A. Tellurite reductase activity of nitrate reductase is responsible for the basal resistance of Escherichia coli to tellurite. Microbiology 1997, 143, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Ganther, H.E. Selenium metabolism, selenoproteins and mechanisms of cancer prevention: Complexities with thioredoxin reductase. Carcinogenesis 1999, 20, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Sabaty, M.; Avaze’ri, C.; Pignol, D.; Verme’glio, A. Characterization of the Reduction of Selenate and Tellurite by Nitrate Reductases. Appl. Environ. Microbiol. 2001, 67, 5122–5126. [Google Scholar] [CrossRef] [PubMed]
- Lehotai, N.; Kolbert, Z.; Peto, A.; Feigl, G.; Ördög, A.; Kumar, D.; Tari, I.; Erdei, L. Selenite-induced hormonal and signalling mechanisms during root growth of Arabidopsis thaliana L. J. Exp. Bot. 2012, 63, 5677–5687. [Google Scholar] [CrossRef] [PubMed]
- Bao, P.; Huang, H.; Hu, Z.Y.; Häggblom, M.M.; Zhu, Y.G. Impact of temperature, CO2 fixation and nitrate reduction on selenium reduction, by a paddy soil Clostridium strain. J. Appl. Microbiol. 2013, 114, 703–712. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf | Root | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| None | K2SO4 | KNO3 | (NH4)2SO4 | None | K2SO4 | KNO3 | (NH4)2SO4 | ||
| NIA1 | 1.00 ± 0.18 | 1.02 ± 0.15 | 3.33 ± 0.27 | 2.23 ± 0.24 | 1.00 ± 0.12 | 1.10 ± 0.21 | 1.62 ± 0.44 | 1.56 ± 0.07 | |
| NIA2 | 1.00 ± 0.19 | 1.01 ± 0.13 | 2.00 ± 0.11 | 1.36 ± 0.12 | 1.00 ± 0.13 | 0.98 ± 0.14 | 1.59 ± 0.36 | 1.34 ± 0.08 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.Y.; Park, B.S.; Park, S.W.; Lee, H.Y.; Song, J.T.; Seo, H.S. Nitrate Reductases Are Relocalized to the Nucleus by AtSIZ1 and Their Levels Are Negatively Regulated by COP1 and Ammonium. Int. J. Mol. Sci. 2018, 19, 1202. https://doi.org/10.3390/ijms19041202
Kim JY, Park BS, Park SW, Lee HY, Song JT, Seo HS. Nitrate Reductases Are Relocalized to the Nucleus by AtSIZ1 and Their Levels Are Negatively Regulated by COP1 and Ammonium. International Journal of Molecular Sciences. 2018; 19(4):1202. https://doi.org/10.3390/ijms19041202
Chicago/Turabian StyleKim, Joo Yong, Bong Soo Park, Sang Woo Park, Han Yong Lee, Jong Tae Song, and Hak Soo Seo. 2018. "Nitrate Reductases Are Relocalized to the Nucleus by AtSIZ1 and Their Levels Are Negatively Regulated by COP1 and Ammonium" International Journal of Molecular Sciences 19, no. 4: 1202. https://doi.org/10.3390/ijms19041202
APA StyleKim, J. Y., Park, B. S., Park, S. W., Lee, H. Y., Song, J. T., & Seo, H. S. (2018). Nitrate Reductases Are Relocalized to the Nucleus by AtSIZ1 and Their Levels Are Negatively Regulated by COP1 and Ammonium. International Journal of Molecular Sciences, 19(4), 1202. https://doi.org/10.3390/ijms19041202