1. Introduction
With an increasing concern about the negative impact of certain antibiotic use aspects on public health, many countries are gradually banning the use of antibiotics in animal production. Thus, searching for potential alternatives to antibiotics is pivotal for sustainable development of animal production. To date, many research reports have been focused on the plant extracts as they are natural and abundant with strong antibacterial, antifungal and antiviral activities [
1,
2]. These phytochemicals can disturb microbial cell structures, increase bacterial cell permeability, and consequently result in bacterial death [
2]. Essential oils, one kind of the plant extracts, are commonly obtained by steam distillation and have been used in food preservation, pharmaceutical therapies, alternative medicine, and natural therapies for many years [
3].
Oleum cinnamomi (OCM) a type of essential oil commonly used in the food industry because of its special aroma [
4]. Previous studies have shown the antibacterial activity of OCM that was extracted from both the bark and leaves of
Cinnamomum burmannii and
Cinnamomun osmophloeum [
3,
4]. The predominant bioactive compound in OCM was reported to be cinnamaldehyde, a potent inhibitor of bacteria, yeast, and filamentous molds [
5]. The underlying mechanisms through which cinnamaldehyde exerts the inhibitory effects were possibly via inhibition of microbial ATPase activity and cell wall biosynthesis, and changes in bacterial membrane structure and integrity [
5]. Cinnamaldehyde was also developed for medicinal purposes, including the use as anti-inflammatory, anti-emetic and analgesic curatives [
6]. There were virtually no side effects reported in cinnamaldehyde application. Gowder [
7] found that the acute toxicity of cinnamaldehyde is low, with lethal dose 50% (LD
50) values ranging from 0.6 to more than 2 g per kg of body weight (BW) in various animal species. Specifically, the oral LD
50 values of cinnamaldehyde were 1.16 g/kg BW and 2.22 g/kg BW for pigs and rats, respectively [
8].
Collectively, there is a paucity of literature on the application of OCM in animal production, although cinnamaldehyde has been extensively studied in human medicine. Our previous study has reported that dietary supplementation with 50 mg/kg OCM attenuated the intestinal injury induced by lipopolysaccharide in piglets [
9]. To extend the findings of our previous work, the present study was carried out to determine the effects of OCM on intestinal gene expression and function in piglets.
3. Discussion
There is an urgent need to develop antibiotics-free feed, which aims at stopping the spread of antibiotic resistance and can also maintain the current level of animal production. In the present study, as shown by the increased average daily feed intake (ADFI) and reduced diarrhoea incidence, dietary supplementation with 50 mg/kg OCM improved the growth performance of pigs. Similarly, Wang et al. [
9] reported that OCM supplementation increased ADFI and average daily gain (ADG), but reduced diarrhoea incidence in pigs that had been challenged with lipopolysaccharide (LPS). The active ingredient of OCM is cinnamaldehyde, which was reported to enhance food intake due to the increased expression of hypothalamic neuropeptide Y in mice [
10] and rats [
11]. Therefore, dietary OCM may serve as a potential appetite-enhancing feedstuff in weaning piglets. Additionally, concentrations of plasma insulin were increased by the OCM diet, showing that OCM may promote anabolic metabolism in piglets.
In the present study, dietary supplementation with OCM decreased diarrhoea incidence, indicating that OCM may have beneficial effects on intestinal function. Specifically, OCM supplementation improved the intestinal histology since both villus width and villous surface area were increased in the duodenum and jejunum of pigs in the OCM group. An increase in villous surface area is expected to enhance the digestion and absorption of nutrients [
12]. Thus, the OCM diet may augment the bioavailability of dietary nutrients in young pigs. Moreover, dietary OCM supplementation increased ileal DNA concentrations and RNA/DNA ratios, both of which can be used to assess intestinal development [
13]. DNA concentration reflects the rate of mitosis for producing new columnar epithelial cells, whereas an RNA/DNA ratio indicates cellular efficiency and a protein/DNA ratio implicates the efficiency of protein synthesis in cells [
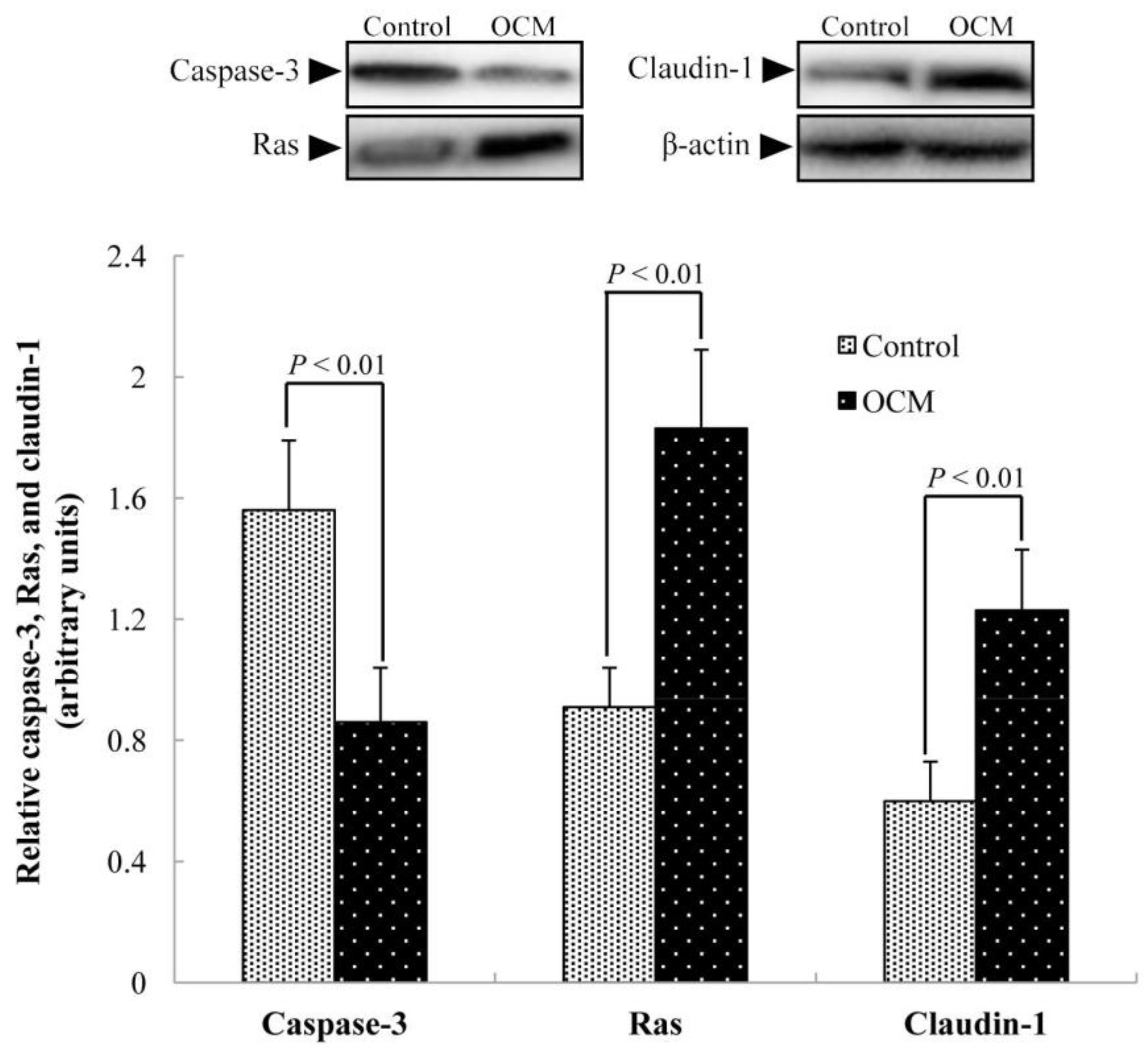
14]. Therefore, our results demonstrated that dietary OCM supplementation could stimulate the growth of the intestinal mucosa. This notion was substantiated by the results that OCM decreased the intestinal abundance of the caspase-3 protein, which is one of the key components of the apoptotic pathway in the gut [
15].
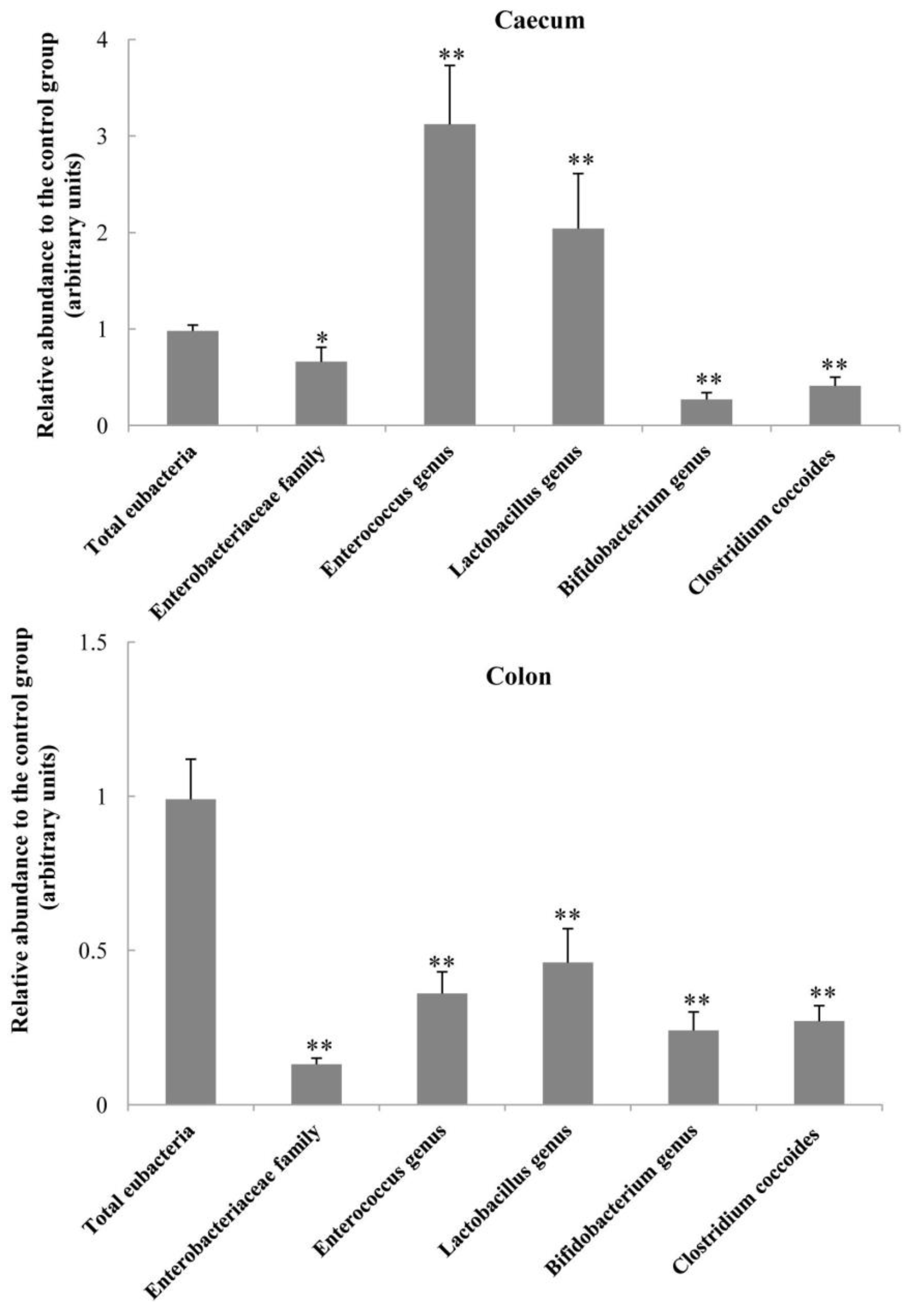
Another important finding in the present study is the alteration of bacterial populations in the intestine by dietary OCM supplementation. The intestinal bacterial mibrobiota contributes to the luminal biological barrier and interacts with the host, thereby affecting the health status of the host [
16]. Intestinal diseases are often associated with the changes in enteric bacterial microbiota, such as the increased proliferation of harmful bacteria and the decreased proliferation of beneficial bacteria [
17]. In the present study, pigs fed OCM diet exhibited an increase in the populations of
Enterococcus genus and
Lactobacillus genus, but a reduction in the abundance of
Enterobacteriaceae family,
Clostridium coccoides and
Bifidobacterium genus in the caecum digesta (
Figure 1). However, all of these five bacterial groups were decreased in the colon by OCM treatment, although the total bacterial population was not altered. Our results indicate the differential effects of dietary OCM on intestinal microorganisms. Of note, cinnamaldehyde (the main component of OCM) was reported to be an active inhibitor of bacterial growth via inhibiting microbial ATPase activity, cell wall biosynthesis, and altering membrane structure and integrity [
5], whereas OCM had little inhibitory action on
Lactobacillus and
Bifidobacterium [
1]. Similarly, Nieto-Bobadilla et al. [
17] found that oral gavage of CIN-102 (cinnamaldehyde, 86.7%
w/
w) significantly reduced the number of luminal and mucosal enterobacteria, and decreased the percentages of bloody stools and diarrhoea in mice. Our results are in line with previous reports, showing that dietary supplementation of OCM can alter the composition of intestinal bacterial microbiota towards the optimal balance of microbiota and consequently lead to the reduction in diarrhoea incidence of piglets. Collectively, considering the reduced diarrhoea incidence and the improvements of intestinal morphology and barrier function, we suggest that the changes in the abundance of selected microorganisms in the caecum and colon in response to dietary OCM supplementation are beneficial for intestinal health in piglets. Further studies are warranted to identify the specific changes in the gut microbiota at the species level.
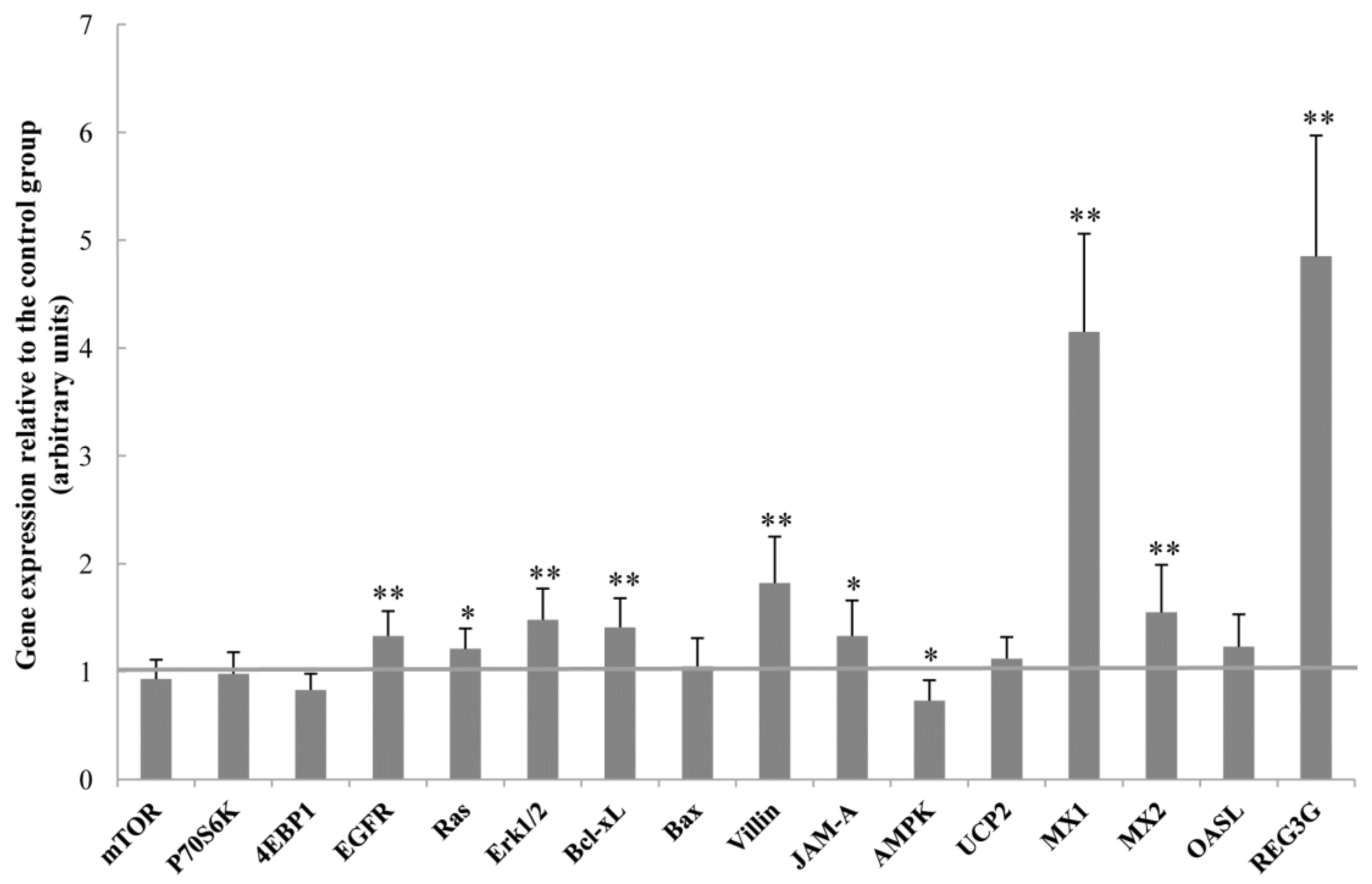
It is also possible that OCM affects intestinal function by regulating the expression of genes associated with intestinal development, intestinal barrier function, energy metabolism, and anti-viral function. Specifically, OCM up-regulated expression Bcl-xL, an anti-apoptotic protein that promotes cell survival [
18], and decreased the abundance of the caspase-3 protein in the jejunum of piglets (
Figure 2 and
Figure 3), indicating that dietary supplementation with OCM could stimulate intestinal cell growth. This notion was further substantiated by the results that OCM increased the mRNA levels of villin. Villin is a marker of villus cell differentiation [
19] and the enhanced expression of villin implies that more villus cells undergo differentiation [
20]. Moreover, dietary OCM supplementation increased the mRNA levels of EGFR, Ras, and Erk1/2 in the jejunum (
Figure 2). Previous studies indicate that EGF and EGFR are two important mediators for enterocyte proliferation and the regeneration of the mucosal epithelium [
21]. Furthermore, the EGFR signaling pathway (the two key downstream molecules are Ras and Erk1/2) was involved in the enhancement of cell proliferation, repair and migration, and stabilization of the internal environment [
22]. Given the elevation of the Ras protein abundance by dietary OCM supplementation (
Figure 3), we speculate that OCM stimulates enterocyte growth via activating the EGFR signalling pathway. More studies are required to test this hypothesis.
Regarding the intestinal barrier function, OCM increased junctional adhesion molecule A (JAM-A) mRNA levels and the abundance of the claudin-1 protein in the jejunum. JAM-A is a critical signalling component of the apical junctional complex, which controls the passage of nutrients and solutes across epithelial surfaces [
23]. The Claudin family of proteins plays an important role in tight junction formation, thereby influencing intestinal permeability [
12]. Thus, OCM may improve intestinal barrier function through regulating intestinal JAM-A and claudin-1 expression. AMPK is a critical regulator of energy metabolism and is activated when the cellular energy level is low [
24]. The current study showed that AMPK expression was decreased by dietary OCM supplementation, indicating that OCM may regulate intestinal energy metabolism. Further research is warranted to test this new hypothesis.
The last but important finding of the present work is that dietary OCM supplementation regulated the mRNA levels of antiviral proteins. The myxovirus resistance (MX) proteins are major effector molecules that prevent influenza-infected animals from developing severe phenotypes [
25]. MX1 is an important downstream effector of type I interferons and has been reported to possess antiviral activity against a variety of RNA viruses [
25]. Our results showed that dietary OCM supplementation increased the mRNA levels for both MX1 and MX2. Therefore, OCM may have the potential to benefit intestinal health by controlling viral infection. Additionally, OCM may consolidate intestinal antimicrobial effect since dietary OCM supplementation dramatically increased the mRNA levels of REG3G, which is expressed predominantly in the small intestine and has been reported to be bactericidal to pathogenic bacteria [
26]. Thus, dietary supplementation with OCM can play an important role in improving the intestinal and whole-body health of young pigs.
In summary, dietary supplementation with 50 mg/kg OCM improved feed intake and and intestinal functions in piglets. The beneficial actions of OCM on intestinal functions include: (1) improvements of intestinal growth and histology (indicated by increased villus height and villous surface area in the duodenum and jejunum; DNA concentrations and RNA/DNA ratios in the ileum; and elevated mRNA levels for EGFR, Ras, Erk1/2, Bcl-xL and villin, and reduced caspase-3 protein abundance); (2) increases in intestinal mucosal barrier function (shown by increased mRNA levels for JAM-A and claudin-1 protein abundance); (3) alterations in the composition of intestinal bacterial microbiota (demonstrated by the reduced population of Enterococcus, Enterobacterium and Clostridium in the colon, and increased abundance of Enterococcus and Lactobacillus in the caecum); (4) enhancement of antivirus function (indicated by elevated mRNA levels for MX1, MX2, and REG3G); and (5) the reduced incidence of diarrhoea. OCM is a low-cost feed additive and alternative to feed antibiotics for improving intestinal health in weanling pigs.





{kind=link}
{kind=link}
{kind=link}