MCPIP3 as a Potential Metastasis Suppressor Gene in Human Colorectal Cancer

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
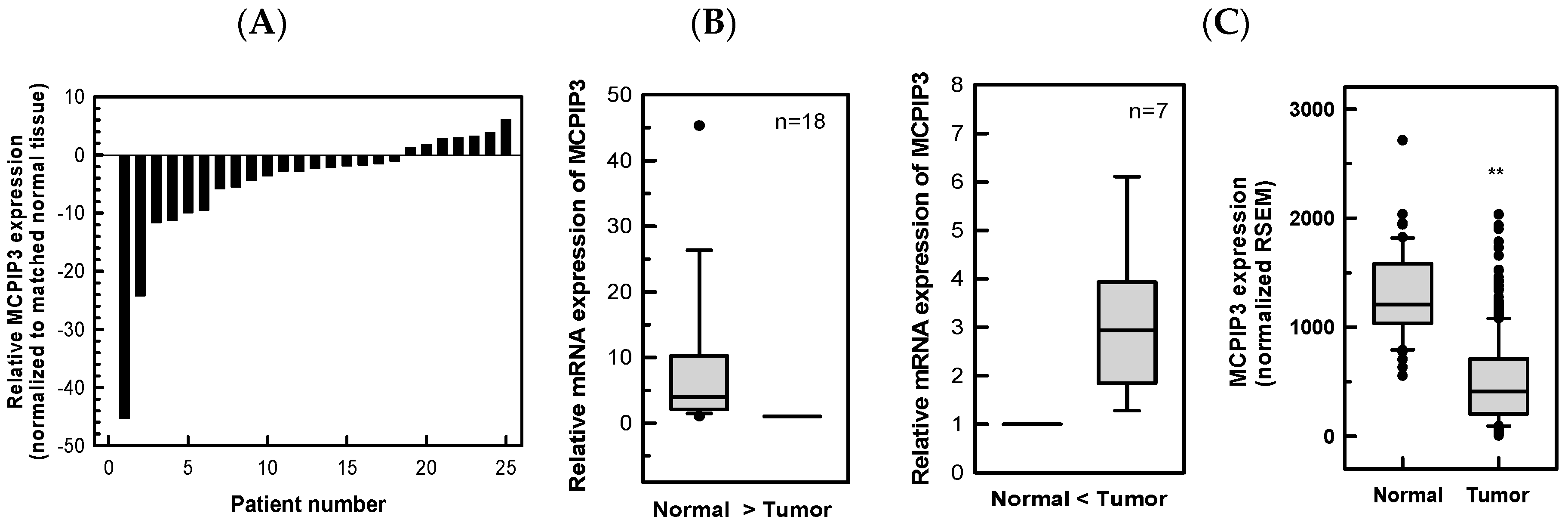
2.1. Downregulation of MCPIP3 in Human CRC Tissues
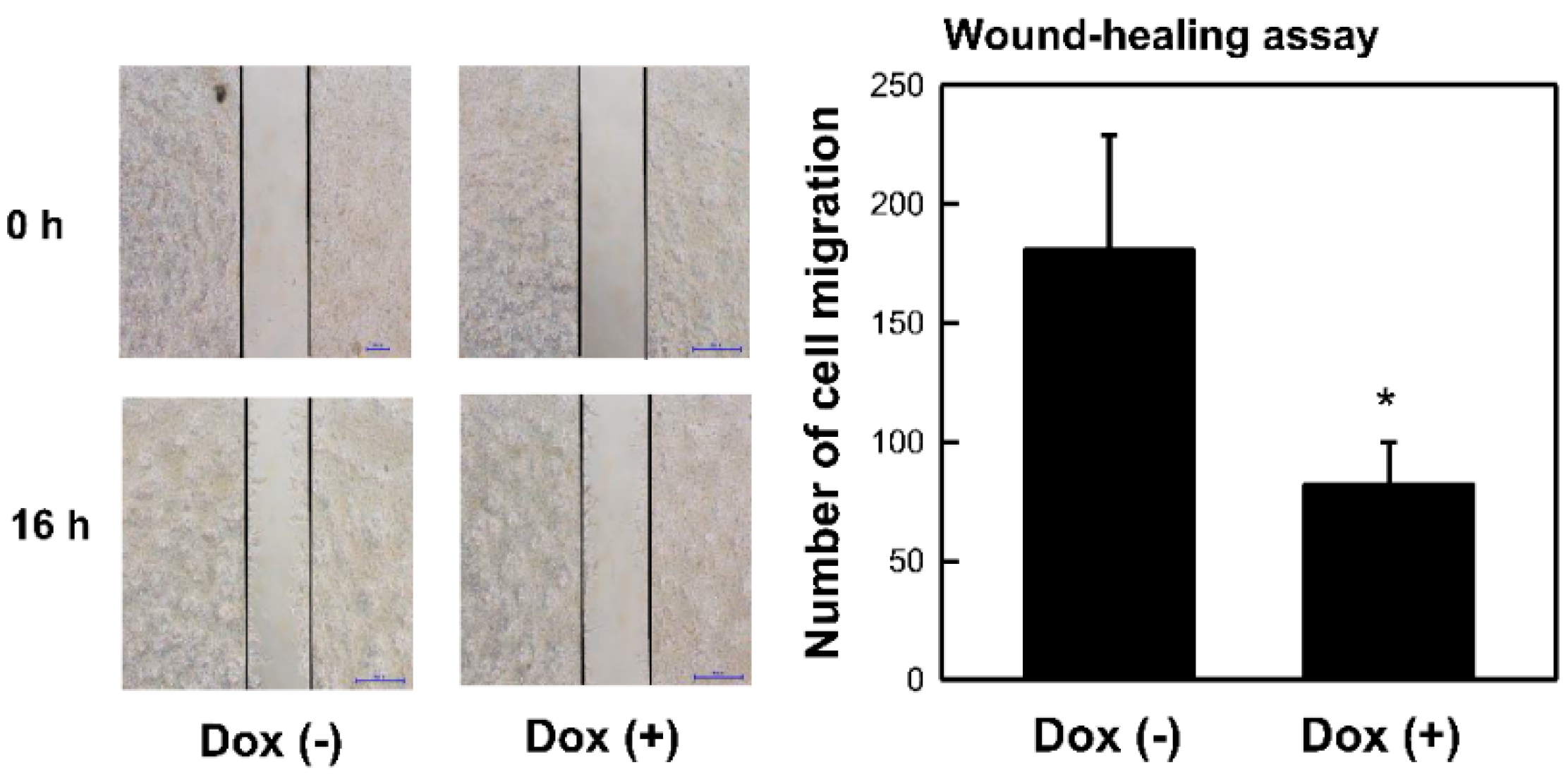
2.2. Overexpression of MCPIP3 Inhibits Cell Migration in T-REx-293/HA-MCPIP3 Cells
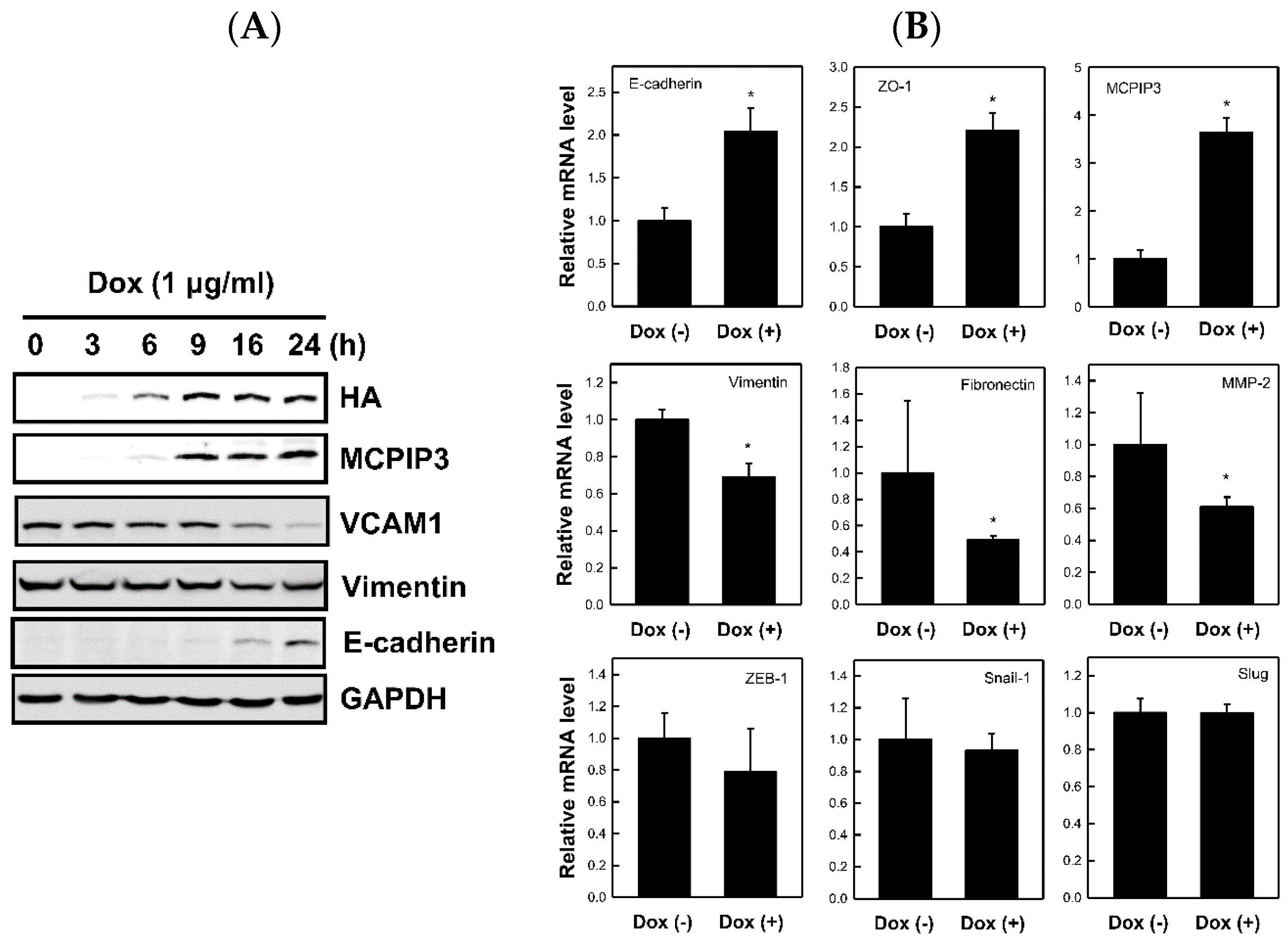
2.3. Overexpression of MCPIP3 Affects EMT-Related Protein Expressions in T-REx-293/HA-MCPIP3 Cells
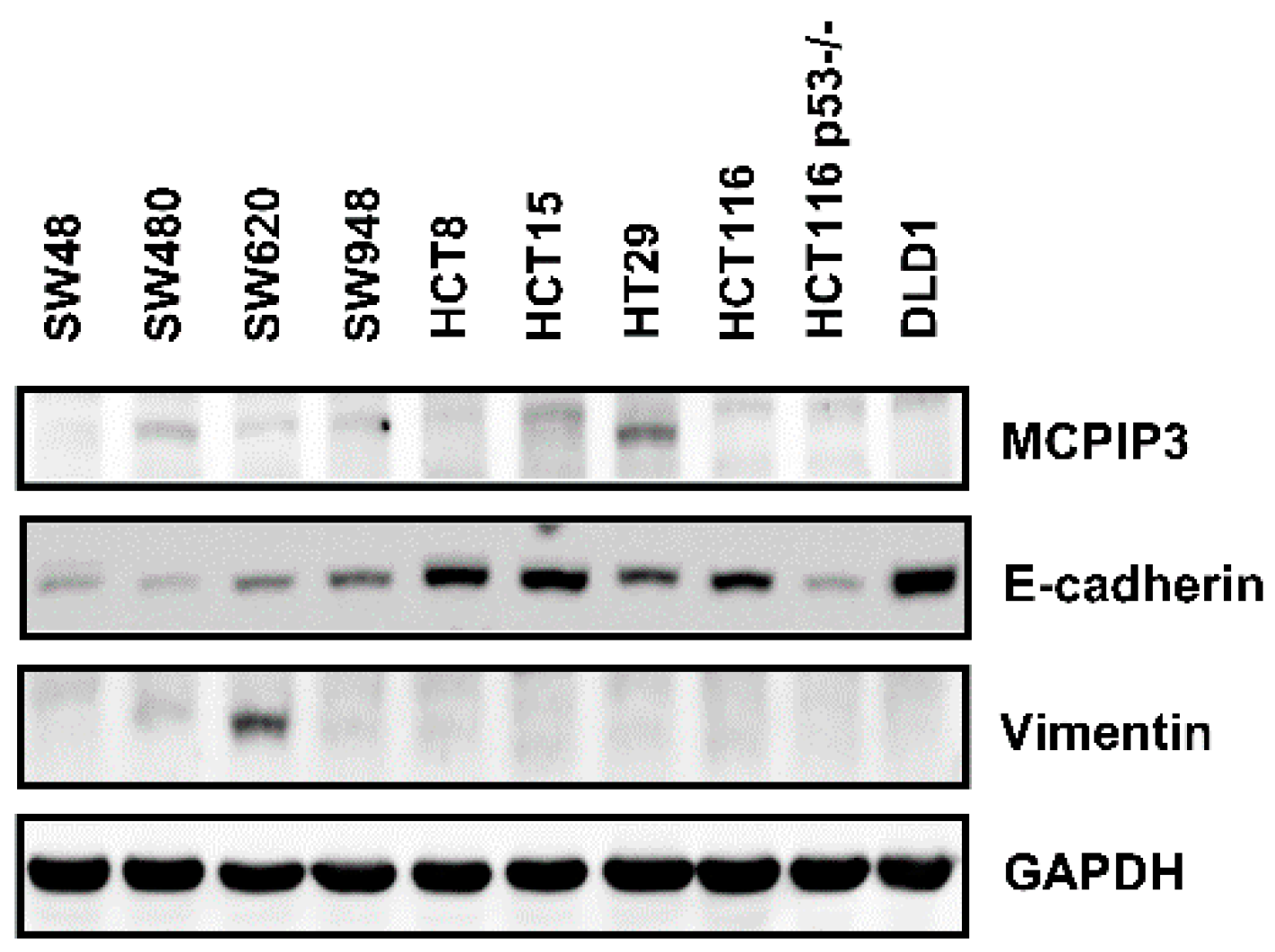
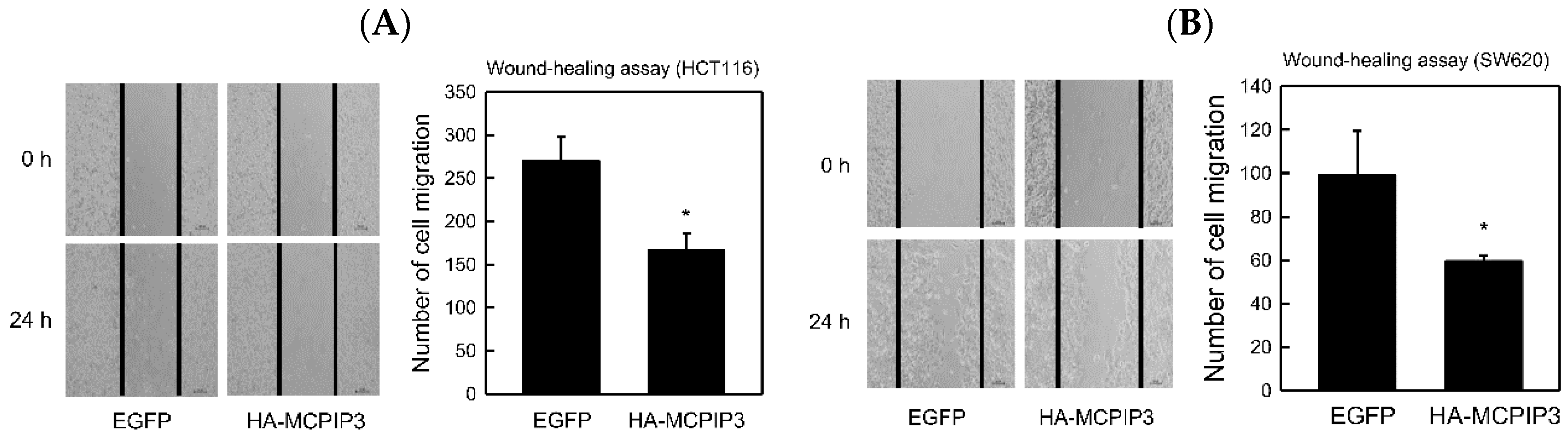
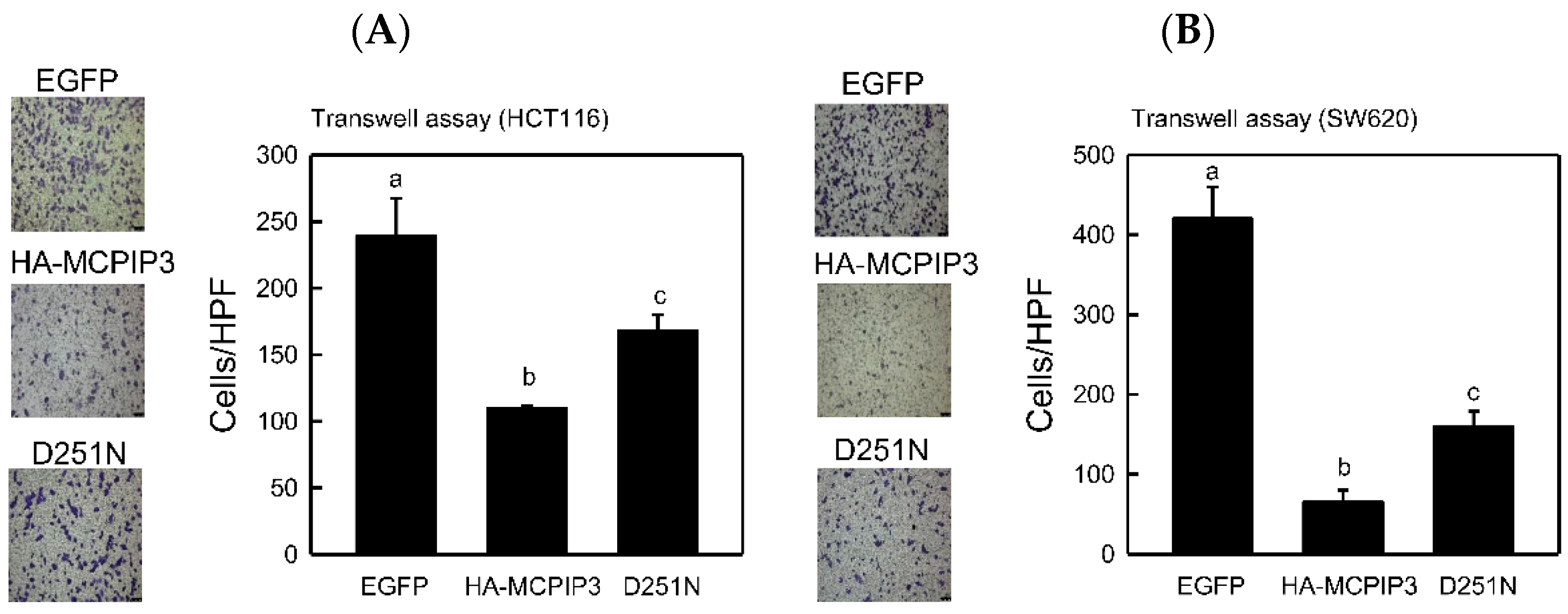
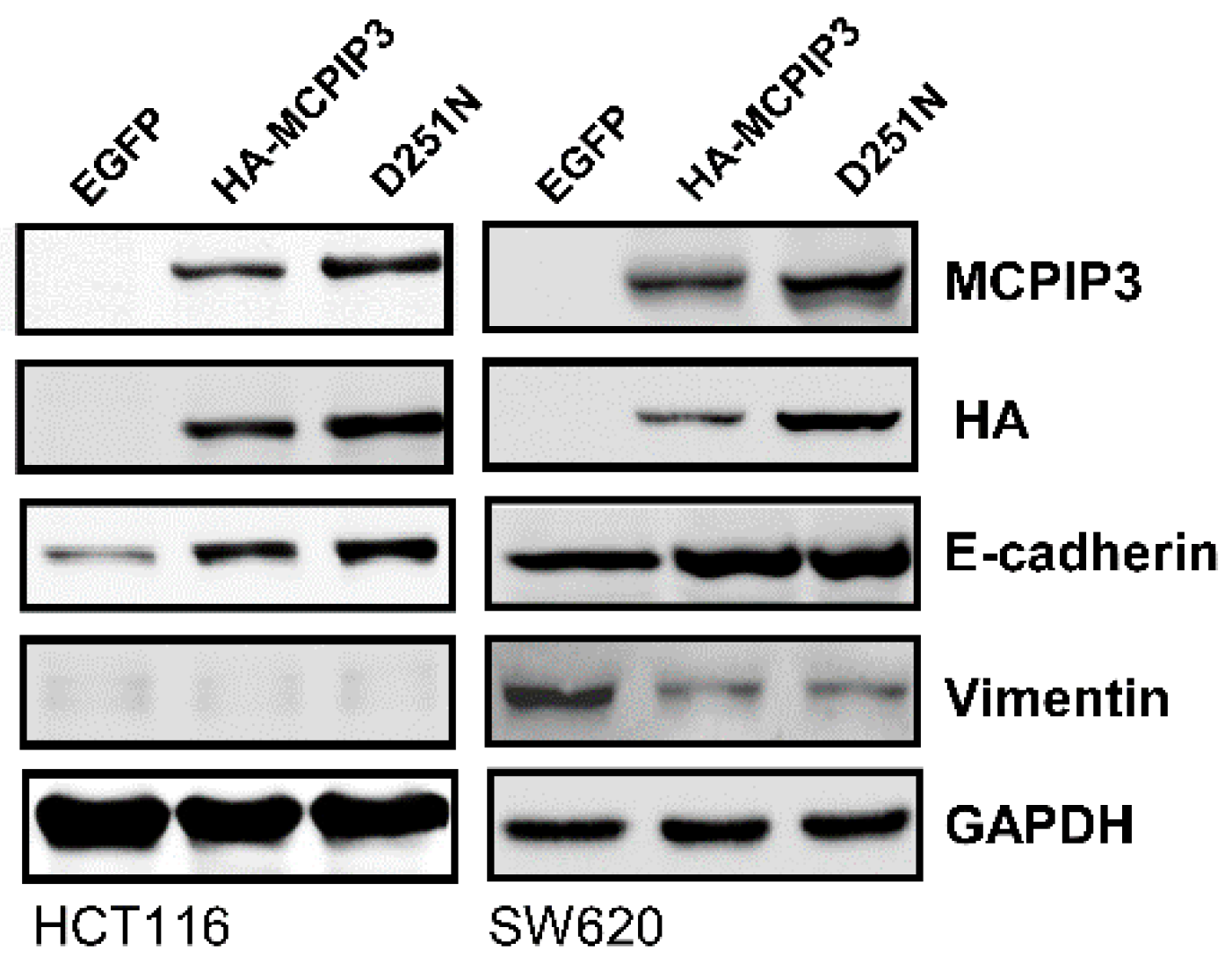
2.4. Overexpression of MCPIP3 Inhibits Cell Migration and Affects EMT-Related Gene Expressions in Human CRC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Antibodies, and Patient Samples
4.2. Plasmids and Lentivirus
4.3. Western Blotting
4.4. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.5. Cell Viability Assay
4.6. Wound-Healing Assay
4.7. Invasion Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, P.; Jenab, M.; Norat, T.; Moskal, A.; Slimani, N.; Olsen, A.; Tjønneland, A.; Overvad, K.; Jensen, M.K.; Boutron-Ruault, M.C.; et al. Lifetime and baseline alcohol intake and risk of colon and rectal cancers in the European prospective investigation into cancer and nutrition (EPIC). Int. J. Cancer 2007, 121, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Gomez, S.J.; Maziveyi, M.; Alahari, S.K. Regulation of epithelial-mesenchymal transition through epigenetic and post-translational modifications. Mol. Cancer 2016, 15, 18. [Google Scholar] [CrossRef] [PubMed]
- Samatov, T.R.; Tonevitsky, A.G.; Schumacher, U. Epithelial-mesenchymal transition: Focus on metastatic cascade, alternative splicing, non-coding RNAs and modulating compounds. Mol. Cancer 2013, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Dedhar, S.; Kalluri, R.; Thompson, E.W. The epithelial-mesenchymal transition: New insights in signaling, development, and disease. J. Cell Biol. 2006, 172, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Wang, J.; Azfer, A.; Song, W.; Tromp, G.; Kolattukudy, P.E.; Fu, M. A novel CCCH-zinc finger protein family regulates proinflammatory activation of macrophages. J. Biol. Chem. 2008, 283, 6337–6346. [Google Scholar] [CrossRef] [PubMed]
- Uehata, T.; Akira, S. mRNA degradation by the endoribonuclease Regnase-1/ZC3H12a/MCPIP-1. Biochim. Biophys. Acta 2013, 1829, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Takeuchi, O.; Standley, D.M.; Kumagai, Y.; Kawagoe, T.; Miyake, T.; Satoh, T.; Kato, H.; Tsujimura, T.; Nakamura, H.; et al. Zc3h12a is an RNase essential for controlling immune responses by regulating mRNA decay. Nature 2009, 458, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Mizgalska, D.; Wegrzyn, P.; Murzyn, K.; Kasza, A.; Koj, A.; Jura, J.; Jarzab, B.; Jura, J. Interleukin-1-inducible MCPIP protein has structural and functional properties of RNase and participates in degradation of IL-1β mRNA. FEBS J. 2009, 276, 7386–7399. [Google Scholar] [CrossRef] [PubMed]
- Skalniak, L.; Mizgalska, D.; Zarebski, A.; Wyrzykowska, P.; Koj, A.; Jura, J. Regulatory feedback loop between NF-κB and MCP-1-induced protein 1 RNase. FEBS J. 2009, 276, 5892–5905. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, K.; Katayama, Y.; Nishikawa, S.; Yamamoto, K.; Sada, A.; Okamura, A.; Shimoyama, M.; Matsui, T. Inhibition of G1 to S phase progression by a novel zinc finger protein P58(TFL) at P-bodies. Mol. Cancer Res. 2009, 7, 880–889. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Vikis, H.G.; Wang, Y.; Jia, D.; Wang, D.; Bierut, L.J.; Bailey-Wilson, J.E.; Amos, C.I.; Pinney, S.M.; Petersen, G.M.; et al. Identification of a novel tumor suppressor gene p34 on human chromosome 6q25.1. Cancer Res. 2007, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Okamura, A.; Minagawa, K.; Yakushijin, K.; Urahama, N.; Gomyo, H.; Shimoyama, M.; Itoh, M.; Matsui, T. A novel t(2;6)(p12;q23) appearing during transformation of follicular lymphoma with t(18;22)(q21;q11) to diffuse large cell lymphoma. Cancer Genet. Cytogenet. 2003, 147, 128–133. [Google Scholar] [CrossRef]
- Chan, C.H.; Jo, U.; Kohrman, A.; Rezaeian, A.H.; Chou, P.C.; Logothetis, C.; Lin, H.K. Posttranslational regulation of Akt in human cancer. Cell Biosci. 2014, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Bhoj, V.G.; Chen, Z.J. Ubiquitylation in innate and adaptive immunity. Nature 2009, 458, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Huang, X.; Xin, H.B.; Fu, M.; Xue, A.; Wu, Z.H. TRAF Family Member-associated NF-κB Activator (TANK) Inhibits Genotoxic Nuclear Factor κB Activation by Facilitating Deubiquitinase USP10-dependent Deubiquitination of TRAF6 Ligase. J. Biol. Chem. 2015, 290, 13372–13385. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Shi, Y.; Xue, J.; Miao, R.; Huang, S.; Wang, T.; Wu, J.; Fu, M.; Wu, Z.H. USP10 inhibits genotoxic NF-κB activation by MCPIP1-facilitated deubiquitination of NEMO. EMBO J. 2013, 32, 3206–3219. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Qi, D.; Liang, J.; Miao, R.; Minagawa, K.; Quinn, T.; Matsui, T.; Fan, D.; Liu, J.; Fu, M. The putative tumor suppressor Zc3h12d modulates toll-like receptor signaling in macrophages. Cell. Signal. 2012, 24, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhou, Z.; Huang, S.; Guo, Y.; Fan, Y.; Zhang, J.; Zhang, J.; Fu, M.; Chen, Y.E. Zc3h12c inhibits vascular inflammation by repressing NF-κB activation and pro-inflammatory gene expression in endothelial cells. Biochem. J. 2013, 451, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Kang, D.H.; Choi, M.; Choi, Y.J.; Lee, J.Y.; Park, J.H.; Park, Y.J.; Lee, K.W.; Kang, S.W. Peroxiredoxin-2 represses melanoma metastasis by increasing E-Cadherin/β-Catenin complexes in adherens junctions. Cancer Res. 2013, 73, 4744–4757. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Cao, W.; Liu, H.; Zhang, W.; Liu, X.; Cai, Z.; Guo, J.; Wang, X.; Hui, Z.; Zhang, H.; et al. MCPIP1 down-regulates IL-2 expression through an ARE-independent pathway. PLoS ONE 2012, 7, e49841. [Google Scholar] [CrossRef] [PubMed]
- Zehner, Z.E.; Shepherd, R.K.; Gabryszuk, J.; Fu, T.F.; Al-Ali, M.; Holmes, W.M. RNA-protein interactions within the 3′ untranslated region of vimentin mRNA. Nucleic Acids Res. 1997, 25, 3362–3370. [Google Scholar] [CrossRef] [PubMed]
- Giese, G.; Kubbies, M.; Traub, P. Cell cycle-dependent vimentin expression in elutriator-synchronized, TPA-treated MPC-11 mouse plasmacytoma cells. Exp. Cell Res. 1992, 200, 118–125. [Google Scholar] [CrossRef]
- Rittling, S.R.; Coutinho, L.; Amram, T.; Kolbe, M. AP-1/jun binding sites mediate serum inducibility of the human vimentin promoter. Nucleic Acids Res. 1989, 17, 1619–1633. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Saad, Y.; Lei, T.; Wang, J.; Qi, D.; Yang, Q.; Kolattukudy, P.E.; Fu, M. MCP-induced protein 1 deubiquitinates TRAF proteins and negatively regulates JNK and NF-κB signaling. J. Exp. Med. 2010, 207, 2959–2973. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Moon, I.S.; Choi, B.K.; Paik, J.W.; Kim, Y.S.; Choi, S.H.; Kim, C.K. Effects of sub-antimicrobial dose doxycycline therapy on crevicular fluid MMP-8, and gingival tissue MMP-9, TIMP-1 and IL-6 levels in chronic periodontitis. J. Periodontal Res. 2004, 39, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.C.; Weng, C.C.; Hou, Y.S.; Jian, S.F.; Fang, K.T.; Hou, M.F.; Cheng, K.H. Activation of VCAM-1 and its associated molecule CD44 leads to increased malignant potential of breast cancer cells. Int. J. Mol. Sci. 2014, 15, 3560–3579. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.J.; Chu, J.S.; Chien, H.L.; Tseng, C.H.; Ko, P.C.; Mei, Y.Y.; Tang, W.C.; Kao, Y.T.; Cheng, H.Y.; Liang, Y.C.; et al. MCPIP1 suppresses hepatitis C virus replication and negatively regulates virus-induced proinflammatory cytokine responses. J. Immunol. 2014, 193, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Suk, F.M.; Lin, S.Y.; Lin, R.J.; Hsine, Y.H.; Liao, Y.J.; Fang, S.U.; Liang, Y.C. Bortezomib inhibits Burkitt’s lymphoma cell proliferation by downregulating sumoylated hnRNP K and c-Myc expression. Oncotarget 2015, 6, 25988–26001. [Google Scholar] [CrossRef] [PubMed]
- Chin, L.H.; Hsu, S.P.; Zhong, W.B.; Liang, Y.C. Involvement of cysteine-rich protein 61 in the epidermal growth factor-induced migration of human anaplastic thyroid cancer cells. Mol. Carcinog. 2016, 55, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Wu, H.H.; Chen, T.H.; Leung, W.; Liang, Y.C. 15,16-Dihydrotanshinone I from the functional food salvia miltiorrhiza exhibits anticancer activity in human HL-60 leukemia cells: in vitro and in vivo Studies. Int. J. Mol. Sci. 2015, 16, 19387–19400. [Google Scholar] [CrossRef] [PubMed]
- Chin, L.H.; Hsu, S.P.; Zhong, W.B.; Liang, Y.C. Combined treatment with troglitazone and lovastatin inhibited epidermal growth factor-induced migration through the downregulation of cysteine-rich protein 61 in human anaplastic thyroid cancer cells. PLoS ONE 2015, 10, e0118674. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suk, F.-M.; Chang, C.-C.; Lin, R.-J.; Lin, S.-Y.; Chen, Y.-T.; Liang, Y.-C. MCPIP3 as a Potential Metastasis Suppressor Gene in Human Colorectal Cancer. Int. J. Mol. Sci. 2018, 19, 1350. https://doi.org/10.3390/ijms19051350
Suk F-M, Chang C-C, Lin R-J, Lin S-Y, Chen Y-T, Liang Y-C. MCPIP3 as a Potential Metastasis Suppressor Gene in Human Colorectal Cancer. International Journal of Molecular Sciences. 2018; 19(5):1350. https://doi.org/10.3390/ijms19051350
Chicago/Turabian StyleSuk, Fat-Moon, Chi-Ching Chang, Ren-Jye Lin, Shyr-Yi Lin, Ya-Ting Chen, and Yu-Chih Liang. 2018. "MCPIP3 as a Potential Metastasis Suppressor Gene in Human Colorectal Cancer" International Journal of Molecular Sciences 19, no. 5: 1350. https://doi.org/10.3390/ijms19051350