The Role of Peroxisome Proliferator-Activated Receptors and Their Transcriptional Coactivators Gene Variations in Human Trainability: A Systematic Review


 ,
,  , and
, and
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
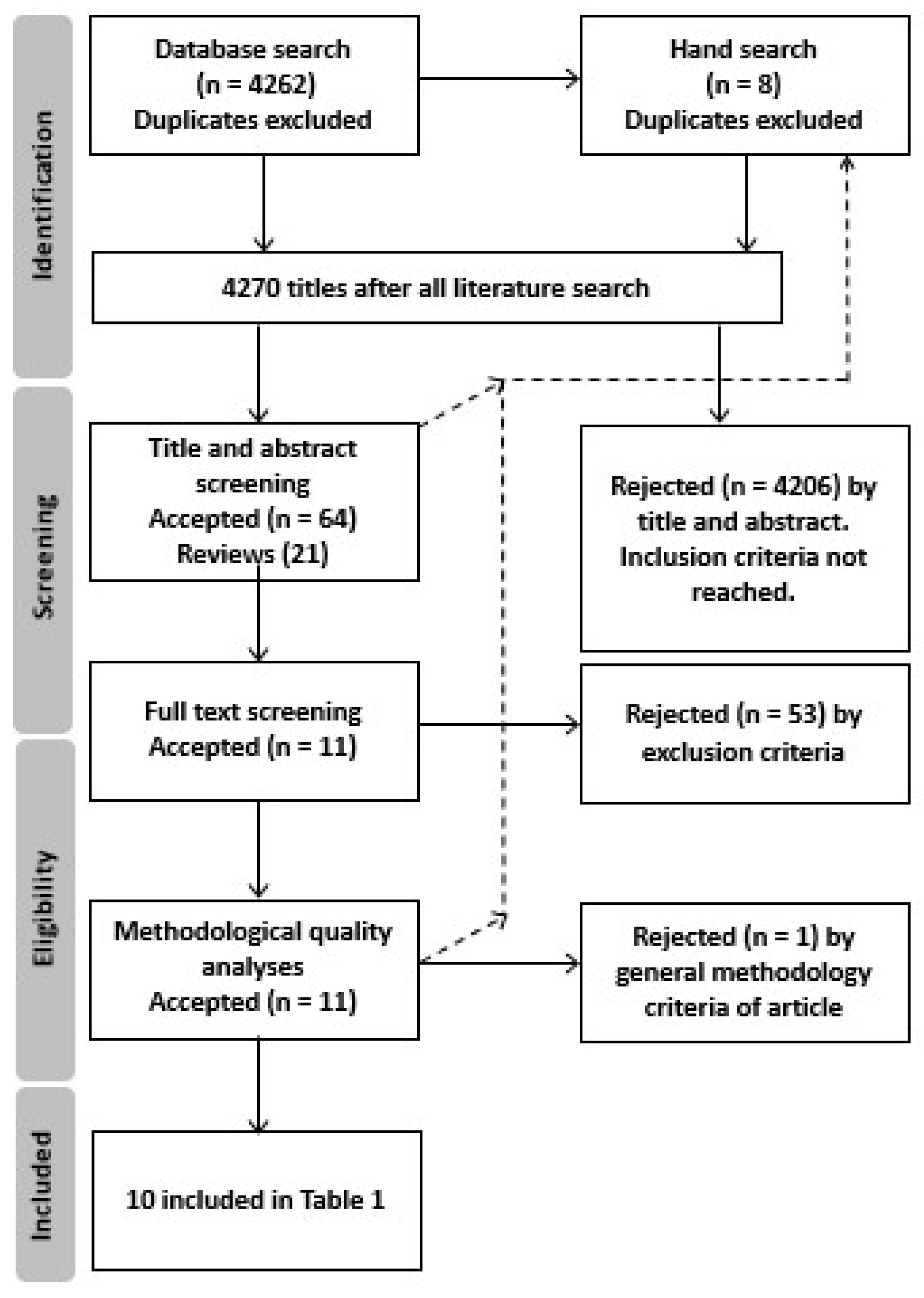
4.1. Review Process
4.2. Literature Search
4.3. Literature Selection
- Sampling of genetic polymorphisms in the PPARA, PPARG, PPARD, PPARGC1A and PPARGC1B, genes.
- Analyses of genetic polymorphisms on sport phenotype (markers of sport phenotype) or related physical activity domains (e.g., body mass, fat mass, energy uptake, performance, physical fitness).
- Population of athletes and other healthy populations with a physical activity record.
- Cross-sectional, cohort, case control, intervention, control trials or GWAS.
- (1)
- the full text was not available in English;
- (2)
- the study did not contain an appropriate description of measuring devices, physical activity or genetic sampling procedures;
- (3)
- the study did not include a specification of physical activity;
- (4)
- the study did not report a quantitative performance outcome;
- (5)
- the study did not perform the intervention of a physical training program;
- (6)
- the study was not reproducible by the methodological quality criteria.
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| GWAS | Genome-wide association study |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta Analyses |
| SPIRIT | Standard Protocol Item Recommendation for Interventional Trials |
| PPAR | Peroxisome proliferator-activated receptor |
| IRI | Fasting immunoreactive insulin |
| HOMA-R | Homeostasis model assessment-insulin resistance index |
References
- Bouchard, C.; Rankinen, T. Individual differences in response to regular physical activity. Med. Sci. Sports Exerc. 2001, 33, S446–S451; discussion S52–S53. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Blair, S.N.; Church, T.S.; Earnest, C.P.; Hagberg, J.M.; Hakkinen, K.; Jenkins, N.T.; Karavirta, L.; Kraus, W.E.; Leon, A.S.; et al. Adverse metabolic response to regular exercise: Is it a rare or common occurrence? PLoS ONE 2012, 7, e37887. [Google Scholar] [CrossRef] [PubMed]
- Dubuquoy, L.; Dharancy, S.; Nutten, S.; Pettersson, S.; Auwerx, J.; Desreumaux, P. Role of peroxisome proliferator-activated receptor γ and retinoid X receptor heterodimer in hepatogastroenterological diseases. Lancet 2002, 360, 1410–1418. [Google Scholar] [CrossRef]
- Cabrero, A.; Laguna, J.; Vazquez, M. Peroxisome proliferator-activated receptors and the control of inflammation. Curr. Drug Targets-Inflamm. Allergy 2002, 1, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.W.; Christophi, C.A.; Jablonski, K.A.; Billings, L.K.; Delahanty, L.M.; Horton, E.S.; Knowler, W.C.; Florez, J.C.; Diabetes Prevention Program Research Group. Common variation at PPARGC1A/B and change in body composition and metabolic traits following preventive interventions: The Diabetes Prevention Program. Diabetologia 2014, 57, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Williams, A.G.; Popov, D.V.; Lyubaeva, E.V.; Hakimullina, A.M.; Fedotovskaya, O.N.; Mozhayskaya, I.A.; Vinogradova, O.L.; Astratenkova, I.V.; Montgomery, H.E.; et al. The combined impact of metabolic gene polymorphisms on elite endurance athlete status and related phenotypes. Hum. Genet. 2009, 126, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Popov, D.V.; Mozhaiskaia, I.A.; Missina, S.S.; Astratenkova, I.V.; Vinogradova, O.L.; Rogozkin, V.A. Association of regulatory genes polymorphisms with aerobic and anaerobic performance of athletes. Rossiskii Fiziologicheski Zhurnal Imeni IM Sechenova/Rossiskaia Akademiia Nauk 2007, 93, 837–843. [Google Scholar]
- Franks, P.W.; Barroso, I.; Luan, J.; Ekelund, U.; Crowley, V.E.F.; Brage, S.; Sandhu, M.S.; Jakes, R.; Middelberg, R.P.S.; Harding, A.-H.; et al. PGC-1α Genotype Modifies the Association of Volitional Energy Expenditure with VO2 max. Med. Sci. Sports Exerc. 2003, 35, 1998–2004. [Google Scholar] [CrossRef] [PubMed]
- Petr, M.; Št’Astný, P.; Pecha, O.; Šteffl, M.; Šeda, O.; Kohlíková, E. PPARA intron polymorphism associated with power performance in 30-s anaerobic wingate test. PLoS ONE 2014, 9, e107171. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Vivar, J.C.; Sarzynski, M.A.; Sung, Y.J.; Timmons, J.A.; Bouchard, C.; Rankinen, T. Integrative pathway analysis of a genome-wide association study of VO2 max response to exercise training. J. Appl. Physiol. 2013, 115, 1343–1359. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Y.; Wang, Y.B.; Shen, X.Z.; Cai, G. Association of elite athlete performance and gene polymorphisms. Chin. J. Tissue Eng. Res. 2014, 18, 1121–1128. [Google Scholar]
- Lopez-Leon, S.; Tuvblad, C.; Forero, D.A. Sports genetics: The PPARA gene and athletes’ high ability in endurance sports. A systematic review and meta-analysis. Biol. Sport 2016, 33, 3–6. [Google Scholar] [PubMed]
- Ahmetov, I.I.; Fedotovskaya, O.N. Current Progress in Sports Genomics; Advances in Clinical Chemistry; Academic Press Inc.: New York, NY, USA, 2015; pp. 247–314. [Google Scholar]
- Leońska-Duniec, A.; Ahmetov, I.I.; Zmijewski, P. Genetic variants influencing effectiveness of exercise training programmes in obesity—An overview of human studies. Biol. Sport 2016, 33, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Aksenov, M.O.; Ilyin, A.B. Training process design in weightlifting sports customized to genetic predispositions. Teoriya Praktika Fizicheskoy Kultury 2017, 2017, 75–77. [Google Scholar]
- He, Z.; Hu, Y.; Feng, L.; Bao, D.; Wang, L.; Li, Y.; Wang, J.; Liu, G.; Xi, Y.; Wen, L.; et al. Is there an association between PPARGC1A genotypes and endurance capacity in Chinese men? Scand. J. Med. Sci. Sports 2008, 18, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Thamer, C.; Staiger, H.; Machicao, F.; Machann, J.; Schick, F.; Venter, C.; Niess, A.; Laakso, M.; Fritsche, A.; et al. Genetic variations in PPARD and PPARGC1A determine mitochondrial function and change in aerobic physical fitness and insulin sensitivity during lifestyle intervention. J. Clin. Endocrinol. Metab. 2007, 92, 1827–1833. [Google Scholar] [CrossRef] [PubMed]
- Steinbacher, P.; Feichtinger, R.G.; Kedenko, L.; Kedenko, I.; Reinhardt, S.; Schönauer, A.L.; Leitner, I.; Sänger, A.M.; Stoiber, W.; Kofler, B.; et al. The single nucleotide polymorphism Gly482Ser in the PGC-1α gene impairs exercise-induced slow-twitch muscle fibre transformation in humans. PLoS ONE 2015, 10, e0123881. [Google Scholar] [CrossRef] [PubMed]
- Tobina, T.; Mori, Y.; Doi, Y.; Nakayama, F.; Kiyonaga, A.; Tanaka, H. Peroxisome proliferator-activated receptor gamma co-activator 1 gene Gly482Ser polymorphism is associated with the response of low-density lipoprotein cholesterol concentrations to exercise training in elderly Japanese. J. Physiol. Sci. 2017, 67, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Ring-Dimitriou, S.; Kedenko, L.; Kedenko, I.; Feichtinger, R.G.; Steinbacher, P.; Stoiber, W.; Förster, H.; Felder, T.K.; Müller, E.; Kofler, B.; et al. Does genetic variation in PPARGC1A affect exercise-induced changes in ventilatory thresholds and metabolic syndrome? J. Exerc. Physiol. Online 2014, 17, 1–18. [Google Scholar]
- Hautala, A.J.; Leon, A.S.; Skinner, J.S.; Rao, D.C.; Bouchard, C.; Rankinen, T. Peroxisome proliferator-activated receptor-delta polymorphisms are associated with physical performance and plasma lipids: The HERITAGE Family Study. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2498–H2505. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.P.; Kulaputana, O.; Ghiu, I.A.; Brandauer, J.; Wohn, C.R.; Phares, D.A.; Shuldiner, A.R.; Hagberg, J.M. Endurance training-induced changes in the insulin response to oral glucose are associated with the peroxisome proliferator-activated receptor-γ2 Pro12Ala genotype in men but not in women. Metabolism 2005, 54, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Zarebska, A.; Jastrzebski, Z.; Cieszczyk, P.; Leonska-Duniec, A.; Kotarska, K.; Kaczmarczyk, M.; Sawczuk, M.; Maciejewska-Karlowska, A. The Pro12Ala polymorphism of the peroxisome proliferator-activated receptor gamma gene modifies the association of physical activity and body mass changes in Polish women. PPAR Res. 2014, 2014, 373782. [Google Scholar] [CrossRef] [PubMed]
- Pérusse, L.; Ruchat, S.M.; Rankinen, T.; Weisnagel, S.J.; Rice, T.; Rao, D.C.; Bergman, R.N.; Bouchard, C.; Pérusse, L. Improvements in glucose homeostasis in response to regular exercise are influenced by the PPARG Pro12Ala variant: Results from the HERITAGE family study. Diabetologia 2010, 53, 679–689. [Google Scholar]
- Kahara, T.; Takamura, T.; Hayakawa, T.; Nagai, Y.; Yamaguchi, H.; Katsuki, T.; Katsuki, K.; Katsuki, M.; Kobayashi, K. PPARγ gene polymorphism is associated with exercise-mediated changes of insulin resistance in healthy men. Metabolism 2003, 52, 209–212. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, Y.; Hu, Y.; Ren, Z.; Yi, L. Delta-aminolevulinate synthase 2 polymorphism is associated with maximal oxygen uptake after living-high exercise-high training-low in a male chinese population. Int. J. Clin. Exp. Med. 2015, 8, 21617. [Google Scholar] [PubMed]
- Clarkson, P.M.; Devaney, J.M.; Gordish-Dressman, H.; Thompson, P.D.; Hubal, M.J.; Urso, M.; Price, T.B.; Angelopoulos, T.J.; Gordon, P.M.; Moyna, N.M.; et al. ACTN3 genotype is associated with increases in muscle strength in response to resistance training in women. J. Appl. Physiol. 2005, 99, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, A.; Sawczuk, M.; Cieszczyk, P.; Mozhayskaya, I.A.; Ahmetov, I. The PPARGC1A gene Gly482Ser in Polish and Russian athletes. J. Sports Sci. 2012, 30, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-L.; Lu, W.-S.; Yan, L.; Wu, M.-C.; Xu, M.-T.; Chen, L.-H.; Cheng, H. Association between peroxisome proliferator-activated receptor-gamma coactivator-1alpha gene polymorphisms and type 2 diabetes in southern Chinese population: Role of altered interaction with myocyte enhancer factor 2C. Chin. Med. J. 2007, 120, 1878–1885. [Google Scholar] [PubMed]
- Michael, L.F.; Wu, Z.; Cheatham, R.B.; Puigserver, P.; Adelmant, G.; Lehman, J.J.; Kelly, D.P.; Spiegelman, B.M. Restoration of insulin-sensitive glucose transporter (GLUT4) gene expression in muscle cells by the transcriptional coactivator PGC-1. Proc. Natl. Acad. Sci. USA 2001, 98, 3820–3825. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.-Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator PGC-1α drives the formation of slow-twitch muscle fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Skogsberg, J.; Kannisto, K.; Roshani, L.; Gagne, E.; Hamsten, A.; Larsson, C.; Ehrenborg, E. Characterization of the human peroxisome proliferator activated receptor delta gene and its expression. Int. J. Mol. Med. 2000, 6, 73–154. [Google Scholar] [CrossRef] [PubMed]
- Skogsberg, J.; Kannisto, K.; Cassel, T.N.; Hamsten, A.; Eriksson, P.; Ehrenborg, E. Evidence that peroxisome proliferator–activated receptor delta influences cholesterol metabolism in men. Arterioscler. Thromb. Vas. Biol. 2003, 23, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Karpe, F.; Ehrenborg, E.E. PPARδ in humans: Genetic and pharmacological evidence for a significant metabolic function. Curr. Opin. Lipidol. 2009, 20, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, G.; Rivellese, A. Dietary treatment of the metabolic syndrome—The optimal diet. Br. J. Nutr. 2000, 83, S143–S148. [Google Scholar] [CrossRef] [PubMed]
- Nathan, D.M.; Davidson, M.B.; DeFronzo, R.A.; Heine, R.J.; Henry, R.R.; Pratley, R.; Zinman, B.; American Diabetes Association. Impaired fasting glucose and impaired glucose tolerance: Implications for care. Diabetes Care 2007, 30, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.E.; Goodpaster, B.H. Effects of physical activity on insulin action and glucose tolerance in obesity. Med. Sci. Sports Exerc. 1999, 31 (Suppl. 11), S619–S623. [Google Scholar] [CrossRef] [PubMed]
- Adamo, K.; Sigal, R.; Williams, K.; Kenny, G.; Prud’homme, D.; Tesson, F. Influence of Pro12Ala peroxisome proliferator-activated receptor γ2 polymorphism on glucose response to exercise training in type 2 diabetes. Diabetologia 2005, 48, 1503–1509. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Deeb, S.S.; Fajas, L.; Nemoto, M.; Pihlajamäki, J.; Mykkänen, L.; Kuusisto, J.; Laakso, M.; Fujimoto, W.; Auwerx, J. A Pro12Ala substitution in PPARγ2 associated with decreased receptor activity, lower body mass index and improved insulin sensitivity. Nat. Genet. 1998, 20, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Razquin, C.; Martinez, J.A.; Martinez-Gonzalez, M.A.; Corella, D.; Santos, J.M.; Marti, A. The Mediterranean diet protects against waist circumference enlargement in 12Ala carriers for the PPARγ gene: 2 years’ follow-up of 774 subjects at high cardiovascular risk. Br. J. Nutr. 2009, 102, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.-J.; Beamer, B.A.; Negri, C.; Silver, K.; Brown, K.A.; Yarnall, D.P.; Burns, D.K.; Roth, J.; Shuldiner, A.R. Molecular scanning of the human peroxisome proliferator activated receptor γ (hPPARγ) gene in diabetic Caucasians: Identification of a Pro12Ala PPARγ2 missense mutation. Biochem. Biophys. Res. Commun. 1997, 241, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Masugi, J.; Tamori, Y.; Mori, H.; Koike, T.; Kasuga, M. Inhibitory effect of a proline-to-alanine substitution at codon 12 of peroxisome proliferator-activated receptor-γ2 on thiazolidinedione-induced adipogenesis. Biochem. Biophys. Res. Commun. 2000, 268, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Kreuzer, J.; Hamann, A.; Nawroth, P.P.; Dugi, K.A. The proline 12 alanine substitution in the peroxisome proliferator–Activated receptor-γ2 gene is associated with lower lipoprotein lipase activity in vivo. Diabetes 2002, 51, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Ek, J.; Andersen, G.; Urhammer, S.; Hansen, L.; Carstensen, B.; Borch-Johnsen, K.; Drivsholm, T.; Berglund, L.; Hansen, T.; Lithell, H.; et al. Studies of the Pro12Ala polymorphism of the peroxisome proliferator-activated receptor-γ2 (PPAR-γ2) gene in relation to insulin sensitivity among glucose tolerant Caucasians. Diabetologia 2001, 44, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- Honka, M.-J.; Vänttinen, M.; Iozzo, P.; Virtanen, K.A.; Lautamäki, R.; Hällsten, K.; Borraa, R.J.H.; Takalaa, T.; Viljanena, A.P.M.; Kemppainen, J.; et al. The Pro12Ala polymorphism of the PPARγ2 gene is associated with hepatic glucose uptake during hyperinsulinemia in subjects with type 2 diabetes mellitus. Metabolism 2009, 58, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wu, Y.; Qi, Q.; Liu, C.; Gan, W.; Zhu, J.; Li, H.; Lin, X. Associations of type 2 diabetes with common variants in PPARD and the modifying effect of vitamin D among middle-aged and elderly Chinese. PLoS ONE 2012, 7, e34895. [Google Scholar] [CrossRef] [PubMed]
- Thamer, C.; Machann, J.; Stefan, N.; Schäfer, S.A.; Machicao, F.; Staiger, H.; Laakso, M.; Böttcher, M.; Claussen, C.; Schick, F.; et al. Variations in PPARD determine the change in body composition during lifestyle intervention: A whole-body magnetic resonance study. J. Clin. Endocrinol. Metab. 2008, 93, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Masschelein, E.; Puype, J.; Broos, S.; Van Thienen, R.; Deldicque, L.; Lambrechts, D.; Hespel, P.; Thomis, M. A genetic predisposition score associates with reduced aerobic capacity in response to acute normobaric hypoxia in lowlanders. High Alt. Med. Biol. 2015, 16, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Schulz, K.F.; Simera, I.; Altman, D.G. Guidance for developers of health research reporting guidelines. PLoS Med. 2010, 7, e1000217. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Study | Gene/Polymorphism | Population | Aim | Main Result |
|---|---|---|---|---|
| Stefan et al., 2007 [17] | PPARGC1A Gly482Ser (rs8192678) PPARD (rs2267668) (rs6902123) (rs2076167) (rs1053049) | German; n = 136 (men 63, women 73), Tuebingen Lifestyle Intervention Program. Age 45 ± 1 years, body mass 86.5 ± 1.5 kg | To investigate, whether selected SNPs predict the response of aerobic exercise training on changes in aerobic physical fitness and insulin sensitivity and whether they affect mitochondrial function in human myotubes in vitro. | Genetic variations in PPARD and PPARGC1A modulate mitochondrial function and changes in aerobic physical fitness and insulin sensitivity during lifestyle intervention. |
| Steinbacher et al., 2015 [18] | PPARGC1A Gly482Ser (rs8192678) | Austrian; n = 28 (men only), Salzburg Atherosclerosis Prevention Programme in Subjects at High Individual Risk. Age 59 ± 7 years (range 50–69), body mass 88 ± 2.2 kg | To investigate the myocellular responses in the vastus lateralis muscle of untrained male carriers of this SNP and of a control group after 10 weeks of endurance training. | The single nucleotide polymorphism Gly482Ser in the PPARGC1A gene impairs exercise-induced slow-twitch muscle fiber transformation in humans. |
| Tobina et al., 2017 [19] | PPARGC1A Gly482Ser (rs8192678) | Japanese; n = 119 (men 49, women 70), all participants >65 years of age. Age 71 ± 6 years, body mass 57.5 ± 9.8 kg | This study investigated the effects of PPARGC1A Gly482Ser polymorphisms on alterations in glucose and lipid metabolism induced by 12 weeks of exercise training. | The PPARGC1A Gly482Ser polymorphism is associated with the response of low-density lipoprotein cholesterol concentrations following exercise training in elderly Japanese. |
| Ring-Dimitriou, et al., 2014 [20] | PPARGC1A Gly482Ser (rs8192678) | Austrian; n = 24 (men only), untrained individuals selected from SAPHIR program. Age 58.3 ± 5.7 years, body mass 87.2 ± 7.6 kg | To test if untrained men who are homozygous or heterozygous carriers of the rare allele in PPARGC1A show a reduced change in oxygen uptake and work rate at the submaximal performance level compared to men characterized by the common genotype after 10 weeks of endurance exercise. | Investigated SNP affects the trainability of aerobic capacity measured as VO2 or work rate at the respiratory compensation point of previously untrained middle-aged men. The highest responders were Gly/Gly genotypes compared to Gly/Ser and Ser/Ser genotypes. |
| He et al., 2008 [16] | PPARGC1A Thr394Thr (rs17847357) Gly482Ser (rs8192678) A2962G (rs6821591) | Chinese of Han origin; n = 102 (men only), soldiers from a local police army. Age 19 ± 1 years, height 171.7 ± 5.8 cm, body mass 60.3 ± 6.5 kg | To examine the possible association between PPARGC1A genotypes and both maximal (i.e., VO2max) and submaximal endurance capacity (i.e., running economy in a pre-training state (baseline) and after endurance training. | None of the VO2max and RE-related traits were associated with the Gly482Ser and Thr394Thr polymorphisms at baseline nor after training. The A2962G polymorphism was however associated with VO2max at baseline, as carriers of the G allele (AG1GG genotypes; n = 49) had higher levels of VO2max than the AA group (n = 53). |
| Weiss et al., 2005 [22] | PPARG Pro12Ala (rs1801282) | Caucasian; n = 73, (men 32, women 41), healthy sedentary subjects aged 50–75 years. | To investigate whether a common functional gene variant predicts insulin action and whether improvements in insulin action in response to endurance exercise training are associated with PPARG Pro12Ala. | Endurance training-induced changes in the insulin response to oral glucose are associated with the PPARG Pro12Ala genotype in men, but not in women. |
| Zarebska et al., 2014 [23] | PPARG Pro12Ala (rs1801282) | Polish; n = 201 (women only), no history of any metabolic or cardiovascular diseases. Age 21 ± 1 years | To examine the genotype distribution of the PPARG Pro12Ala allele in a group of Polish women measured for selected body mass and body composition variables before and after the completion of a 12-week training program. | The Pro12Ala polymorphism modifies the association of physical activity and body mass changes in Polish women. |
| Péruse et al., 2010 [24] | PPARG Pro12Ala (rs1801282) | White; n = 481 (men 233, women 248 from 98 nuclear families), sedentary non-diabetic subjects from the HERITAGE study who finished a 20-week endurance training program. Age 36 ± 0.67 years | To investigate whether variants either confirmed or newly identified as diabetes susceptibility variants through GWAS modulate changes in phenotypes derived from an intravenous glucose tolerance test (IVGTT) in response to an endurance training program. | Improvements in glucose homeostasis in response to regular exercise are influenced by the PPARG Pro12Ala variant. |
| Kahara et al., 2003 [25] | PPARG Pro12Ala | Japanese; n = 123, men, age 21–69 years. Age ± SD, 45.2 ± 11.6 years | To examine the association of PPARG gene polymorphism in Japanese healthy men with changes in insulin resistance after intervention with an exercise program. | The PPARG gene polymorphism may be a reliable indicator of whether exercise will have a beneficial effect as part of the treatment of insulin resistance syndrome. |
| Hautala et al., 2007 [21] | PPARD +15C/T (rs2016520) +65A/G (rs2076167) | American; n = 462 white subjects (223 males, 239 females) n = 256 black subjects (87 males, 169 females) from the HERITAGE study. Age 17–65 years | To test the hypothesis that PPARD gene polymorphisms are associated with cardiorespiratory fitness and plasma lipid responses to endurance training. | DNA sequence variation in the PPARD locus is a potential modifier of changes in cardiorespiratory fitness and plasma high-density lipoprotein cholesterol in healthy individuals in response to regular exercise. |
| Gene/Polymorphism | Intervention | Genotype/Allele Difference | Parameters | Study |
|---|---|---|---|---|
| PPARGC1A Gly482Ser rs8192678 | 9 months 3 h/w of moderate sports endurance exercise (e.g., walking, swimming) | Gly/Gly > Ser allele carriers | ↑ individual anaerobic threshold (W) | Stefan et al., 2007 [17] |
| 10 w 3/w 60 min cycling training at a heart rate equaling 70–90% of peak oxygen uptake (VO2peak) | Gly/Gly > Ser allele carriers (Ser allele-no response) | ↑ slow muscle fibers’ proportion | Steinbacher et al., 2015 [18] | |
| Gly/Gly > Ser allele carriers | ↑ mitochondria activity–Complex II | Steinbacher et al., 2015 [18] | ||
| 12 w 140 min/w 20 cm bench-stepping exercise training at lactate threshold intensity | Gly/Gly > Ser allele carriers (Ser allele-no response) | ↓ low-density and total lipoprotein cholesterol | Tobina et al., 2017 [19] | |
| 10 w 3/w 45–60 min HR equaling 80–100% of the anaerobic threshold (ANT) | Gly/Gly > Ser allele carriers (Ser/Ser-no response) | ↑ VO2peak (mL·min−1·kg) | Ring-Dimitriou, et al., 2014 [20] | |
| PPARD rs1053049 (complete LD with rs2076167) | 9 months 3 h/w of moderate sports endurance exercise (e.g., walking, swimming) | TT > C allele carriers | ↑ insulin sensitivity | Stefan et al., 2007 [17] |
| TT > C allele carriers | ↓ fasting insulin levels | Stefan et al., 2007 [17] | ||
| PPARD rs2267668 | 9 months 3 h/w of moderate sports endurance exercise (e.g., walking, swimming) | AA > G allele carriers | ↑ insulin sensitivity | Stefan et al., 2007 [17] |
| AA > G allele carriers | ↑ individual anaerobic threshold (W) | Stefan et al., 2007 [17] | ||
| AA > G allele carriers (G allele-negative response) | ↑ VO2peak (mL·min−1·kg lbm) | Stefan et al., 2007 [17] | ||
| PPARD +15C/T (rs2016520) +65A/G (rs2076167) | 20 w 3/w at 55–75% of baseline VO2max for 30–50 min | T allele carriers > +15CC (only in black subjects) | ↑ VO2max ↑ maximum power output | Hautala et al., 2007 [21] |
| +15CC > T allele carriers +65GG > A allele carriers (only in white subjects) | ↑ plasma HDL cholesterol | Hautala et al., 2007 [21] | ||
| PPARG Pro12Ala rs1801282 | 10 w 3–4/w 40 min sessions of endurance treadmill walking and stationary cycling at 65–75% of heart rate reserve | Men: Pro/Pro < Ala/Pro | ↓ fasting insulin and insulin AUC following intervention | Weiss et al., 2005 [22] |
| 12 w 3/w 60 min at 50–75% heart rate max. aerobic | Pro/Pro > Ala allele carriers | ↓ body fat | Zarebska et al., 2014 [23] | |
| 20 w 3/w at 55–75% of baseline VO2max for 30–50 min | Ala carriers > Pro/Pro (Pro allele-negative response in some parameters) | ↑ glucose tolerance (glucose effectiveness, acute insulin response to glucose, and disposition index) | Péruse et al., 2010 [24] | |
| 3 months 2–3/w 2–3/day 20–60 min at 50% of the maximal heart rate of brisk walking | Ala allele carriers > Pro/Pro | ↓ fasting immunoreactive insulin (IRI) ↓ homeostasis model assessment–insulin resistance index (HOMA-R) | Kahara et al., 2003 [25] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petr, M.; Stastny, P.; Zajac, A.; Tufano, J.J.; Maciejewska-Skrendo, A. The Role of Peroxisome Proliferator-Activated Receptors and Their Transcriptional Coactivators Gene Variations in Human Trainability: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 1472. https://doi.org/10.3390/ijms19051472
Petr M, Stastny P, Zajac A, Tufano JJ, Maciejewska-Skrendo A. The Role of Peroxisome Proliferator-Activated Receptors and Their Transcriptional Coactivators Gene Variations in Human Trainability: A Systematic Review. International Journal of Molecular Sciences. 2018; 19(5):1472. https://doi.org/10.3390/ijms19051472
Chicago/Turabian StylePetr, Miroslav, Petr Stastny, Adam Zajac, James J. Tufano, and Agnieszka Maciejewska-Skrendo. 2018. "The Role of Peroxisome Proliferator-Activated Receptors and Their Transcriptional Coactivators Gene Variations in Human Trainability: A Systematic Review" International Journal of Molecular Sciences 19, no. 5: 1472. https://doi.org/10.3390/ijms19051472
APA StylePetr, M., Stastny, P., Zajac, A., Tufano, J. J., & Maciejewska-Skrendo, A. (2018). The Role of Peroxisome Proliferator-Activated Receptors and Their Transcriptional Coactivators Gene Variations in Human Trainability: A Systematic Review. International Journal of Molecular Sciences, 19(5), 1472. https://doi.org/10.3390/ijms19051472