Effects of Cyclic Mechanical Stretch on the Proliferation of L6 Myoblasts and Its Mechanisms: PI3K/Akt and MAPK Signal Pathways Regulated by IGF-1 Receptor


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Cyclic Mechanical Stretch on the Proliferation of L6 Myoblasts
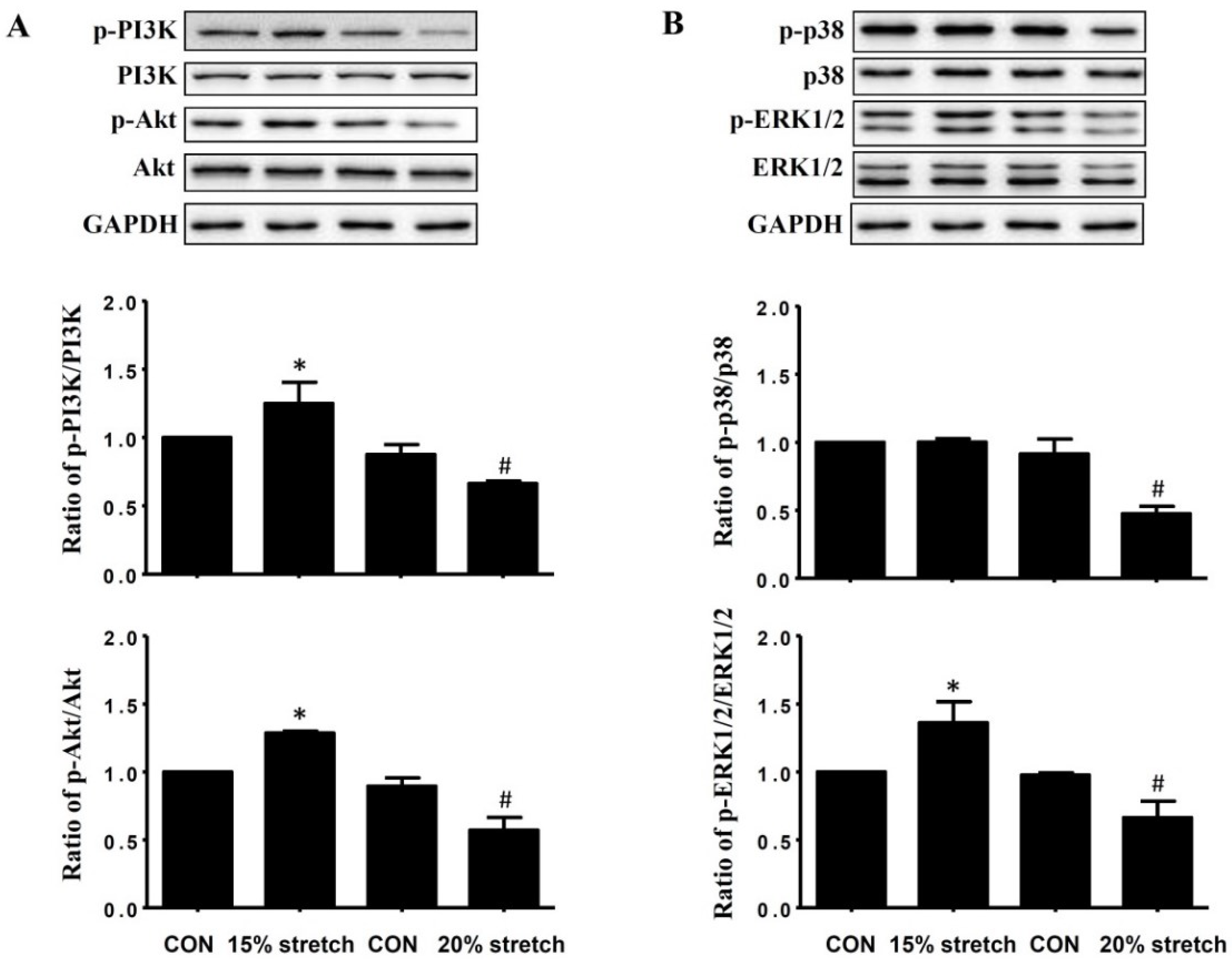
2.2. Effects of Cyclic Mechanical Stretch on the Expressions and Activities of PI3K/Akt and MAPKs (p38 and ERK1/2) in L6 Myoblasts
2.3. Pro-Proliferative Effect of 15% Mechanical Stretch on L6 Myoblasts Was Reversed by PI3K/Akt and ERK1/2 Inhibitors Rather Than p38 Inhibitor
2.4. Effects of Cyclic Mechanical Stretch on IGF-1 Secretion and IGF-1R Protein Level of L6 Myoblasts
2.5. IGF-1 Recombinant Peptide Alleviated 20% Stretch-Induced Proliferation Inhibition of L6 Myoblasts, Accompanied with the Increase of IGF-1R Protein Level as Well as Enhancements of PI3K/Akt and MAPKs (p38 and ERK1/2) Activities
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cyclic Mechanical Stretch
4.3. Detection of Cell Proliferation
4.4. Determination of IGF-1 Concentration in Cell Culture Supernatant
4.5. Treatments of L6 Myoblasts with IGF-1 Recombinant Peptide, and Specific Inhibitors of PI3K, p38, and ERK1/2
4.6. Western Blot
4.7. Statistical Analyses
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Vlietstra, L.; Hendrickx, W.; Waters, D.L. Exercise interventions in healthy older adults with sarcopenia: A systematic review and meta-analysis. Australas J. Ageing 2018. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.O.T.; Hoshino, K.; Nakagawa, T. Influence of exercise intensity on atrophied quadriceps muscle in the rat. J. Phys. Ther. Sci. 2015, 27, 3445–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frystyk, J. Exercise and the growth hormone-insulin-like growth factor axis. Med. Sci. Sports Exerc. 2010, 42, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, A.L.; Pereira, B.C.; Pauli, J.R.; de Souza, C.T.; Teixeira, G.R.; Lira, F.S.; Cintra, D.E.; Ropelle, E.R.; Junior, C.R.; da Silva, A.S. Downhill Running Excessive Training Inhibits Hypertrophy in Mice Skeletal Muscles with Different Fiber Type Composition. J. Cell. Physiol. 2016, 231, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, J.X.; Fry, A.C.; Galpin, A.J.; Sterczala, A.J.; Thomason, D.B.; Moore, C.A.; Weiss, L.W.; Chiu, L.Z. Changes in resting mitogen-activated protein kinases following resistance exercise overreaching and overtraining. Eur. J. Appl. Physiol. 2016, 116, 2401–2413. [Google Scholar] [CrossRef] [PubMed]
- Fujimaki, S.; Machida, M.; Wakabayashi, T.; Asashima, M.; Takemasa, T.; Kuwabara, T. Functional Overload Enhances Satellite Cell Properties in Skeletal Muscle. Stem Cells Int. 2016, 2016, 7619418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazgir, B.; Fathi, R.; Rezazadeh Valojerdi, M.; Mozdziak, P.; Asgari, A. Satellite Cells Contribution to Exercise Mediated Muscle Hypertrophy and Repair. Cell J. 2017, 18, 473–484. [Google Scholar] [PubMed]
- Snijders, T.; Nederveen, J.P.; McKay, B.R.; Joanisse, S.; Verdijk, L.B.; van Loon, L.J.; Parise, G. Satellite cells in human skeletal muscle plasticity. Front. Physiol. 2015, 6, 283. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Fu, S.; Lu, L.; Wang, X. Role of androgen receptor on cyclic mechanical stretch-regulated proliferation of C2C12 myoblasts and its upstream signals: IGF-1-mediated PI3K/Akt and MAPKs pathways. Mol. Cell. Endocrinol. 2017, 450, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Feng, L.; Ruan, M.; Liu, X.; Adriouch, S.; Liao, H. Mechanical-stretch of C2C12 myoblasts inhibits expression of Toll-like receptor 3 (TLR3) and of autoantigens associated with inflammatory myopathies. PLoS ONE 2013, 8, e79930. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Ozawa, K.; Zhao, J.; Kyotani, Y.; Nagayama, K.; Yoshizumi, M. Olmesartan inhibits cultured rat aortic smooth muscle cell death induced by cyclic mechanical stretch through the inhibition of the c-Jun N-terminal kinase and p38 signaling pathways. J. Pharmacol. Sci. 2015, 127, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Yuan, X.; Feng, Y.; Liu, H. Cyclic-stretch induces the apoptosis of myoblast by activation of Caspase-3 protease in a magnitude-dependent manner. Int. J. Biochem. Cell Biol. 2010, 42, 2004–2011. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, X.; Chen, L.; Zhang, Y.; Li, M.; Wang, L.; Ge, C.; Wang, H.; Zhang, M. Mechanical stretch promotes matrix metalloproteinase-2 and prolyl-4-hydroxylase alpha1 production in human aortic smooth muscle cells via Akt-p38 MAPK-JNK signaling. Int. J. Biochem. Cell Biol. 2015, 62, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zhang, Y.; Jiang, H. Signaling pathways mediating the effects of insulin-like growth factor-I in bovine muscle satellite cells. Mol. Cell. Endocrinol. 2013, 372, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Luo, Y.; Huang, Z.; Jia, G.; Liu, G.; Zhao, H. Akirin2 regulates proliferation and differentiation of porcine skeletal muscle satellite cells via ERK1/2 and NFATc1 signaling pathways. Sci. Rep. 2017, 7, 45156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, G.; Liu, D.; Liu, J.; Gao, H.; Yuan, X.; Shen, G. p38 mitogen-activated protein kinase up-regulates NF- κB transcriptional activation through RelA phosphorylation during stretch-induced myogenesis. Biochem. Biophys. Res. Commun. 2010, 391, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Nakai, N.; Kawano, F.; Oke, Y.; Nomura, S.; Ohira, T.; Fujita, R.; Ohira, Y. Mechanical stretch activates signaling events for protein translation initiation and elongation in C2C12 myoblasts. Mol. Cells 2010, 30, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Carson, J.A. Lewis lung carcinoma regulation of mechanical stretch-induced protein synthesis in cultured myotubes. Am. J. Physiol. Cell Physiol. 2015, 310, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, H.; Xu, Y.; Yu, D.; Li, D.; Liu, X.; Du, W. Insulin-like growth factor-1 (IGF-1) promotes myoblast proliferation and skeletal muscle growth of embryonic chickens via the PI3K/Akt signalling pathway. Cell Biol. Int. 2015, 39, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Duan, P.; Guo, P.; Li, D.; Li, S.; Xu, Y.; Zhou, Q. Downregulation of miR-223 and miR-153 mediates mechanical stretch-stimulated proliferation of venous smooth muscle cells via activation of the insulin-like growth factor-1 receptor. Arch. Biochem. Biophys. 2012, 528, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Lv, T.; Wu, Y.; Mu, C.; Liu, G.; Yan, M.; Xu, X.; Wu, H.; Du, J.; Yu, J.; Mu, J. Insulin-like growth factor 1 promotes the proliferation and committed differentiation of human dental pulp stem cells through MAPK pathways. Arch. Oral. Biol. 2016, 72, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Kong, F.; Wang, J.; Lu, Q.; Xu, H.; Qi, T.; Meng, J. Involvement of IGF-1 and MEOX2 in PI3K/Akt1/2 and ERK1/2 pathways mediated proliferation and differentiation of perivascular adipocytes. Exp. Cell Res. 2015, 331, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, J.; Lei, Y.; Han, Z.; Rong, D.; Yu, Q.; Zhao, M.; Tian, J. Low frequency pulsed electromagnetic field promotes C2C12 myoblasts proliferation via activation of MAPK/ERK pathway. Biochem. Biophys. Res. Commun. 2016, 479, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Yue, X.; Luo, D.; Wazir, R.; Wang, J.; Wu, T.; Chen, L.; Liao, B.; Wang, K. Increased proliferation of human bladder smooth muscle cells is mediated by physiological cyclic stretch via the PI3KSGK1Kv1.3 pathway. Mol. Med. Rep. 2013, 8, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Liu, J.; Fan, J.; Li, R.; Li, D.; Yin, J.; Cui, S. Novel evidence that testosterone promotes cell proliferation and differentiation via G protein-coupled receptors in the rat L6 skeletal muscle myoblast cell line. J. Cell. Physiol. 2012, 227, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Lu, A.M.; Wang, Y.; Hong, A.; Chen, Y.; Hu, J.; Li, X.; Qin, Z.H. Chronic resistance training activates autophagy and reduces apoptosis of muscle cells by modulating IGF-1 and its receptors, Akt/mTOR and Akt/FOXO3a signaling in aged rats. Exp. Gerontol. 2013, 48, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Lou, Q.; Zhai, G.; Peng, X.; Cheng, X.; Dai, X.; Zhuo, Z.; Shang, G.; Jin, X.; Chen, X.; et al. Hyperplasia and cellularity changes in IGF-1-overexpressing skeletal muscle of crucian carp. Endocrinology 2014, 155, 2199–2212. [Google Scholar] [CrossRef] [PubMed]
- Bonifacio, A.; Sanvee, G.M.; Brecht, K.; Kratschmar, D.V.; Odermatt, A.; Bouitbir, J.; Krahenbuhl, S. IGF-1 prevents simvastatin-induced myotoxicity in C2C12 myotubes. Arch. Toxicol. 2017, 91, 2223–2234. [Google Scholar] [CrossRef] [PubMed]
- Incerpi, S.; Hsieh, M.T.; Lin, H.Y. Thyroid hormone inhibition in L6 myoblasts of IGF-I-mediated glucose uptake and proliferation: New roles for integrin αvβ3. Am. J. Physiol. Cell Physiol. 2014, 307, 150–161. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Yin, L.; Lin, X.; Lu, J.; Wang, X. Effects of Cyclic Mechanical Stretch on the Proliferation of L6 Myoblasts and Its Mechanisms: PI3K/Akt and MAPK Signal Pathways Regulated by IGF-1 Receptor. Int. J. Mol. Sci. 2018, 19, 1649. https://doi.org/10.3390/ijms19061649
Fu S, Yin L, Lin X, Lu J, Wang X. Effects of Cyclic Mechanical Stretch on the Proliferation of L6 Myoblasts and Its Mechanisms: PI3K/Akt and MAPK Signal Pathways Regulated by IGF-1 Receptor. International Journal of Molecular Sciences. 2018; 19(6):1649. https://doi.org/10.3390/ijms19061649
Chicago/Turabian StyleFu, Shaoting, Lijun Yin, Xiaojing Lin, Jianqiang Lu, and Xiaohui Wang. 2018. "Effects of Cyclic Mechanical Stretch on the Proliferation of L6 Myoblasts and Its Mechanisms: PI3K/Akt and MAPK Signal Pathways Regulated by IGF-1 Receptor" International Journal of Molecular Sciences 19, no. 6: 1649. https://doi.org/10.3390/ijms19061649