Physiological and Proteomics Analyses Reveal Low-Phosphorus Stress Affected the Regulation of Photosynthesis in Soybean

 ,
,
Abstract
1. Introduction
2. Results
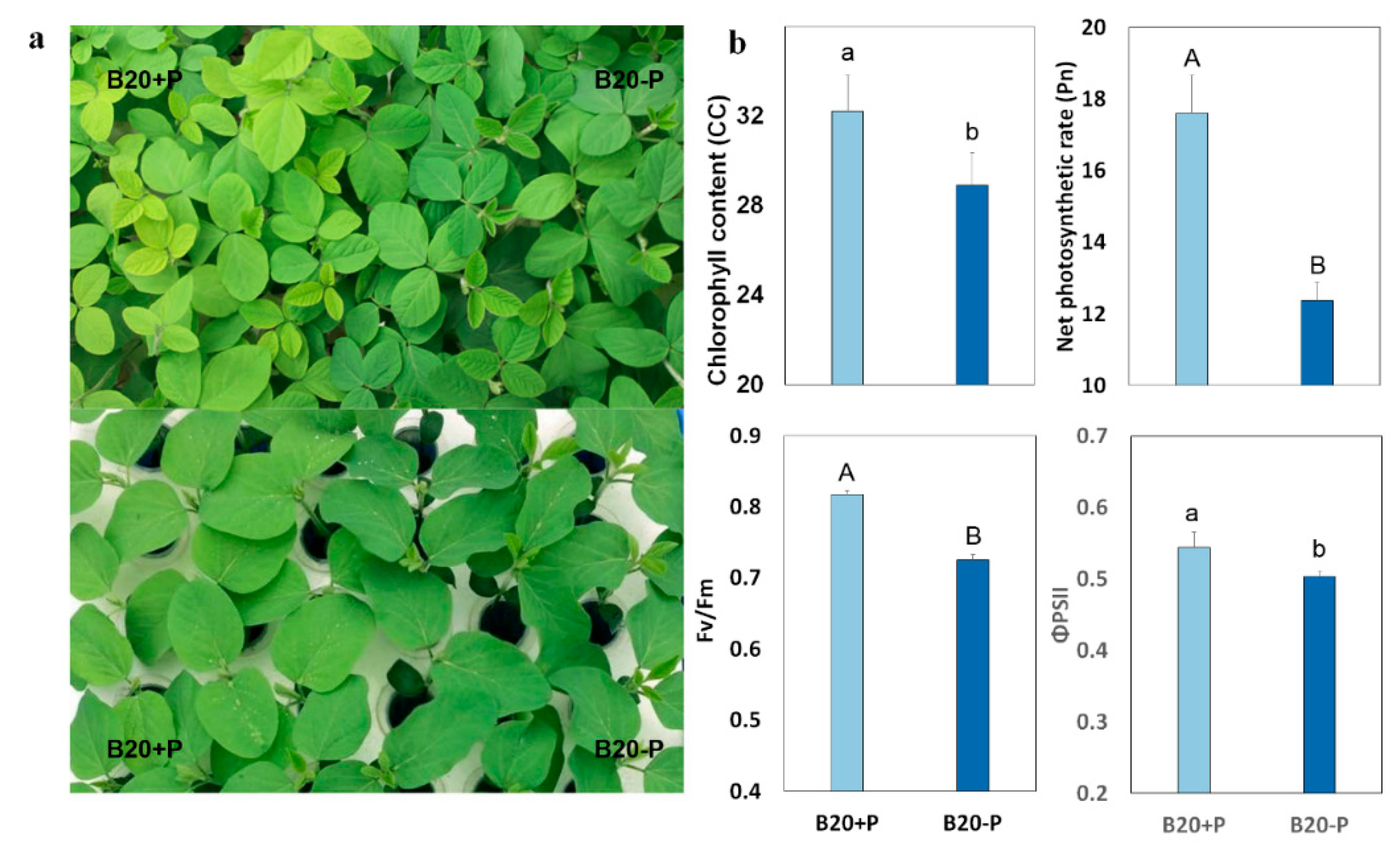
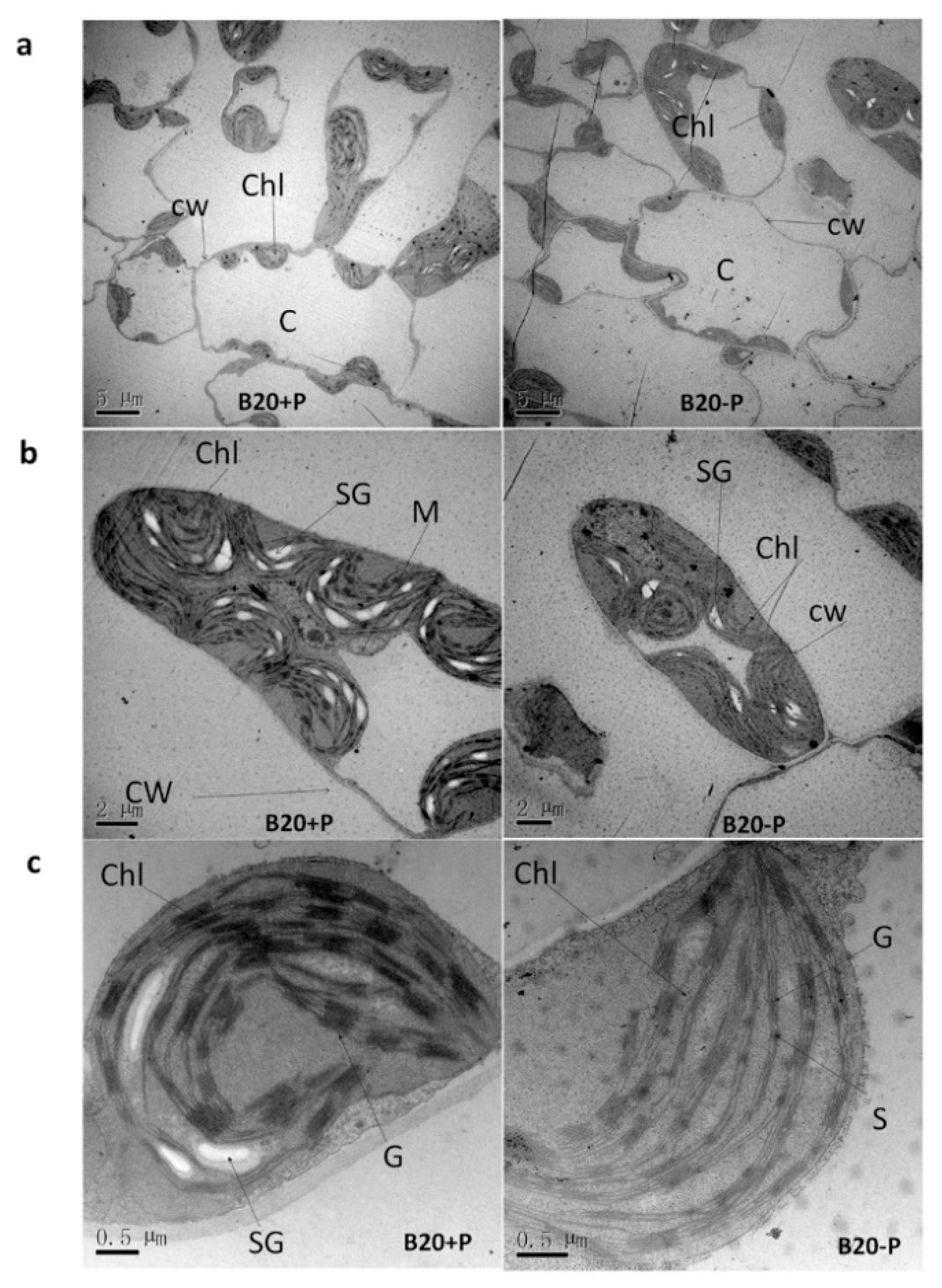
2.1. Low-P Stress Affected Soybean Plant Growth and Leaf Structure
2.2. Low-P Stress Affected Photosynthetic Functions
2.3. Low-P Stress Affected the Expression of the Genes Involved in Photosynthesis
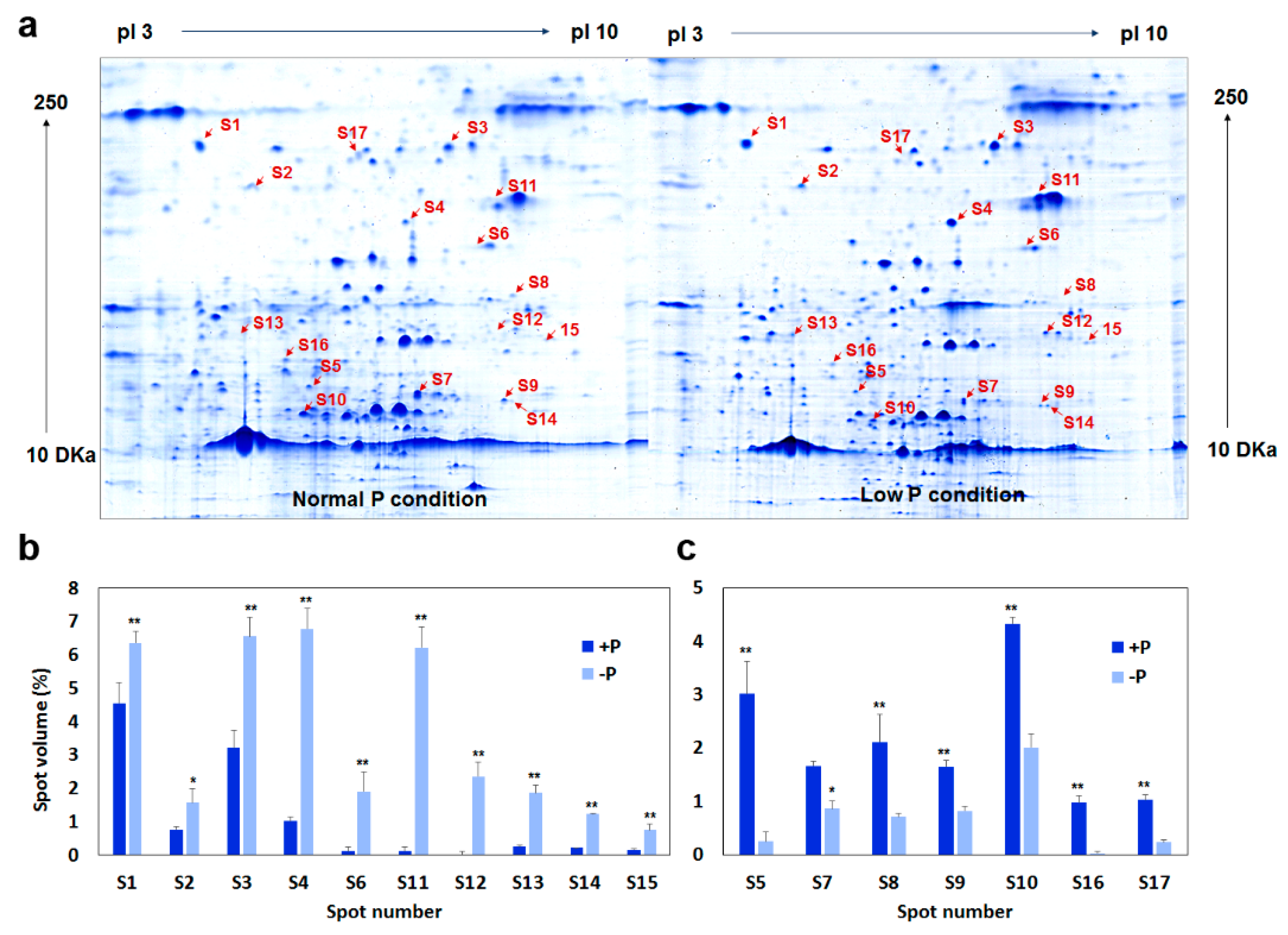
2.4. Photosynthesis-Related Proteins Were Differentially Expressed after Low-P Stress
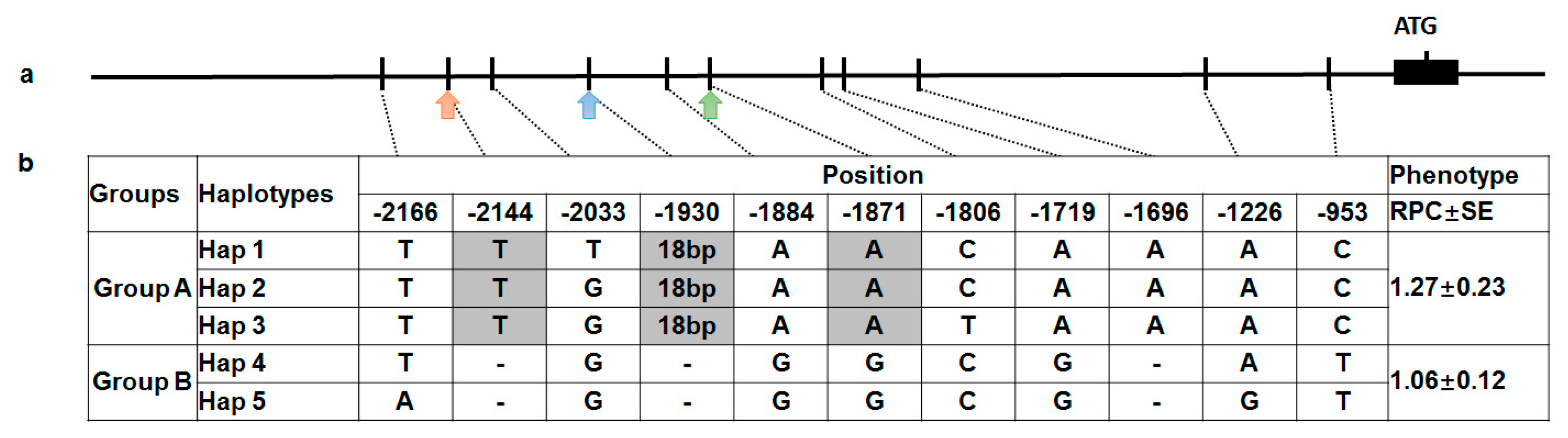
2.5. Association Analysis for the SNPs of GmRCAβ Promoter
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Hydroponics Experiments
4.2. Measurement of P Efficiency and Photosynthesis-Related Traits
4.3. Determination of Gene Expression by qPCR
4.4. Leaf Anatomical Structure by Transmission Electron Microscope
4.5. Protein Extraction
4.6. Two-Dimensional Isoelectric Focusing (2D-IEF)/SDS-PAGE
4.7. Image and Statistical Analysis
4.8. Peptide Mass Fingerprinting (PMF) Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| APA | Acid phosphatase activity |
| ATPase | ATP synthase |
| CC | Chlorophyll content |
| Ci | intercellular CO2 concentration |
| Fv/Fm | Maximum quantum efficiency of PSII |
| Gs | Stomatal conductance |
| NPQ | Non-photochemical quenching |
| PAE | Phosphorus absorption efficiency |
| PC | Phosphorus concentration |
| Pn | Net photosynthetic rate |
| PUE | Phosphorus use efficiency |
| qP | Photochemical quenching |
| RDW | Root dry weight |
| RuBPase | Ribulose-1,5-bisphosphate carboxylase |
| R/S | Root: shoot ratio |
| TDW | Total dry weight |
| Tr | Transpiration rate |
| SDW | Shoot dry weight |
| ΦPSII | Quantum efficiency of PSII |
References
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Y.; Zhang, H.; Chu, S.; Zhang, X.; Yin, D.; Yu, D.; Zhang, D. A genetic relationship between phosphorus efficiency and photosynthetic traits in soybean as revealed by QTL analysis using a high-density genetic map. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, H.; Song, J.; Wu, W.; Li, K.; Zhang, J. Physiological and comparative proteome analyses reveal low-phosphate tolerance and enhanced photosynthesis in a maize mutant owing to reinforced inorganic phosphate recycling. BMC Plant Biol. 2016, 16, 129. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61. [Google Scholar] [CrossRef] [PubMed]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Hill, J.H.; Loebenstein, G.; Thottappilly, G. Soybean. Virus and Virus-like Diseases of Major Crops in Developing Countries; Springer: New York, NY, USA, 2003; pp. 377–395. [Google Scholar]
- Zhang, D.; Song, H.; Cheng, H.; Hao, D.; Wang, H.; Kan, G.; Jin, H.; Yu, D. The acid phosphatase-encoding gene GmACP1 contributes to soybean tolerance to low-phosphorus stress. PLoS Genet. 2014, 10, e1004061. [Google Scholar] [CrossRef] [PubMed]
- Gowin, S. Phosphorus and potassium effects on soybeans. In Horticultural Crops Plant Nutrition Series; Department of Horticultural Science, Texas A&M University: College Station, TX, USA, 1997; pp. 44–51. [Google Scholar]
- Batjes, N.H. A world dataset of derived soil properties by FAO-UNESCO soil unit for global modelling. Soil Use Manag. 2010, 13, 9–16. [Google Scholar] [CrossRef]
- Yamaji, N.; Takemoto, Y.; Miyaji, T.; Mitani-Ueno, N.; Yoshida, K.T.; Jian, F.M. Reducing phosphorus accumulation in rice grains with an impaired transporter in the node. Nature 2017, 541, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, H.; Chu, S.; Li, H.; Chi, Y.; Triebwasser-Freese, D.; Lv, H.; Yu, D. Integrating QTL mapping and transcriptomics identifies candidate genes underlying QTLs associated with soybean tolerance to low-phosphorus stress. Plant Mol. Biol. 2017, 93, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Delaney, K.J.; Weaver, D.K.; Peterson, R.K.D. Photosynthesis and yield reductions from wheat stem sawfly (Hymenoptera: Cephidae): Interactions with wheat solidness, water stress, and phosphorus deficiency. J. Econ. Entomol. 2010, 103, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Reddy, V.R. Combined effects of phosphorus nutrition and elevated carbon dioxide concentration on chlorophyll fluorescence, photosynthesis, and nutrient efficiency of cotton. J. Nutr. Soil Sci. 2014, 177, 892–902. [Google Scholar] [CrossRef]
- Lan, P.; Li, W.; Schmidt, W. Complementary proteome and transcriptome profiling in phosphate-deficient Arabidopsis roots reveals multiple levels of gene regulation. Mol. Cell Proteom. 2012, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.K.; Zheng, J.W.; Niu, Y.F.; Yang, W.D.; Liu, J.S.; Li, H.Y. Systems-level analysis of the metabolic responses of the diatom Phaeodactylum tricornutum to phosphorus stress. Environ. Microbial. 2014, 16, 1793–1807. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.; Woo, K.C.; Wong, S.C. Effects of phosphorus nutrition on the response of photosynthesis to CO2 and O2, activation of ribulose bisphosphate carboxylase and amounts of ribulose bisphosphate and 3-phosphoglycerate in spinach leaves. Photosyn. Res. 1988, 15, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Badgujar, G.; Reddy, V.R.; Fleisher, D.H.; Bunce, J.A. Carbon dioxide diffusion across stomata and mesophyll and photo-biochemical processes as affected by growth CO2 and phosphorus nutrition in cotton. J. Plant Physiol. 2013, 170, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.; Yin, Z.; Hao, D.; Zhang, J.; Song, H.; Ning, A.; Xu, X.; Yu, D. Variation in Rubisco activase (RCAβ) gene promoters and expression in soybean [Glycine max (L.) Merr]. J. Exp. Bot. 2014, 65, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhang, Z.; Deng, D.; Chao, M.; Gao, Q.; Wang, Y.; Yang, Z.; Bian, Y.; Hao, D.; Xu, C. Characterization of Rubisco activase genes in maize: An alpha-isoform gene functions alongside a beta-isoform gene. Plant Physiol. 2014, 164, 2096–2106. [Google Scholar] [CrossRef] [PubMed]
- Toorchi, M.; Yukawa, K.; Nouri, M.Z.; Komatsu, S. Proteomics approach for identifying osmotic-stress-related proteins in soybean roots. Peptides 2009, 30, 2108–2117. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.B.; Eticha, D.; Fuhrs, H.; Heintz, D.; Ayoub, D.; Van Dorsselaer, A.; Schlingmann, B.; Rao, I.M.; Braun, H.P.; Horst, W.J. Proteomic and phosphoproteomic analysis of polyethylene glycol-induced osmotic stress in root tips of common bean (Phaseolus vulgaris L.). J. Exp. Bot. 2013, 64, 5569–5586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chu, S.; Zhang, D. Transcriptome dataset of soybean (Glycine max) grown under phosphorus-deficient and-sufficient conditions. Data 2017, 2, 17. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H.; Sun, H.X.; Bowler, C.; Chua, N.H. Noncoding and coding transcriptome responses of a marine diatom to phosphate fluctuations. New Phytol. 2016, 210, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Takahashi, S.; Makino, A.; Price, G.D.; Badger, M.R.; Caemmerer, S.V. The roles of ATP synthase and the cytochrome b6/f complexes in limiting chloroplast electron transport and determining photosynthetic capacity. Plant Physiol. 2011, 155, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Thuynsma, R.; Kleinert, A.; Kossmann, J.; Valentine, A.J.; Hills, P.N. The effects of limiting phosphate on photosynthesis and growth of Lotus japonicus. S. Afr. J. Bot. 2016, 104, 244–248. [Google Scholar] [CrossRef]
- Jiang, C.D.; Wang, X.; Gao, H.Y.; Shi, L.; Chow, W.S. Systemic regulation of leaf anatomical structure, photosynthetic performance, and high-light tolerance in sorghum. Plant Physiol. 2011, 155, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Sowiński, P.; Szczepanik, J.; Minchin, P.E.H. On the mechanism of C4 photosynthesis intermediate exchange between Kranz mesophyll and bundle sheath cells in grasses. J. Exp. Bot. 2008, 59, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, L.N.; Jiang, C.D.; Liu, Y.J.; Shi, L. Effects of mutual shading on the regulation of photosynthesis in field-grown sorghum. J. Photochem. Photobiol. B 2014, 137, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Silva, A.E.; Gore, M.A.; Andrade-Sanchez, P.; French, A.N.; Hunsaker, D.J.; Salvucci, M.E. Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field. Environ. Exp. Bot. 2012, 83, 1–11. [Google Scholar] [CrossRef]
- He, Y.; Yu, C.; Zhou, L.; Chen, Y.; Liu, A.; Jin, J.; Hong, J.; Qi, Y.; Jiang, D. Rubisco decrease is involved in chloroplast protrusion and Rubisco-containing body formation in soybean (Glycine max. L) under salt stress. Plant Physiol. Biochem. 2014, 74, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Govindjee; Bosa, K.; Kościelniak, J.; Żukgołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation in two syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Timmermans, K.R.; Wagt, B.V.D.; Veldhuis, M.J.W.; Maatman, A.; Baar, H.J.W.D. Physiological responses of three species of marine pico-phytoplankton to ammonium, phosphate, iron and light limitation. J. Sea Res. 2005, 53, 109–120. [Google Scholar] [CrossRef]
- Perdomo, J.A.; Carmo-Silva, E. Improving the activation of Rubisco in wheat. In Proceedings of the 26th Congress of the Scandinavian Plant Physiology Society, Stockholm, Sweden, 9–13 August 2015. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Baldwin, D.S.; Beattie, J.K.; Coleman, L.M.; Jones, D.R. Hydrolysis of an organophosphate ester by manganese dioxide. Environ. Sci. Technol. 2001, 4, 713–716. [Google Scholar] [CrossRef]
- Yin, Z.; Meng, F.; Song, H.; He, X.; Xu, X.; Yu, D. Mapping quantitative trait loci associated with chlorophyll a fluorescence parameters in soybean (Glycine max (L.) Merr). Planta 2010, 231, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Salah, S.M.; Guan, Y.; Cao, D.; Li, J.; Aamir, N.; Hu, Q.; Hu, W.; Ning, M.; Hu, J. Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci. Rep. 2015, 5, 14278. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Rui, F.; Zhao, T.J.; Yu, J.J.; Yu, D.Y. Seed storage protein components are associated with curled cotyledon phenotype in soybean. Afr. J. Biotechnol. 2009, 8, 6063–6067. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, J.; Cao, M.; Zhao, K.; Shao, J.; Lei, T.; Yin, J.; Hill, G.G.; Xu, N.; Liu, S. Proteomic changes in rice leaves during development of field-grown rice plants. Proteomics 2005, 5, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Katam, R.; Basha, S.M.; Suravajhala, P.; Pechan, T. Analysis of peanut leaf proteome. J. Proteome Res. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zhang, E.; Li, H.; Cheng, X.; Zhu, F.; Hong, Y.; Liao, B.; Liu, S.; Liang, X. Comparative proteomics analysis of developing peanut aerial and subterranean pods identifies pod swelling related proteins. J. Proteome 2013, 91, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Mascot. Available online: http://www.matrixscience.com (accessed on 22 March 2016).
- Integrating Genetics and Genomics to Advance Soybean Research. Available online: http://www.soybase.org (accessed on 25 March 2016).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits Name (Units) | Traits Abbreviation | Means ± SD | Means ± SD |
|---|---|---|---|
| B20 + P | B20 − P | ||
| Growth-related traits | |||
| Shoot dry weight (g) | SDW | 0.424 ± 0.022 aA | 0.352 ± 0.047 bB |
| Root dry weight (g) | RDW | 0.073 ± 0.00 bB | 0.096 ± 0.008 aA |
| Total dry weight (g) | TDW | 0.497 ± 0.019 aA | 0.448 ± 0.053 bA |
| Root: shoot ratio | R/S | 0.172 ± 0.0174 bB | 0.274 ± 0.027 aA |
| Acid phosphatase activity | APA | 1.33 ± 0.12 aA | 3.24 ± 0.40 bB |
| ATP synthase (μmolPi·mgprot−1·h−1) | ATPase | 18.48 ± 0.92 aA | 16.27 ± 0.94 aA |
| Ribulose-1,5-bisphosphate carboxylase | RuBPase | 37.31 ± 1.02 aA | 31.41 ± 0.95 bB |
| P efficiency-related traits | |||
| Phosphorus concentration (mmol·L−1) | PC | 23.47 ± 0.99 aA | 7.19 ± 0.69 bB |
| Phosphorus absorption efficiency | PUP | 11.67 ± 0.88 aA | 3.24 ± 0.64 bB |
| Phosphorus use efficiency | PUE | 0.04 ± 0.01 bB | 0.14 ± 0.01 aA |
| Photosynthesis-related traits | |||
| Chlorophyll content (mg·g−1) | CC | 32.2 ± 0.72 aA | 28.9 ± 1.65 aA |
| Net photosynthetic rate (μmol·m2·s−1) | Pn | 17.60 ± 1.05 aA | 12.37 ± 0.50 bB |
| Stomatal conductance (mmol·m−2·s−1) | Gs | 210 ± 8.19 aA | 240.33 ± 5.51 bB |
| Transpiration rate (g· m−2·h−1) | Tr | 4.44 ± 0.35 aA | 2.97 ± 0.46 bB |
| Intercellular CO2 concentration | Ci | 0.25 ± 0.04 aA | 0.19 ± 0.01 bB |
| Maximum quantum efficiency of PSII | Fv/Fm | 0.82 ± 0.01 aA | 0.72 ± 0.01 bB |
| Quantum efficiency of PSII | ΦPSII | 0.54 ± 0.02 aA | 0.50 ± 0.01 bA |
| Photochemical quenching | qP | 0.72 ± 0.02 aA | 0.62 ± 0.03 bB |
| Non-photochemical quenching | NPQ | 0.38 ± 0.02 aA | 0.46 ± 0.02 bB |
| Genes | Fold Change (−P/+P) | Expression | Annotation | |
|---|---|---|---|---|
| qPCR | Transcriptome | |||
| Glyma.01G095900 | 1.32 | 2.43 | down | Photosystem II PsbZ protein |
| Glyma.03G065000 | 3.33 | 10.07 | down | Photosystem II PsbH protein (PsbH) |
| Glyma.03G068100 | 5.89 | 2.07 | down | Ribulose-1,5-bisphosphate carboxylase |
| Glyma.05G022900 | 2.79 | 8.75 | down | Photosystem I subunit III (PsaF) |
| Glyma.06G277500 | 1.79 | 2.16 | down | Phosphoenolpyruvate carboxylase |
| Glyma.08G201200 | 1.29 | 4.50 | up | NADP+-dependent malic enzyme |
| Glyma.12G096200 | 1.56 | 6.80 | down | ATP synthesis coupled proton transport |
| Glyma.13G149800 | 1.19 | 2.20 | up | Pyruvate kinase activity |
| Glyma.15G188400 | 1.51 | 3.71 | down | Photosystem II (PsbN) |
| Glyma.15G248600 | 1.72 | 1.42 | down | Photosystem II PsbK protein |
| Glyma.17G020600 | 2.04 | 2.03 | down | Pyruvate orthophosphate dikinase |
| Glyma.18G036400 | 1.63 | 2.51 | down | Rubisco activase |
| Glyma.19G190100 | 1.27 | 2.14 | up | Pyruvate kinase, barrel domain |
| Glyma.20G122500 | 1.33 | 2.15 | down | Fructose-biphosphate aldolase |
| Spot No. | Gene ID | Matched Protein Name | Pattern | pI | Mw | Coverage Rate (%) | Protein Score | FC (−P/+P) |
|---|---|---|---|---|---|---|---|---|
| S1 | Glyma.11G108500 | Chloroplast movement | Up | 4.52 | 95.2 | 33.45 | 73.1 | 1.57 |
| S2 | Glyma.06G277500 | Phosphoenolpyruvate carboxylase | Up | 5.33 | 75.1 | 30.60 | 67.49 | 2.05 |
| S3 | Glyma.19G224200 | Phytochrome A | Up | 6.69 | 94.6 | 28.56 | 68.94 | 2.03 |
| S4 | Glyma.16G173100 | Pyruvate kinase | Up | 6.54 | 62.2 | 38 | 73.32 | 6.65 |
| S5 | Glyma.19G186000 | Triosephosphate isomerase (TPI) | Down | 6.33 | 33.2 | 45.60 | 65.4 | 0.36 |
| S6 | Glyma.10G197700 | Malate dehydrogenase activity | Up | 6.63 | 48.8 | 19.90 | 69.9 | 8.60 |
| S7 | Glyma.13G078600 | ATP synthase alpha/beta family | Down | 6.62 | 24.4 | 19.68 | 70.3 | 0.52 |
| S8 | Glyma.18G036400 | Rubisco activase | Down | 8.01 | 45.5 | 7.35 | 66.26 | / |
| S9 | Glyma.08G142700 | Iron-sulfur cluster assembly protein | Down | 7.99 | 18.1 | 11.50 | 71.65 | 0.50 |
| S10 | Glyma.20G090000 | Phytochrome A | Down | 6.35 | 15.2 | 6.30 | 69.58 | 0.23 |
| S11 | Glyma.06G091500 | Phosphoenolpyruvate carboxykinase | Up | 7.55 | 70.9 | 8.59 | 63.2 | / |
| S12 | Glyma.05G025300 | Ribulose-phosphate 3-epimerase | Up | 7.89 | 29.6 | 21.80 | 64.75 | / |
| S13 | Glyma.05G022900 | Photosystem I subunit III | Up | 5.01 | 28.6 | 4.90 | 61.49 | / |
| S14 | Glyma.15G266300 | Myosin ATPase | Up | 8.69 | 17.6 | 24.10 | 73.6 | / |
| S15 | Glyma.15G018000 | Electron carrier activity | Up | 8.58 | 22.3 | 35.20 | 72.63 | / |
| S16 | Glyma.07G185400 | Malate dehydrogenase | Down | 5.82 | 37.0 | 28.40 | 77.26 | 0.31 |
| S17 | Glyma.13G089200 | Cryptochrome 1 | Down | 6.32 | 77.0 | 2.90 | 62.19 | / |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, S.; Li, H.; Zhang, X.; Yu, K.; Chao, M.; Han, S.; Zhang, D. Physiological and Proteomics Analyses Reveal Low-Phosphorus Stress Affected the Regulation of Photosynthesis in Soybean. Int. J. Mol. Sci. 2018, 19, 1688. https://doi.org/10.3390/ijms19061688
Chu S, Li H, Zhang X, Yu K, Chao M, Han S, Zhang D. Physiological and Proteomics Analyses Reveal Low-Phosphorus Stress Affected the Regulation of Photosynthesis in Soybean. International Journal of Molecular Sciences. 2018; 19(6):1688. https://doi.org/10.3390/ijms19061688
Chicago/Turabian StyleChu, Shanshan, Hongyan Li, Xiangqian Zhang, Kaiye Yu, Maoni Chao, Suoyi Han, and Dan Zhang. 2018. "Physiological and Proteomics Analyses Reveal Low-Phosphorus Stress Affected the Regulation of Photosynthesis in Soybean" International Journal of Molecular Sciences 19, no. 6: 1688. https://doi.org/10.3390/ijms19061688
APA StyleChu, S., Li, H., Zhang, X., Yu, K., Chao, M., Han, S., & Zhang, D. (2018). Physiological and Proteomics Analyses Reveal Low-Phosphorus Stress Affected the Regulation of Photosynthesis in Soybean. International Journal of Molecular Sciences, 19(6), 1688. https://doi.org/10.3390/ijms19061688