Molecular Regulation of Differentiation in Early B-Lymphocyte Development

{kind=link}
{kind=link}
Abstract
1. Introduction
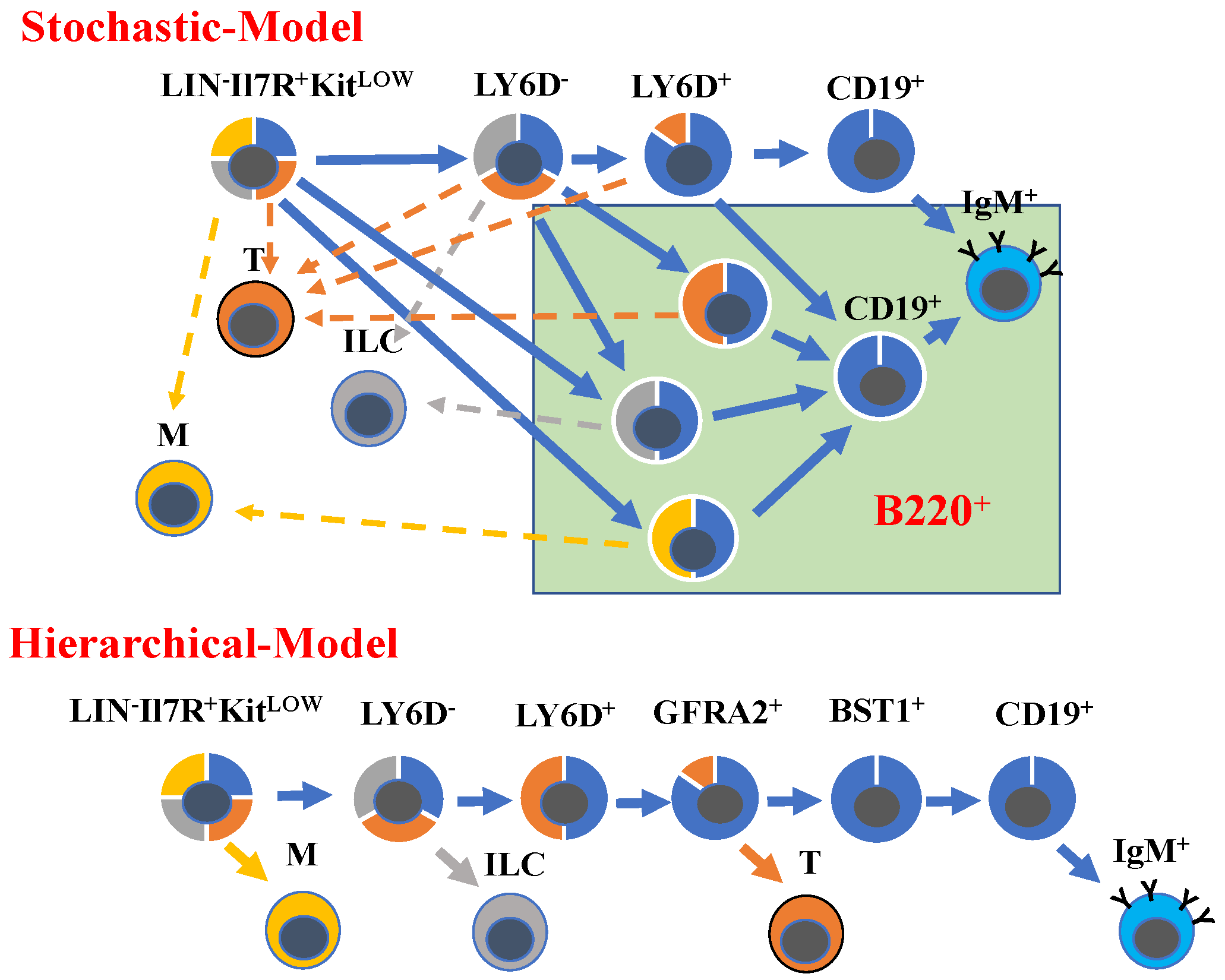
2. Resolving Developmental Trajectories in B-Cell Development
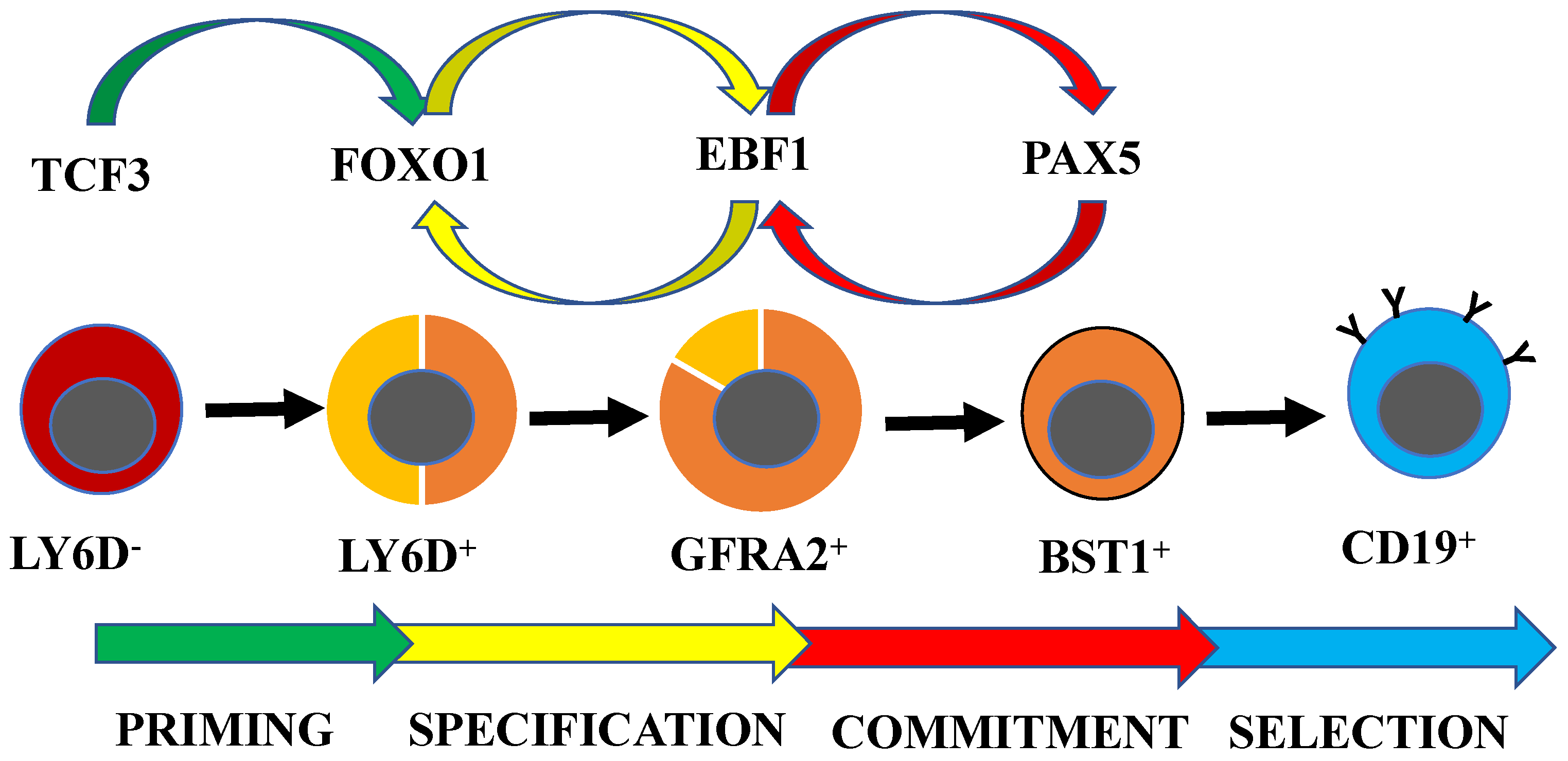
3. Transcription Factor Networks Regulating Early B-Cell Development
4. Transcription Factor Networks Link Development to B-Lymphoid Malignancies
5. Integration of External Signals and Transcription Factor Networks in Early B-Cell Development
6. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Melchers, F. Checkpoints that control B cell development. J. Clin. Investig. 2015, 125, 2203–2210. [Google Scholar] [CrossRef] [PubMed]
- Ghia, P.; ten Boekel, E.; Rolink, A.G.; Melchers, F. B-cell development: A comparison between mouse and man. Immunol. Today 1998, 19, 480–485. [Google Scholar] [CrossRef]
- Ghia, P.; ten Boekel, E.; Sanz, E.; de la Hera, A.; Rolink, A.; Melchers, F. Ordering of human bone marrow B lymphocyte precursors by single-cell polymerase chain reaction analyses of the rearrangement status of the immunoglobulin H and L chain gene loci. J. Exp. Med. 1996, 184, 2217–2229. [Google Scholar] [CrossRef] [PubMed]
- Ghia, P.; Gratwohl, A.; Signer, E.; Winkler, T.H.; Melchers, F.; Rolink, A.G. Immature B cells from human and mouse bone marrow can change their surface light chain expression. Eur. J. Immunol. 1995, 25, 3108–3114. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, R.; Prasad, M.A.; Ungerback, J.; Sigvardsson, M. Transcription factor networks in B-cell differentiation link development to acute lymphoid leukemia. Blood 2015, 126, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Adolfsson, J.; Mansson, R.; Buza-Vidas, N.; Hultquist, A.; Liuba, K.; Jensen, C.T.; Bryder, D.; Yang, L.; Borge, O.J.; Thoren, L.A.; et al. Identification of flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revised road map for adult blood lineage commitment. Cell 2005, 121, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Weissman, I.L.; Akashi, K. Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell 1997, 91, 661–672. [Google Scholar] [CrossRef]
- Hardy, R.R.; Carmack, C.E.; Shinton, S.A.; Kemp, J.D.; Hayakawa, K. Resolution and characterization of pro-B and pre-pro-B cell stages in normal mouse bone marrow. J. Exp. Med. 1991, 173, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Possot, C.; Schmutz, S.; Chea, S.; Boucontet, L.; Louise, A.; Cumano, A.; Golub, R. Notch signaling is necessary for adult, but not fetal, development of RORγt(+) innate lymphoid cells. Nat. Immunol. 2011, 12, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.L.; Tangen, S.N.; Seaburg, L.M.; Thapa, P.; Gwin, K.A.; Shapiro, V.S. Separation of plasmacytoid dendritic cells from B-cell-biased lymphoid progenitor (BLP) and Pre-pro B cells using PDCA-1. PLoS ONE 2013, 8, e78408. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.H.; Aifantis, I.; Scimone, M.L.; von Andrian, U.H.; Reizis, B.; von Boehmer, H.; Gounari, F. Efficient thymic immigration of B220+ lymphoid-restricted bone marrow cells with T precursor potential. Nat. Immunol. 2003, 4, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Mansson, R.; Zandi, S.; Welinder, E.; Tsapogas, P.; Sakaguchi, N.; Bryder, D.; Sigvardsson, M. Single-cell analysis of the common lymphoid progenitor compartment reveals functional and molecular heterogeneity. Blood 2010, 115, 2601–2609. [Google Scholar] [CrossRef] [PubMed]
- Rolink, A.; Melchers, F. B lymphopoiesis in the mouse. Adv. Immunol. 1993, 53, 123–156. [Google Scholar] [PubMed]
- Hardy, R.R.; Hayakawa, K. B-lineage differentiation stages resolved by multiparameter flow cytometry. Ann. N. Y. Acad. Sci. 1995, 764, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Rumfelt, L.L.; Zhou, Y.; Rowley, B.M.; Shinton, S.A.; Hardy, R.R. Lineage specification and plasticity in CD19− early B cell precursors. J. Exp. Med. 2006, 203, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Wasserman, R.; Hayakawa, K.; Hardy, R.R. Identification of the earliest B lineage stage in mouse bone marrow. Immunity 1996, 5, 527–535. [Google Scholar] [CrossRef]
- Rolink, A.; Grawunder, U.; Winkler, T.H.; Karasuyama, H.; Melchers, F. IL-2 receptor α chain (CD25, TAC) expression defines a crucial stage in pre-B cell development. Int. Immunol. 1994, 6, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.T.; Lang, S.; Somasundaram, R.; Soneji, S.; Sigvardsson, M. Identification of Stage-Specific Surface Markers in Early B Cell Development Provides Novel Tools for Identification of Progenitor Populations. J. Immunol. 2016, 197, 1937–1944. [Google Scholar] [CrossRef] [PubMed]
- Rolink, A.; ten Boekel, E.; Melchers, F.; Fearon, D.T.; Krop, I.; Andersson, J. A subpopulation of B220+ cells in murine bone marrow does not express CD19 and contains natural killer cell progenitors. J. Exp. Med. 1996, 183, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Balciunaite, G.; Ceredig, R.; Massa, S.; Rolink, A.G. A B220+ CD117+ CD19± hematopoietic progenitor with potent lymphoid and myeloid developmental potential. Eur. J. Immunol. 2005, 35, 2019–2030. [Google Scholar] [CrossRef] [PubMed]
- Alberti-Servera, L.; von Muenchow, L.; Tsapogas, P.; Capoferri, G.; Eschbach, K.; Beisel, C.; Ceredig, R.; Ivanek, R.; Rolink, A. Single-cell RNA sequencing reveals developmental heterogeneity among early lymphoid progenitors. EMBO J. 2017, 36, 3619–3633. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.J.; Bhandoola, A. The earliest thymic progenitors for T cells possess myeloid lineage potential. Nature 2008, 452, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Katsura, Y. Redefinition of lymphoid progenitors. Nat. Rev. Immunol. 2002, 2, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Pereira de Sousa, A.; Berthault, C.; Granato, A.; Dias, S.; Ramond, C.; Kee, B.L.; Cumano, A.; Vieira, P. Inhibitors of DNA binding proteins restrict T cell potential by repressing Notch1 expression in Flt3-negative common lymphoid progenitors. J. Immunol. 2012, 189, 3822–3830. [Google Scholar] [CrossRef] [PubMed]
- Martensson, I.L.; Melchers, F.; Winkler, T.H. A transgenic marker for mouse B lymphoid precursors. J. Exp. Med. 1997, 185, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Mansson, R.; Zandi, S.; Anderson, K.; Martensson, I.L.; Jacobsen, S.E.; Bryder, D.; Sigvardsson, M. B-lineage commitment prior to surface expression of B220 and CD19 on hematopoietic progenitor cells. Blood 2008, 112, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Karasuyama, H.; Kudo, A.; Melchers, F. The proteins encoded by the VpreB and l5 pre-B cell-specific genes can associate with each other and with the m heavy chain. J. Exp. Med. 1990, 172, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Rolink, A.; Karasuyama, H.; Grawunder, U.; Haasner, D.; Kudo, A.; Melchers, F. B cell development in mice with a defective lambda 5 gene. Eur. J. Immunol. 1993, 23, 1284–1288. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, D.; Kudo, A.; Schaal, S.; Muller, W.; Melchers, F.; Rajewsky, K. A critical role of lambda 5 protein in B cell development. Cell 1992, 69, 823–831. [Google Scholar] [CrossRef]
- Kudo, A.; Thalmann, P.; Sakaguchi, N.; Davidson, W.F.; Pierce, J.H.; Kearney, J.F.; Reth, M.; Rolink, A.; Melchers, F. The expression of the mouse VpreB/lambda 5 locus in transformed cell lines and tumors of the B lineage differentiation pathway. Int. Immunol. 1992, 4, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Karasuyama, H.; Rolink, A.; Shinkai, Y.; Young, F.; Alt, F.W.; Melchers, F. The expression of Vpre-B/lambda 5 surrogate light chain in early bone marrow precursor B cells of normal and B cell-deficient mutant mice. Cell 1994, 77, 133–143. [Google Scholar] [CrossRef]
- Inlay, M.A.; Bhattacharya, D.; Debashis, S.; Serwold, T.; Seita, J.; Karsunky, H.; Plevritis, S.K.; Dill, D.L.; Weissman, I.L. Ly6d marks the earliest stage of B-cell specification and identifies the branchpoint between B-cell and T-cell development. Genes Dev. 2009, 23, 2376–2381. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.T.; Ahsberg, J.; Sommarin, M.N.E.; Strid, T.; Somasundaram, R.; Okuyama, K.; Ungerback, J.; Kupari, J.; Airaksinen, M.S.; Lang, S.; et al. Dissection of progenitor compartments resolves developmental trajectories in B-lymphopoiesis. J. Exp. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bendall, S.C.; Davis, K.L.; Amir el, A.D.; Tadmor, M.D.; Simonds, E.F.; Chen, T.J.; Shenfeld, D.K.; Nolan, G.P.; Pe’er, D. Single-cell trajectory detection uncovers progression and regulatory coordination in human B cell development. Cell 2014, 157, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ma, A.; Young, F.; Alt, F.W. IL-2 receptor α chain expression during early B lymphocyte differentiation. Int. Immunol. 1994, 6, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Hardy, R.R.; Kincade, P.W.; Dorshkind, K. The protean nature of cells in the B lymphocyte lineage. Immunity 2007, 26, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Milne, C.D.; Fleming, H.E.; Paige, C.J. IL-7 does not prevent pro-B/pre-B cell maturation to the immature/sIgM+ stage. Eur. J. Immunol. 2004, 34, 2647–2655. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Krause, D.; Greaves, M.; Sharkis, S.; Dexter, M.; Heyworth, C.; Enver, T. Multilineage gene expression precedes commitment in the hemopoietic system. Genes Dev. 1997, 11, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Mansson, R.; Hultquist, A.; Luc, S.; Yang, L.; Anderson, K.; Kharazi, S.; Al-Hashmi, S.; Liuba, K.; Thoren, L.; Adolfsson, J.; et al. Molecular evidence for hierarchical transcriptional lineage priming in fetal and adult stem cells and multipotent progenitors. Immunity 2007, 26, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Arinobo, Y.; Mizuno, S.; Chong, Y.; Shigematsu, H.; Iino, T.; Iwasaki, H.; Graf, T.; Mayfield, R.; Chan, S.; Kastner, P.; et al. Reciprocal activation of Gata-1 and PU.1 marks initial specification of hematopoietic stem cells into myeloerythroid and myelolymphoid lineages. Cell Stem Cell 2007, 1, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, H.; Gregory, S.C.; Yokota, T.; Sakaguchi, N.; Kincade, P.W. Transcription from the RAG1 locus marks the earliest lymphocyte progenitors in bone marrow. Immunity 2002, 17, 117–130. [Google Scholar] [CrossRef]
- Semerad, C.L.; Mercer, E.M.; Inlay, M.A.; Weissman, I.L.; Murre, C. E2A proteins maintain the hematopoietic stem cell pool and promote the maturation of myelolymphoid and myeloerythroid progenitors. Proc. Natl. Acad. Sci. USA 2009, 106, 1930–1935. [Google Scholar] [CrossRef] [PubMed]
- Dias, S.; Mansson, R.; Gurbuxani, S.; Sigvardsson, M.; Kee, B.L. E2A proteins promote development of lymphoid-primed multipotent progenitors. Immunity 2008, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Miyai, T.; Takano, J.; Endo, T.A.; Kawakami, E.; Agata, Y.; Motomura, Y.; Kubo, M.; Kashima, Y.; Suzuki, Y.; Kawamoto, H.; et al. Three-step transcriptional priming that drives the commitment of multipotent progenitors toward B cells. Genes Dev. 2018, 32, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Ng, S.Y.; Zuniga-Pflucker, J.C.; Georgopoulos, K. Early hematopoietic lineage restrictions directed by Ikaros. Nat. Immunol. 2006, 7, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Carotta, S.; Dakic, A.; D’Amico, A.; Pang, S.H.; Greig, K.T.; Nutt, S.L.; Wu, L. The transcription factor PU.1 controls dendritic cell development and Flt3 cytokine receptor expression in a dose-dependent manner. Immunity 2010, 32, 628–641. [Google Scholar] [CrossRef] [PubMed]
- DeKoter, R.P.; Lee, H.J.; Singh, H. PU.1 regulates expression of the interleukin-7 receptor in lymphoid progenitors. Immunity 2002, 16, 297–309. [Google Scholar] [CrossRef]
- Greig, K.T.; de Graaf, C.A.; Murphy, J.M.; Carpinelli, M.R.; Pang, S.H.; Frampton, J.; Kile, B.T.; Hilton, D.J.; Nutt, S.L. Critical roles for c-Myb in lymphoid priming and early B-cell development. Blood 2010, 115, 2796–2805. [Google Scholar] [CrossRef] [PubMed]
- Spooner, C.J.; Cheng, J.X.; Pujadas, E.; Laslo, P.; Singh, H. A recurrent network involving the transcription factors PU.1 and Gfi1 orchestrates innate and adaptive immune cell fates. Immunity 2009, 31, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Welinder, E.; Mansson, R.; Mercer, E.M.; Bryder, D.; Sigvardsson, M.; Murre, C. The transcription factors E2A and HEB act in concert to induce the expression of FOXO1 in the common lymphoid progenitor. Proc. Natl. Acad. Sci. USA 2011, 108, 17402–17407. [Google Scholar] [CrossRef] [PubMed]
- Mansson, R.; Welinder, E.; Ahsberg, J.; Lin, Y.C.; Benner, C.; Glass, C.K.; Lucas, J.S.; Sigvardsson, M.; Murre, C. Positive intergenic feedback circuitry, involving EBF1 and FOXO1, orchestrates B-cell fate. Proc. Natl. Acad. Sci. USA 2012, 109, 21028–21033. [Google Scholar] [CrossRef] [PubMed]
- Zandi, S.; Mansson, R.; Tsapogas, P.; Zetterblad, J.; Bryder, D.; Sigvardsson, M. EBF1 is essential for B-lineage priming and establishment of a transcription factor network in common lymphoid progenitors. J. Immunol. 2008, 181, 3364–3372. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Grosschedl, R. Failure of B-cell differentiation in mice lacking the transcription factor EBF. Nature 1995, 376, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Jhunjhunwala, S.; Benner, C.; Heinz, S.; Welinder, E.; Mansson, R.; Sigvardsson, M.; Hagman, J.; Espinoza, C.A.; Dutkowski, J.; et al. A global network of transcription factors, involving E2A, EBF1 and Foxo1, that orchestrates B cell fate. Nat. Immunol. 2010, 11, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Maier, H.; Ostraat, R.; Gao, H.; Fields, S.; Shinton, S.A.; Medina, K.L.; Ikawa, T.; Murre, C.; Singh, H.; Hardy, R.R.; et al. Early B cell factor cooperates with Runx1 and mediates epigenetic changes associated with mb-1 transcription. Nat. Immunol. 2004, 5, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Cauchy, P.; Ramamoorthy, S.; Boller, S.; Chavez, L.; Grosschedl, R. Dynamic EBF1 occupancy directs sequential epigenetic and transcriptional events in B-cell programming. Genes Dev. 2018, 32, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Lukin, K.; Ramirez, J.; Fields, S.; Lopez, D.; Hagman, J. Opposing effects of SWI/SNF and Mi-2/NuRD chromatin remodeling complexes on epigenetic reprogramming by EBF and Pax5. Proc. Natl. Acad. Sci. USA 2009, 106, 11258–22363. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Y.; Ramamoorthy, S.; Boller, S.; Rosenbaum, M.; Gil, A.R.; Mittler, G.; Imai, Y.; Kuba, K.; Grosschedl, R. Interaction of CCR4-NOT with EBF1 regulates gene-specific transcription and mRNA stability in B lymphopoiesis. Genes Dev. 2016, 30, 2310–2324. [Google Scholar] [CrossRef] [PubMed]
- Gyory, I.; Boller, S.; Nechanitzky, R.; Mandel, E.; Pott, S.; Liu, E.; Grosschedl, R. Transcription factor Ebf1 regulates differentiation stage-specific signaling, proliferation, and survival of B cells. Genes Dev. 2012, 25, 668–682. [Google Scholar] [CrossRef] [PubMed]
- Tsapogas, P.; Zandi, S.; Ahsberg, J.; Zetterblad, J.; Welinder, E.; Jonsson, J.I.; Mansson, R.; Qian, H.; Sigvardsson, M. IL-7 mediates Ebf-1-dependent lineage restriction in early lymphoid progenitors. Blood 2011, 118, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Thal, M.A.; Carvalho, T.L.; He, T.; Kim, H.G.; Gao, H.; Hagman, J.; Klug, C.A. Ebf1-mediated down-regulation of Id2 and Id3 is essential for specification of the B cell lineage. Proc. Natl. Acad. Sci. USA 2009, 106, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Pongubala, J.M.; Northrup, D.L.; Lancki, D.W.; Medina, K.L.; Treiber, T.; Bertolino, E.; Thomas, M.; Grosschedl, R.; Allman, D.; Singh, H. Transcription factor EBF restricts alternative lineage options and promotes B cell fate commitment independently of Pax5. Nat. Immunol. 2008, 9, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Nechanitzky, R.; Akbas, D.; Scherer, S.; Gyory, I.; Hoyler, T.; Ramamoorthy, S.; Diefenbach, A.; Grosschedl, R. Transcription factor EBF1 is essential for the maintenance of B cell identity and prevention of alternative fates in committed cells. Nat. Immunol. 2013, 14, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Nutt, S.L.; Urbanek, P.; Rolink, A.; Busslinger, M. Essential functions of Pax5 (BSAP) in pro-B cell development: Difference between fetal and adult B lymphopoiesis and reduced V-to-DJ recombination at the IgH locus. Genes Dev. 1997, 11, 476–491. [Google Scholar] [CrossRef] [PubMed]
- Zandi, S.; Ahsberg, J.; Tsapogas, P.; Stjernberg, J.; Qian, H.; Sigvardsson, M. Single-cell analysis of early B-lymphocyte development suggests independent regulation of lineage specification and commitment in vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 15871–15876. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, I.; Heavey, B.; Horcher, M.; Busslinger, M. Reversion of B cell commitment upon loss of Pax5 expression. Science 2002, 297, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Nutt, S.L.; Heavey, B.; Rolink, A.G.; Busslinger, M. Commitment to the B-lymphoid lineage depends on the transcription factor Pax5. Nature 1999, 401, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Rolink, A.G.; Nutt, S.L.; Melchers, F.; Busslinger, M. Long-term in vivo reconstitution of T-cell development by Pax5- deficient B-cell progenitors. Nature 1999, 401, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Hoflinger, S.; Kesavan, K.; Fuxa, M.; Hutter, C.; Heavey, B.; Radtke, F.; Busslinger, M. Analysis of Notch1 function by in vitro T cell differentiation of Pax5 mutant lymphoid progenitors. J. Immunol. 2004, 173, 3935–3944. [Google Scholar] [CrossRef] [PubMed]
- Rolink, A.G.; Schaniel, C.; Bruno, L.; Melchers, F. In vitro and in vivo plasticity of Pax5-deficient pre-B I cells. Immunol. Lett. 2002, 82, 35–40. [Google Scholar] [CrossRef]
- Schaniel, C.; Bruno, L.; Melchers, F.; Rolink, A.G. Multiple hematopoietic cell lineages develop in vivo from transplanted Pax5-deficient pre-B I-cell clones. Blood 2002, 99, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Tagoh, H.; Ingram, R.; Wilson, N.; Salvagiotto, G.; Warren, A.J.; Clarke, D.; Busslinger, M.; Bonifer, C. The mechanism of repression of the myeloid-specific c-fms gene by Pax5 during B lineage restriction. EMBO J. 2006, 25, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Souabni, A.; Cobaleda, C.; Schebesta, M.; Busslinger, M. Pax5 promotes B lymphopoiesis and blocks T cell development by repressing Notch1. Immunity 2002, 17, 781–793. [Google Scholar] [CrossRef]
- Decker, T.; Pasca di Magliano, M.; McManus, S.; Sun, Q.; Bonifer, C.; Tagoh, H.; Busslinger, M. Stepwise activation of enhancer and promoter regions of the B cell commitment gene Pax5 in early lymphopoiesis. Immunity 2009, 30, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Roessler, S.; Gyory, I.; Imhof, S.; Spivakov, M.; Williams, R.R.; Busslinger, M.; Fisher, A.G.; Grosschedl, R. Distinct promoters mediate the regulation of Ebf1 gene expression by interleukin-7 and Pax5. Mol. Cell. Biol. 2007, 27, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.M.; Gisler, R.; Sigvardsson, M. Cloning and Characterization of a Promoter Flanking the Early B Cell Factor (EBF) Gene Indicates Roles for E-Proteins and Autoregulation in the Control of EBF Expression. J. Immunol. 2002, 169, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Ungerback, J.; Ahsberg, J.; Strid, T.; Somasundaram, R.; Sigvardsson, M. Combined heterozygous loss of Ebf1 and Pax5 allows for T-lineage conversion of B cell progenitors. J. Exp. Med. 2015, 212, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, R.; Ahsberg, J.; Okuyama, K.; Ungerback, J.; Lilljebjorn, H.; Fioretos, T.; Strid, T.; Sigvardsson, M. Clonal conversion of B lymphoid leukemia reveals cross-lineage transfer of malignant states. Genes Dev. 2016, 30, 2486–2499. [Google Scholar] [CrossRef] [PubMed]
- Treiber, T.; Mandel, E.M.; Pott, S.; Gyory, I.; Firner, S.; Liu, E.T.; Grosschedl, R. Early B Cell Factor 1 Regulates B Cell Gene Networks by Activation, Repression, and Transcription-Independent Poising of Chromatin. Immunity 2010, 32, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Revilla, I.D.R.; Bilic, I.; Vilagos, B.; Tagoh, H.; Ebert, A.; Tamir, I.M.; Smeenk, L.; Trupke, J.; Sommer, A.; Jaritz, M.; et al. The B-cell identity factor Pax5 regulates distinct transcriptional programmes in early and late B lymphopoiesis. EMBO J. 2012, 31, 3130–3146. [Google Scholar] [CrossRef] [PubMed]
- Nutt, S.L.; Thevenin, C.; Busslinger, M. Essential functions of Pax-5 (BSAP) in pro-B cell development. Immunobiology 1997, 198, 227–235. [Google Scholar] [CrossRef]
- Nutt, S.L.; Morrison, A.M.; Dorfler, P.; Rolink, A.; Busslinger, M. Identification of BSAP (Pax-5) target genes in early B-cell development by loss- and gain-of-function experiments. EMBO J. 1998, 17, 2319–2333. [Google Scholar] [CrossRef] [PubMed]
- Martensson, I.L.; Rolink, A.; Melchers, F.; Mundt, C.; Licence, S.; Shimizu, T. The pre-B cell receptor and its role in proliferation and Ig heavy chain allelic exclusion. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2002; Volume 14, pp. 335–342. [Google Scholar]
- Erlandsson, L.; Licence, S.; Gaspal, F.; Lane, P.; Corcoran, A.E.; Martensson, I.L. Both the pre-BCR and the IL-7Ralpha are essential for expansion at the pre-BII cell stage in vivo. Eur. J. Immunol. 2005, 35, 1969–1976. [Google Scholar] [CrossRef] [PubMed]
- Grawunder, U.; Leu, T.M.J.; Scatz, D.G.; Werner, A.; Rolink, A.G.; Melchers, F.; Winkler, T.H. Down-regulation of RAG1 and RAG2 gene expression in preB cells after functional immunoglobulin heavy chain rearrangement. Immunity 1995, 3, 601–608. [Google Scholar] [CrossRef]
- Mandal, M.; Powers, S.E.; Ochiai, K.; Georgopoulos, K.; Kee, B.L.; Singh, H.; Clark, M.R. Ras orchestrates exit from the cell cycle and light-chain recombination during early B cell development. Nat. Immunol. 2009, 10, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Hashimshony, T.; Sawai, C.M.; Pongubala, J.M.; Skok, J.A.; Aifantis, I.; Singh, H. Regulation of immunoglobulin light-chain recombination by the transcription factor IRF-4 and the attenuation of interleukin-7 signaling. Immunity 2008, 28, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, K.; Maienschein-Cline, M.; Mandal, M.; Triggs, J.R.; Bertolino, E.; Sciammas, R.; Dinner, A.R.; Clark, M.R.; Singh, H. A self-reinforcing regulatory network triggered by limiting IL-7 activates pre-BCR signaling and differentiation. Nat. Immunol. 2012, 13, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Pridans, C.; Holmes, M.L.; Polli, M.; Wettenhall, J.M.; Dakic, A.; Corcoran, L.M.; Smyth, G.K.; Nutt, S.L. Identification of Pax5 target genes in early B cell differentiation. J. Immunol. 2008, 180, 1719–1728. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.C.; Cobb, B.S.; Sabbattini, P.; Meixlsperger, S.; Parelho, V.; Liberg, D.; Taylor, B.; Dillon, N.; Georgopoulos, K.; Jumaa, H.; et al. Ikaros DNA-binding proteins as integral components of B cell developmental-stage-specific regulatory circuits. Immunity 2007, 26, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Medina, K.; Lancki, D.; Singh, H. IRF-4,8 orchestrate the pre-B-to-B transition in lymphocyte development. Genes Dev. 2003, 17, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Amin, R.H.; Schlissel, M.S. Foxo1 directly regulates the transcription of recombination-activating genes during B cell development. Nat. Immunol. 2008, 9, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Dengler, H.S.; Baracho, G.V.; Omori, S.A.; Bruckner, S.; Arden, K.C.; Castrillon, D.H.; DePinho, R.A.; Rickert, R.C. Distinct functions for the transcription factor Foxo1 at various stages of B cell differentiation. Nat. Immunol. 2008, 9, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Mullighan, C.G.; Goorha, S.; Radtke, I.; Miller, C.B.; Coustan-Smith, E.; Dalton, J.D.; Girtman, K.; Mathew, S.; Ma, J.; Pounds, S.B.; et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature 2007, 446, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Mullighan, C.G.; Miller, C.B.; Radtke, I.; Phillips, L.A.; Dalton, J.; Ma, J.; White, D.; Hughes, T.P.; le Beau, M.M.; Pui, C.H.; et al. BCR-ABL1 lymphoblastic leukaemia is characterized by the deletion of Ikaros. Nature 2008, 453, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, R.P.; Schoenmakers, E.F.; van Reijmersdal, S.V.; Hehir-Kwa, J.Y.; van Kessel, A.G.; van Leeuwen, F.N.; Hoogerbrugge, P.M. High-resolution genomic profiling of childhood ALL reveals novel recurrent genetic lesions affecting pathways involved in lymphocyte differentiation and cell cycle progression. Leukemia 2007, 21, 1258–1266. [Google Scholar] [CrossRef] [PubMed]
- Iida, S.; Rao, P.H.; Nallasivam, P.; Hibshoosh, H.; Butler, M.; Louie, D.C.; Dyomin, V.; Ohno, H.; Chaganti, R.S.; Dalla-Favera, R. The t(9;14)(p13;q32) chromosomal translocation associated with lymphoplasmacytoid lymphoma involves the PAX-5 gene. Blood 1996, 88, 4110–4117. [Google Scholar] [PubMed]
- Bouamar, H.; Abbas, S.; Lin, A.P.; Wang, L.; Jiang, D.; Holder, K.N.; Kinney, M.C.; Hunicke-Smith, S.; Aguiar, R.C. A capture-sequencing strategy identifies IRF8, EBF1, and APRIL as novel IGH fusion partners in B-cell lymphoma. Blood 2013, 122, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.G.; Morin, R.D.; Zhang, J.; Hirst, M.; Zhao, Y.; Su, X.; Chen, S.C.; Payne-Turner, D.; Churchman, M.L.; Harvey, R.C.; et al. Genetic alterations activating kinase and cytokine receptor signaling in high-risk acute lymphoblastic leukemia. Cancer Cell 2012, 22, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Coyaud, E.; Struski, S.; Prade, N.; Familiades, J.; Eichner, R.; Quelen, C.; Bousquet, M.; Mugneret, F.; Talmant, P.; Pages, M.P.; et al. Wide diversity of PAX5 alterations in B-ALL: A Groupe Francophone de Cytogenetique Hematologique study. Blood 2010, 115, 3089–3097. [Google Scholar] [CrossRef] [PubMed]
- Kamps, M.P.; Murre, C.; Sun, X.H.; Baltimore, D. A new homeobox gene contributes the DNA binding domain of the t(1;19) translocation protein in pre-B ALL. Cell 1990, 60, 547–555. [Google Scholar] [CrossRef]
- Inaba, T.; Roberts, W.M.; Shapiro, L.H.; Jolly, K.W.; Raimondi, S.C.; Smith, S.D.; Look, A.T. Fusion of the leucine zipper gene HLF to the E2A gene in human acute B-lineage leukemia. Science 1992, 257, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Fazio, G.; Palmi, C.; Rolink, A.; Biondi, A.; Cazzaniga, G. PAX5/TEL acts as a transcriptional repressor causing down-modulation of CD19, enhances migration to CXCL12, and confers survival advantage in pre-BI cells. Cancer Res. 2008, 68, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Schrader, K.A.; Waanders, E.; Timms, A.E.; Vijai, J.; Miething, C.; Wechsler, J.; Yang, J.; Hayes, J.; Klein, R.J.; et al. A recurrent germline PAX5 mutation confers susceptibility to pre-B cell acute lymphoblastic leukemia. Nat. Genet. 2013, 45, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- DeKoter, R.P.; Singh, H. Regulation of B lymphocyte and macrophage development by graded expression of PU.1. Science 2000, 288, 1439–1441. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, J.; Khaled, W.; Burke, S.; Li, P.; Chen, X.; Yang, W.; Jenkins, N.A.; Copeland, N.G.; Zhang, S.; et al. Bcl11a is essential for lymphoid development and negatively regulates p53. J. Exp. Med. 2012, 209, 2467–2483. [Google Scholar] [CrossRef] [PubMed]
- Ferreiros-Vidal, I.; Carroll, T.; Taylor, B.; Terry, A.; Liang, Z.; Bruno, L.; Dharmalingam, G.; Khadayate, S.; Cobb, B.S.; Smale, S.T.; et al. Genome-wide identification of Ikaros targets elucidates its contribution to mouse B-cell lineage specification and pre-B-cell differentiation. Blood 2013, 121, 1769–1782. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, M.; Grosschedl, R. Coordinate regulation of B cell differentiation by the transcription factors EBF and E2A. Immunity 1999, 11, 21–31. [Google Scholar] [CrossRef]
- Urbánek, P.; Wang, Z.-Q.; Fetka, I.; Wagner, E.F.; Busslinger, M. Complete block of early B cell differentiation and altered patterning of the posterior midbrain in mice lacking Pax5/BSAP. Cell 1994, 79, 901–912. [Google Scholar] [CrossRef]
- Ahsberg, J.; Ungerback, J.; Strid, T.; Welinder, E.; Stjernberg, J.; Larsson, M.; Qian, H.; Sigvardsson, M. Early B-cell Factor 1 regulates the expansion of B-cell progenitors in a dose dependent manner. J. Biol. Chem. 2013, 288, 33449–33461. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.N.; Chen, Z.; Braas, D.; Lee, J.W.; Xiao, G.; Geng, H.; Cosgun, K.N.; Hurtz, C.; Shojaee, S.; Cazzaniga, V.; et al. Metabolic gatekeeper function of B-lymphoid transcription factors. Nature 2017, 542, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Lukin, K.; Fields, S.; Lopez, D.; Cherrier, M.; Ternyak, K.; Ramirez, J.; Feeney, A.J.; Hagman, J. Compound haploinsufficiencies of Ebf1 and Runx1 genes impede B cell lineage progression. Proc. Natl. Acad. Sci. USA 2010, 107, 7869–7874. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.A.; Ungerback, J.; Ahsberg, J.; Somasundaram, R.; Strid, T.; Larsson, M.; Mansson, R.; de Paepe, A.; Lilljebjorn, H.; Fioretos, T.; et al. Ebf1 heterozygosity results in increased DNA damage in pro-B cells and their synergistic transformation by Pax5 haploinsufficiency. Blood 2015, 125, 4052–4059. [Google Scholar] [CrossRef] [PubMed]
- Jennings, C.D.; Foon, K.A. Recent advances in flow cytometry: Application to the diagnosis of hematologic malignancy. Blood 1997, 90, 2863–2892. [Google Scholar] [PubMed]
- Craig, F.E.; Foon, K.A. Flow cytometric immunophenotyping for hematologic neoplasms. Blood 2008, 111, 3941–3967. [Google Scholar] [CrossRef] [PubMed]
- Manola, K.N. Cytogenetic abnormalities in acute leukaemia of ambiguous lineage: An overview. Br. J. Haematol. 2013, 163, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Simmons, S.; Knoll, M.; Drewell, C.; Wolf, I.; Mollenkopf, H.J.; Bouquet, C.; Melchers, F. Biphenotypic B-lymphoid/myeloid cells expressing low levels of Pax5: Potential targets of BAL development. Blood 2012, 120, 3688–3698. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, E.; Nguyen, S.M.; Fountaine, T.J.; Welp, K.; Gryder, B.; Qin, H.; Yang, Y.; Chien, C.D.; Seif, A.E.; Lei, H.; et al. CD19 CAR immune pressure induces B-precursor acute lymphoblastic leukaemia lineage switch exposing inherent leukaemic plasticity. Nat. Commun. 2016, 7, 12320. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Allman, D.; Goldschmidt, M.H.; Atchison, M.L.; Monroe, J.G.; Thomas-Tikhonenko, A. Oscillation between B-lymphoid and myeloid lineages in Myc-induced hematopoietic tumors following spontaneous silencing/reactivation of the EBF/Pax5 pathway. Blood 2003, 101, 1950–1955. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Acosta, E.; Pelayo, R. Lineage switching in acute leukemias: A consequence of stem cell plasticity? Bone Marrow Res. 2012, 2012, 406796. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Geyer, M.B.; Brentjens, R.J. CD19-targeted CAR T-cell therapeutics for hematologic malignancies: Interpreting clinical outcomes to date. Blood 2016, 127, 3312–3320. [Google Scholar] [CrossRef] [PubMed]
- Gardner, R.; Wu, D.; Cherian, S.; Fang, M.; Hanafi, L.A.; Finney, O.; Smithers, H.; Jensen, M.C.; Riddell, S.R.; Maloney, D.G.; et al. Acquisition of a CD19-negative myeloid phenotype allows immune escape of MLL-rearranged B-ALL from CD19 CAR-T-cell therapy. Blood 2016, 127, 2406–2410. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.; Nocka, K.; Beier, D.R.; Chu, T.Y.; Buck, J.; Lahm, H.W.; Wellner, D.; Leder, P.; Besmer, P. The hematopoietic growth factor KL is encoded by the Sl locus and is the ligand of the c-kit receptor, the gene product of the W locus. Cell 1990, 63, 225–233. [Google Scholar] [CrossRef]
- Ikuta, K.; Weissman, I.L. Evidence that hematopoietic stem cells express mouse c-kit but do not depend on steel factor for their generation. Proc. Natl. Acad. Sci. USA 1992, 89, 1502–1506. [Google Scholar] [CrossRef] [PubMed]
- Akashi, K.; Traver, D.; Miyamoto, T.; Weissman, I.L. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 2000, 404, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Pronk, C.; Rossi, D.; Månsson, R.; Attema, J.; Norddahl, G.; Chan, C.; Sigvardsson, M.; Weissman, I.; Bryder, D. Elucidation of the phenotype, functional, and molecular topography of a myeloerythroid progenitor cell hierarchy. Cell Stem Cell 2007, 1, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Waskow, C.; Paul, S.; Haller, C.; Gassmann, M.; Rodewald, H.R. Viable c-Kit(W/W) mutants reveal pivotal role for c-kit in the maintenance of lymphopoiesis. Immunity 2002, 17, 277–288. [Google Scholar] [CrossRef]
- Miller, C.L.; Rebel, V.I.; Lemieux, M.E.; Helgason, C.D.; Lansdorp, P.M.; Eaves, C.J. Studies of W mutant mice provide evidence for alternate mechanisms capable of activating hematopoietic stem cells. Exp. Hematol. 1996, 24, 185–194. [Google Scholar] [PubMed]
- Sharma, Y.; Astle, C.M.; Harrison, D.E. Heterozygous kit mutants with little or no apparent anemia exhibit large defects in overall hematopoietic stem cell function. Exp. Hematol. 2007, 35, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Rolink, A.; Streb, M.; Nishikawa, S.; Melchers, F. The c-kit-encoded tyrosine kinase regulates the proliferation of early pre-B cells. Eur. J. Immunol. 1991, 21, 2609–2612. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, A.; Hsu, Y.M.; Day, R.B.; Schuettpelz, L.G.; Christopher, M.J.; Borgerding, J.N.; Nagasawa, T.; Link, D.C. CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 2013, 495, 227–230. [Google Scholar] [CrossRef] [PubMed]
- D’Apuzzo, M.; Rolink, A.; Loetscher, M.; Hoxie, J.A.; Clark-Lewis, I.; Melchers, F.; Baggiolini, M.; Moser, B. The chemokine SDF-1, stromal cell-derived factor 1, attracts early stage B cell precursors via the chemokine receptor CXCR4. Eur. J. Immunol. 1997, 27, 1788–1793. [Google Scholar] [CrossRef] [PubMed]
- Karsunky, H.; Inlay, M.A.; Serwold, T.; Bhattacharya, D.; Weissman, I.L. Flk2+ common lymphoid progenitors possess equivalent differentiation potential for the B and T lineages. Blood 2008, 111, 5562–5570. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.L.; Carotta, S.; Corcoran, L.M.; Nutt, S.L. Repression of Flt3 by Pax5 is crucial for B-cell lineage commitment. Genes Dev. 2006, 20, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Tsapogas, P.; Swee, L.K.; Nusser, A.; Nuber, N.; Kreuzaler, M.; Capoferri, G.; Rolink, H.; Ceredig, R.; Rolink, A. In vivo evidence for an instructive role of fms-like tyrosine kinase-3 (FLT3) ligand in hematopoietic development. Haematologica 2014, 99, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Sitnicka, E.; Bryder, D.; Theilgaard-Monch, K.; Buza-Vidas, N.; Adolfsson, J.; Jacobsen, S.E. Key role of flt3 ligand in regulation of the common lymphoid progenitor but not in maintenance of the hematopoietic stem cell pool. Immunity 2002, 17, 463–472. [Google Scholar] [CrossRef]
- Sitnicka, E.; Brakebusch, C.; Martensson, I.L.; Svensson, M.; Agace, W.W.; Sigvardsson, M.; Buza-Vidas, N.; Bryder, D.; Cilio, C.M.; Ahlenius, H.; et al. Complementary signaling through flt3 and interleukin-7 receptor alpha is indispensable for fetal and adult B cell genesis. J. Exp. Med. 2003, 198, 1495–1506. [Google Scholar] [CrossRef] [PubMed]
- Schlenner, S.M.; Madan, V.; Busch, K.; Tietz, A.; Laufle, C.; Costa, C.; Blum, C.; Fehling, H.J.; Rodewald, H.R. Fate mapping reveals separate origins of T cells and myeloid lineages in the thymus. Immunity 2010, 32, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Peschon, J.J.; Morrissey, P.J.; Grabstein, K.H.; Ramsdell, F.J.; Maraskovsky, E.; Gliniak, B.C.; Park, L.S.; Ziegler, S.F.; Williams, D.E.; Ware, C.B.; et al. Early lymphocyte expansion is severely impaired in interleukin 7 receptor-deficient mice. J. Exp. Med. 1994, 180, 1955–1960. [Google Scholar] [CrossRef] [PubMed]
- Von Freeden-Jeffry, U.; Vieira, P.; Lucian, L.A.; McNeil, T.; Burdach, S.E.; Murray, R. Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a nonredundant cytokine. J. Exp. Med. 1995, 181, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.P.; Izon, D.; DeMuth, W.; Gerstein, R.; Bhandoola, A.; Allman, D. The earliest step in B lineage differentiation from common lymphoid progenitors is critically dependent upon interleukin 7. J. Exp. Med. 2002, 196, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Park, L.S.; Martin, U.; Garka, K.; Gliniak, B.; Di Santo, J.P.; Muller, W.; Largaespada, D.A.; Copeland, N.G.; Jenkins, N.A.; Farr, A.G.; et al. Cloning of the murine thymic stromal lymphopoietin (TSLP) receptor: Formation of a functional heteromeric complex requires interleukin 7 receptor. J. Exp. Med. 2000, 192, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Chappaz, S.; Flueck, L.; Farr, A.G.; Rolink, A.G.; Finke, D. Increased TSLP availability restores T- and B-cell compartments in adult IL-7 deficient mice. Blood 2007, 110, 3862–3870. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Akashi, K.; Domen, J.; Sugamura, K.; Weissman, I.L. Bcl-2 rescues T lymphopoiesis, but not B or NK cell development, in common gamma chain-deficient mice. Immunity 1997, 7, 155–162. [Google Scholar] [CrossRef]
- Akashi, K.; Kondo, M.; von Freeden-Jeffry, U.; Murray, R.; Weissman, I.L. Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Cell 1997, 89, 1033–1041. [Google Scholar] [CrossRef]
- Van der Plas, D.C.; Smiers, F.; Pouwels, K.; Hoefsloot, L.H.; Lowenberg, B.; Touw, I.P. Interleukin-7 signaling in human B cell precursor acute lymphoblastic leukemia cells and murine BAF3 cells involves activation of STAT1 and STAT5 mediated via the interleukin-7 receptor alpha chain. Leukemia 1996, 10, 1317–1325. [Google Scholar] [PubMed]
- Dias, S.; Silva, H., Jr.; Cumano, A.; Vieira, P. Interleukin-7 is necessary to maintain the B cell potential in common lymphoid progenitors. J. Exp. Med. 2005, 201, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Kosan, C.; Saba, I.; Godmann, M.; Herold, S.; Herkert, B.; Eilers, M.; Moroy, T. Transcription factor miz-1 is required to regulate interleukin-7 receptor signaling at early commitment stages of B cell differentiation. Immunity 2010, 33, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Von Muenchow, L.; Alberti-Servera, L.; Klein, F.; Capoferri, G.; Finke, D.; Ceredig, R.; Rolink, A.; Tsapogas, P. Permissive roles of cytokines interleukin-7 and Flt3 ligand in mouse B-cell lineage commitment. Proc. Natl. Acad. Sci. USA 2016, 113, E8122–E8130. [Google Scholar] [CrossRef] [PubMed]
- Malin, S.; McManus, S.; Cobaleda, C.; Novatchkova, M.; Delogu, A.; Bouillet, P.; Strasser, A.; Busslinger, M. Role of STAT5 in controlling cell survival and immunoglobulin gene recombination during pro-B cell development. Nat. Immunol. 2010, 11, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Oliver, P.M.; Wang, M.; Zhu, Y.; White, J.; Kappler, J.; Marrack, P. Loss of Bim allows precursor B cell survival but not precursor B cell differentiation in the absence of interleukin 7. J. Exp. Med. 2004, 200, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Pribyl, J.; LeBien, T.W. Interleukin 7 independent development of human B cells. Proc. Natl. Acad. Sci. USA 1996, 93, 10348–10353. [Google Scholar]
- Puel, A.; Ziegler, S.F.; Buckley, R.H.; Leonard, W.J. Defective IL7R expression in T−B+NK+ severe combined immunodeficiency. Nat. Genet. 1998, 20, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Shochat, C.; Tal, N.; Bandapalli, O.R.; Palmi, C.; Ganmore, I.; te Kronnie, G.; Cario, G.; Cazzaniga, G.; Kulozik, A.E.; Stanulla, M.; et al. Gain-of-function mutations in interleukin-7 receptor-α (IL7R) in childhood acute lymphoblastic leukemias. J. Exp. Med. 2011, 208, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Heltemes-Harris, L.M.; Willette, M.J.; Ramsey, L.B.; Qiu, Y.H.; Neeley, E.S.; Zhang, N.; Thomas, D.A.; Koeuth, T.; Baechler, E.C.; Kornblau, S.M.; et al. Ebf1 or Pax5 haploinsufficiency synergizes with STAT5 activation to initiate acute lymphoblastic leukemia. J. Exp. Med. 2011, 208, 1135–1149. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sigvardsson, M. Molecular Regulation of Differentiation in Early B-Lymphocyte Development. Int. J. Mol. Sci. 2018, 19, 1928. https://doi.org/10.3390/ijms19071928
Sigvardsson M. Molecular Regulation of Differentiation in Early B-Lymphocyte Development. International Journal of Molecular Sciences. 2018; 19(7):1928. https://doi.org/10.3390/ijms19071928
Chicago/Turabian StyleSigvardsson, Mikael. 2018. "Molecular Regulation of Differentiation in Early B-Lymphocyte Development" International Journal of Molecular Sciences 19, no. 7: 1928. https://doi.org/10.3390/ijms19071928
APA StyleSigvardsson, M. (2018). Molecular Regulation of Differentiation in Early B-Lymphocyte Development. International Journal of Molecular Sciences, 19(7), 1928. https://doi.org/10.3390/ijms19071928