Co-Culturing Human Adipose Derived Stem Cells and Schwann Cells on Spider Silk—A New Approach as Prerequisite for Enhanced Nerve Regeneration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
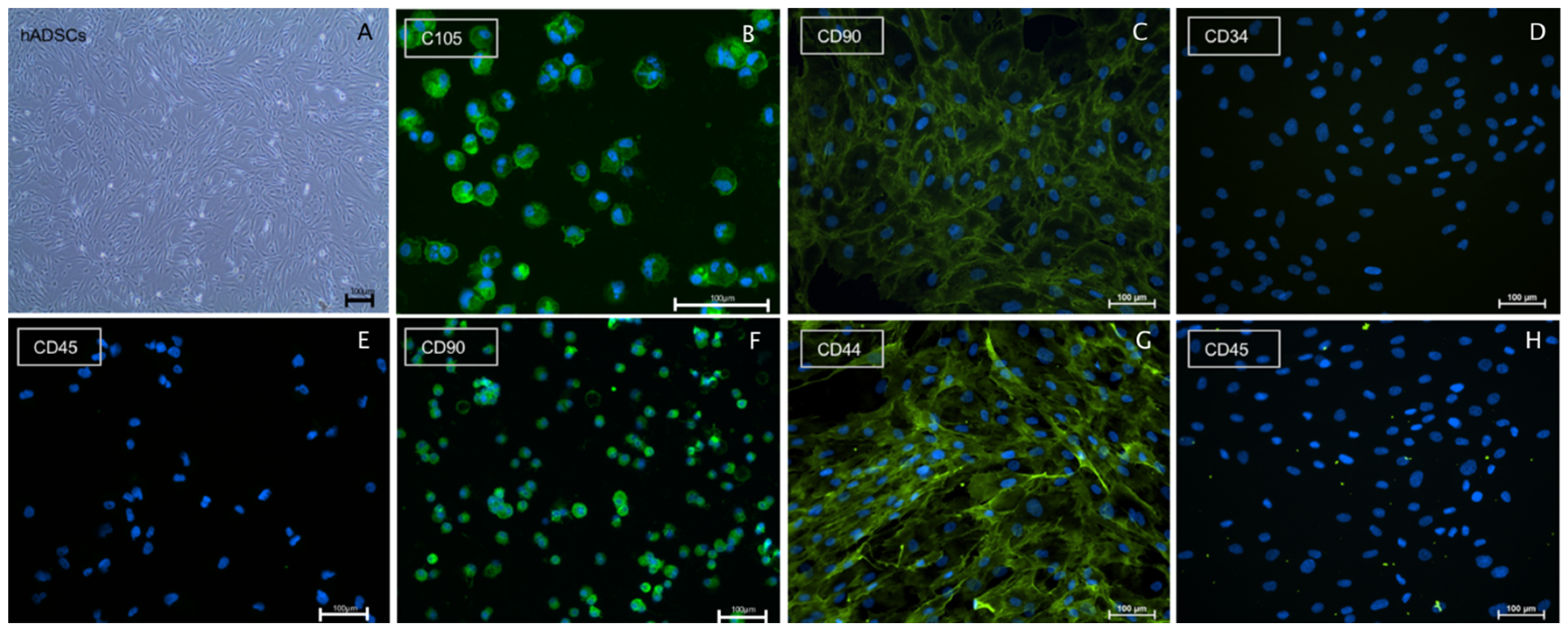
2.1. Immunostaining Confirmed Human Schwann Cell and Human ADSC Identity
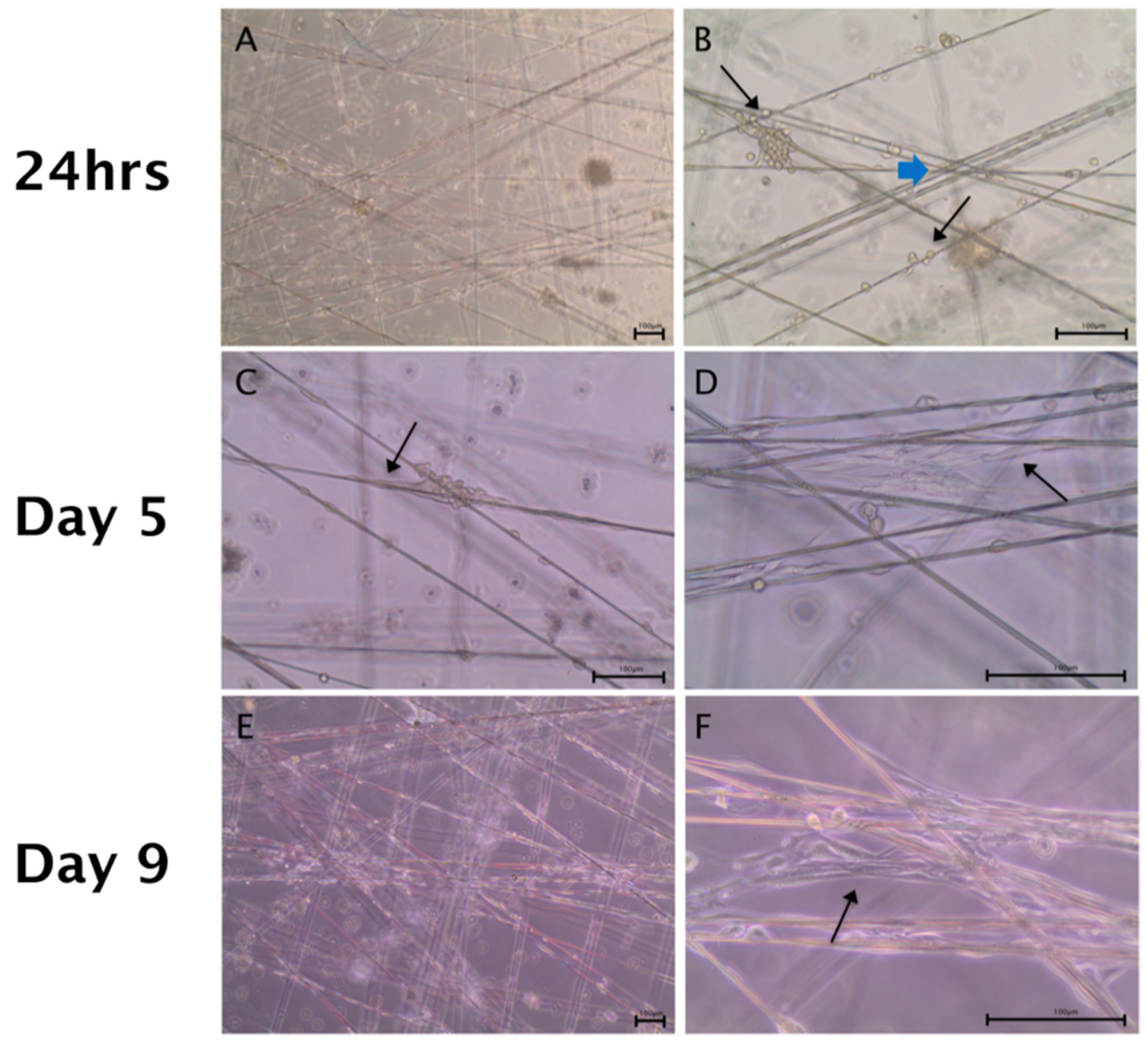
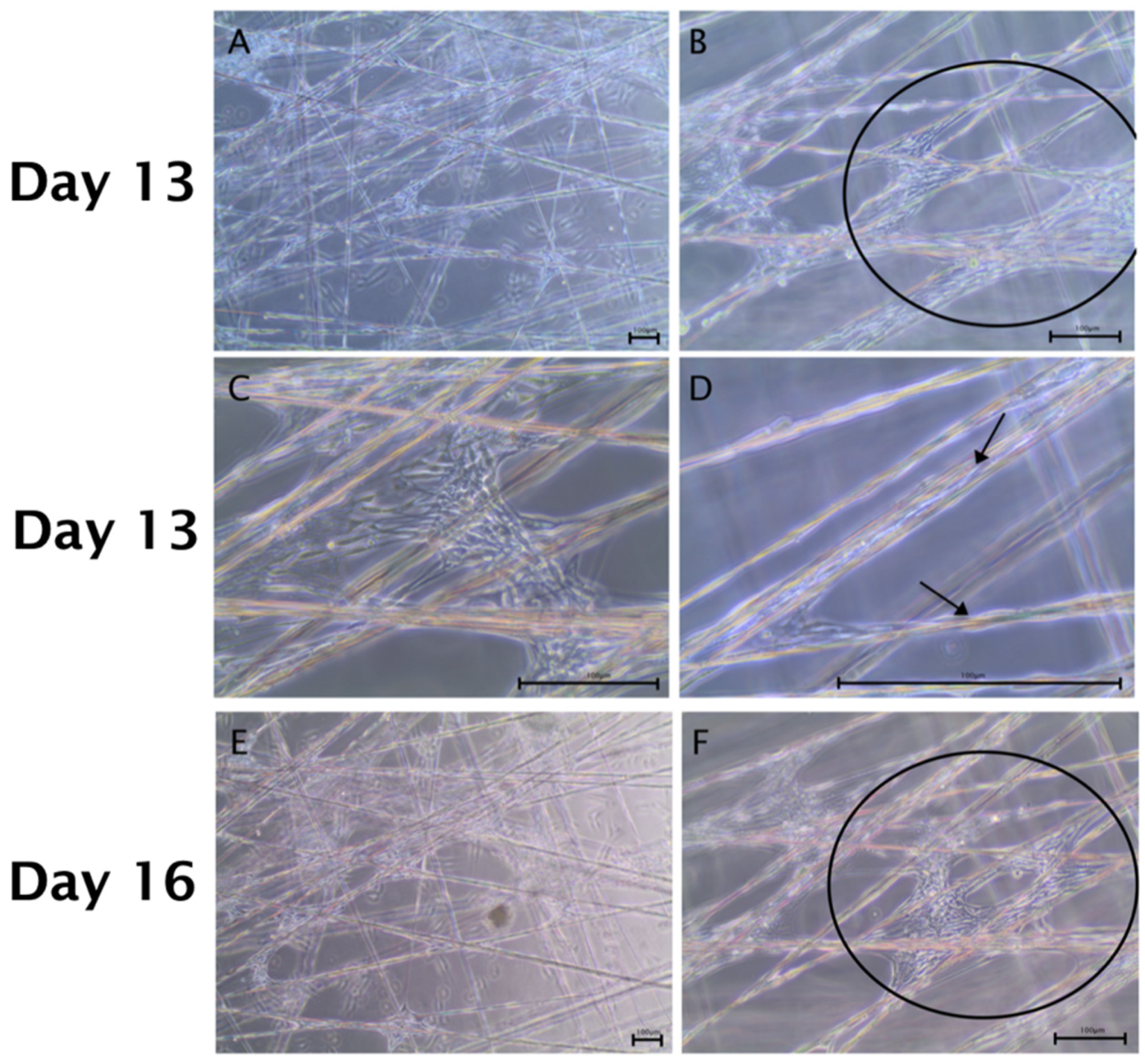
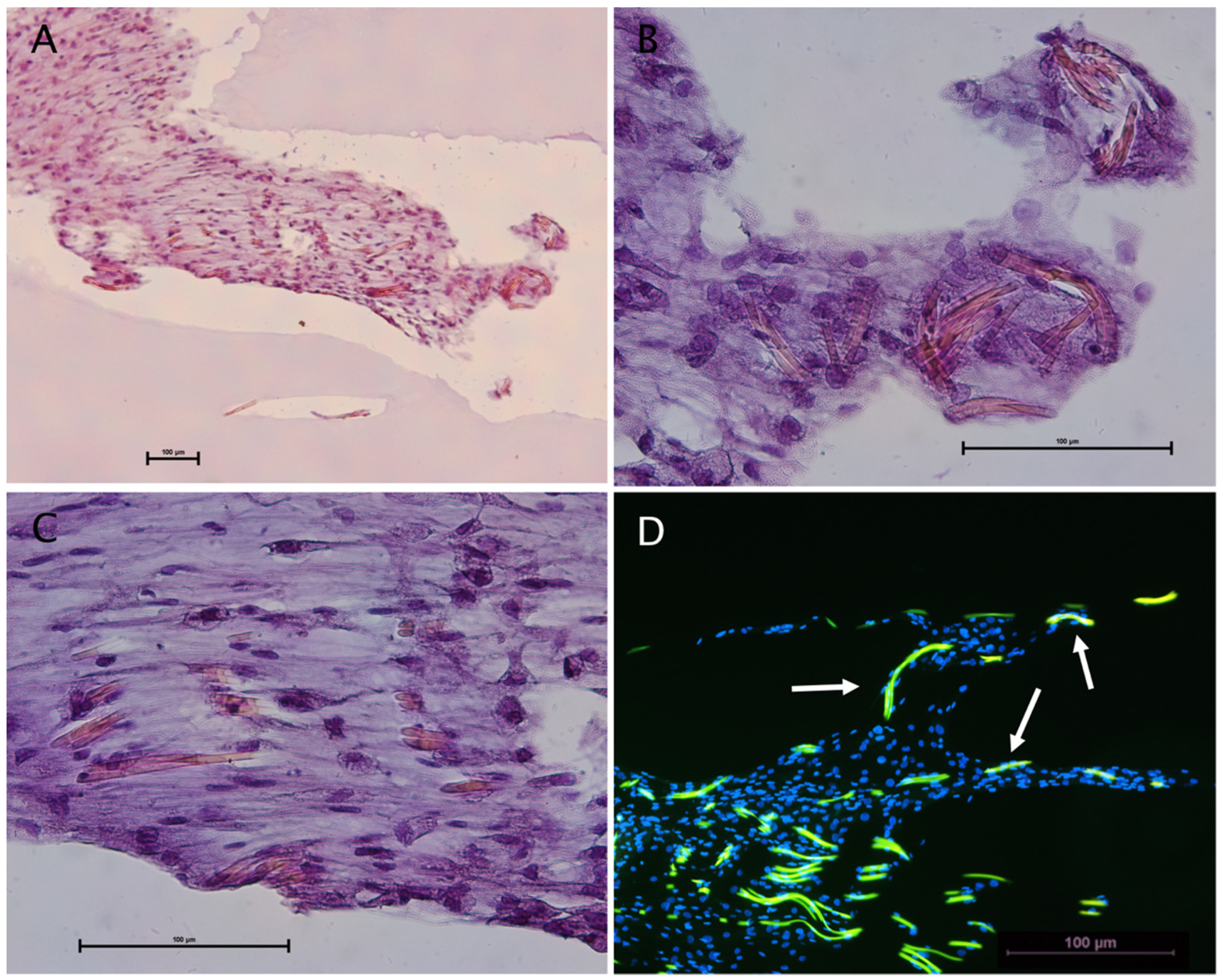
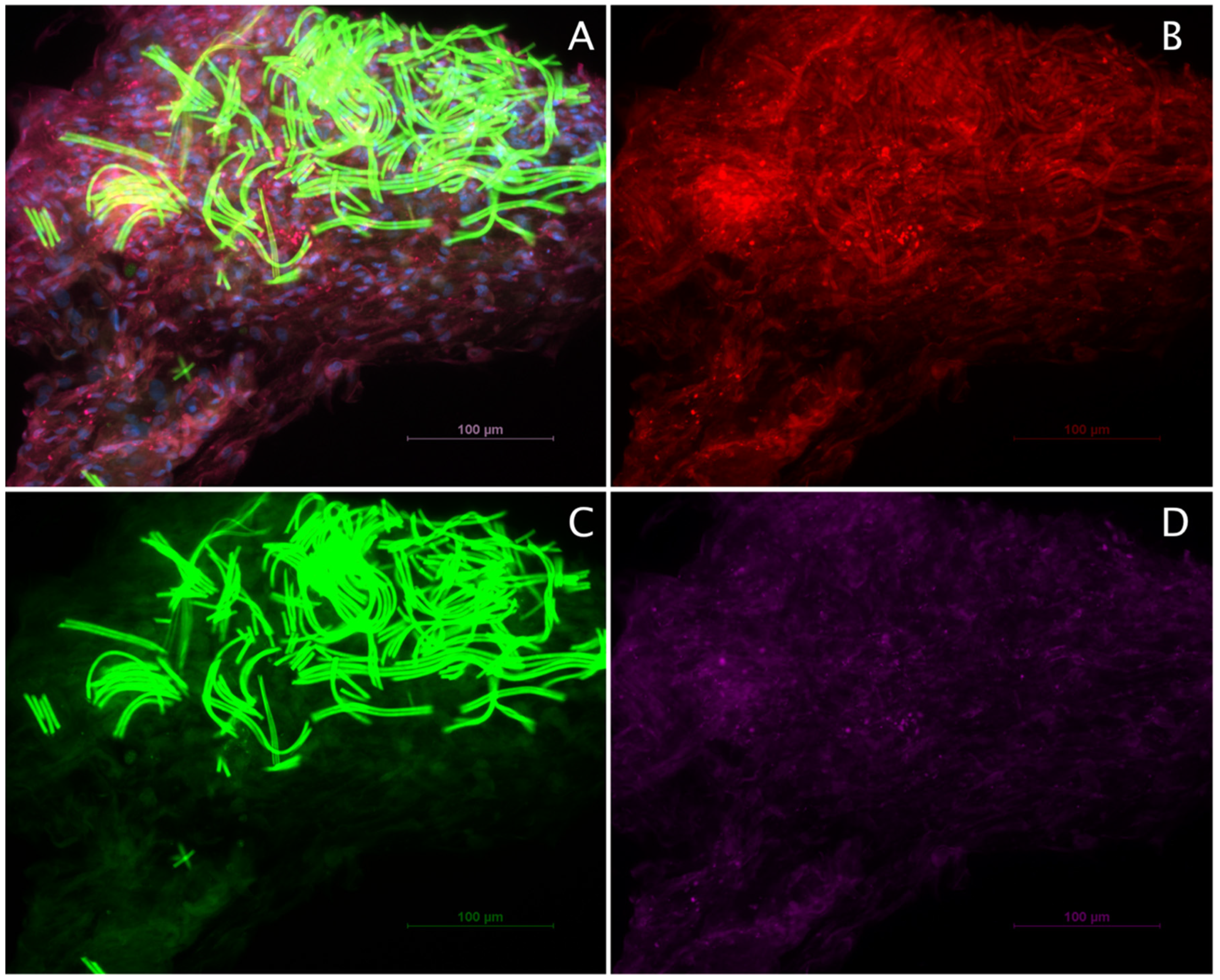
2.2. Spider Silk Enables Successful Adherence, Migration and Cell Expansion of Co-Cultures Composed of Human Schwann Cells and ADSCs
3. Discussion
4. Materials and Methods
4.1. Isolation of Human ADSCs
4.2. Isolation of Human Schwann Cells
4.3. Poly-l-Lysin/Laminin Coating
4.4. Harvesting Spider Silk
4.5. Seeding Co-Culture on Spider Silk
4.6. Cytospin Method
4.7. Immunofluorescence Staining
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADSC | adipose-derived stem cell |
| DAPI | 4′,6-Diamidin-2-phenylindol |
| FGF | Fibroblast growth factor |
| PBS | phosphate buffer solution |
| DMEM | Dulbecco’s Modified Eagle Medium |
| PLL | Poly-l-Lysin |
| hSCEM | human Schwann cell expansion medium |
References
- Chen, M.B.; Zhang, F.; Lineaweaver, W.C. Luminal fillers in nerve conduits for peripheral nerve repair. Ann. Plast. Surg. 2006, 57, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Madduri, S.; Papaloïzos, M.; Gander, B. Trophically and topographically functionalized silk fibroin nerve conduits for guided peripheral nerve regeneration. Biomaterials 2010, 31, 2323–2334. [Google Scholar] [CrossRef] [PubMed]
- Brohlin, M.; Mahay, D.; Novikov, L.N.; Terenghi, G.; Wiberg, M.; Shawcross, S.G.; Novikova, L.N. Characterisation of human mesenchymal stem cells following differentiation into Schwann cell-like cells. Neurosci. Res. 2009, 64, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Radtke, C.; Allmeling, C.; Waldmann, K.H.; Reimers, K.; Thies, K.; Schenk, H.C.; Hillmer, A.; Guggenheim, M.; Brandes, G.; Vogt, P.M. Spider silk constructs enhance axonal regeneration and remyelination in long nerve defects in sheep. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Kingham, P.J.; Kalbermatten, D.F.; Mahay, D.; Armstrong, S.J.; Wiberg, M.; Terenghi, G. Adipose-derived stem cells differentiate into a Schwann cell phenotype and promote neurite outgrowth in vitro. Exp. Neurol. 2007, 207, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Allmeling, C.; Jokuszies, A.; Reimers, K.; Kall, S.; Vogt, P.M. Use of spider silk fibres as an innovative material in a biocompatible artificial nerve conduit. J. Cell. Mol. Med. 2006, 10, 770–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allmeling, C.; Jokuszies, A.; Reimers, K.; Kall, S.; Choi, C.Y.; Brandes, G.; Kasper, C.; Scheper, T.; Guggenheim, M.; Vogt, P.M. Spider silk fibres in artificial nerve constructs promote peripheral nerve regeneration. Cell Prolif. 2008, 41, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Roloff, F.; Strauß, S.; Vogt, P.M.; Bicker, G.; Radtke, C. Spider silk as guiding biomaterial for human model neurons. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Kornfeld, T.; Vogt, P.M.; Bucan, V.; Peck, C.T.; Reimers, K.; Radtke, C. Characterization and Schwann cell seeding of up to 15.0 cm long spider silk nerve conduits for reconstruction of peripheral nerve defects. J. Funct. Biomater. 2016, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Radtke, C. Natural occurring silks and their analogues as materials for nerve conduits. Int. J. Mol. Sci. 2016, 17, 1754. [Google Scholar] [CrossRef]
- Di Summa, P.G.; Kingham, P.J.; Raffoul, W.; Wiberg, M.; Terenghi, G.; Kalbermatten, D.F. Adipose-derived stem cells enhance peripheral nerve regeneration. J. Plast. Reconstr. Aesthetic Surg. 2010, 63, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Faroni, A.A.; Rothwell, S.S.; Grolla, A.A.; Terenghi, G.G.; Magnaghi, V.V.; Verkhratsky, A.A. Differentiation of adipose-derived stem cells into Schwann cell phenotype induces expression of P2X receptors that control cell death. Cell Death Dis. 2013, 4, e743. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.G.; Huang, G.S.; Hsu, S.H. Sciatic nerve regeneration by cocultured schwann cells and stem cells on microporous nerve conduits. Cell Transplant. 2013, 22, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R. The repair Schwann cell and its function in regenerating nerves. J. Physiol. 2016, 594, 3521–3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Gong, K.; Zheng, Z.; Liu, L.; Wang, A.; Zhang, L.; Ao, Q.; Gong, Y.; Zhang, X. Schwann-like cell differentiation of rat adipose-derived stem cells by indirect co-culture with Schwann cells in vitro. Cell Prolif. 2010, 43, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Madura, T.; Mantovani, C.; Terenghi, G. Differentiated adipose-derived stem cells promote myelination and enhance functional recovery in a rat model of chronic denervation. J. Neurosci. Res. 2012, 90, 1392–1402. [Google Scholar] [CrossRef]
- Baksh, D.; Song, L.; Tuan, R.S. Adult mesenchymal stem cells: Characterization, differentiation, and application in cell and gene therapy. J. Cell. Mol. Med. 2004, 8, 301–316. [Google Scholar] [CrossRef]
- Gao, S.; Zheng, Y.; Cai, Q.; Wu, X.; Yao, W.; Wang, J. Different methods for inducing adipose-derived stem cells to differentiate into Schwann-like cells. Arch. Med. Sci. 2015, 11, 886–892. [Google Scholar] [CrossRef]
- Anghileri, E.; Marconi, S.; Pignatelli, A.; Cifelli, P.; Galie, M.; Sbarbati, A.; Krampera, M.; Belluzzi, O.; Bonetti, B. Neuronal Differentiation Potential of Human Adipose-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2008, 17, 909–916. [Google Scholar] [CrossRef]
- Wang, J.; Liao, L.; Tan, J. Mesenchymal-stem-cell-based experimental and clinical trials: Current status and open questions. Expert Opin. Biol. Ther. 2011, 11, 893–909. [Google Scholar] [CrossRef]
- Kaewkhaw, R.; Scutt, A.M.; Haycock, J.W. Anatomical site influences the differentiation of adipose-derived stem cells for Schwann-cell phenotype and function. Glia 2011, 59, 734–749. [Google Scholar] [CrossRef]
- Park, H.-W.; Lim, M.-J.; Jung, H.; Lee, S.-P.; Paik, K.-S.; Chang, M.-S. Human mesenchymal stem cell-derived Schwann cell-like cells exhibit neurotrophic effects, via distinct growth factor production, in a model of spinal cord injury. Glia 2010, 58, 1118–1132. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Qin, Y.; Wu, H.; Lv, Q.; Chen, X.; Deng, W. A new method for Schwann-like cell differentiation of adipose derived stem cells. Neurosci. Lett. 2013, 551, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Erba, P.; Mantovani, C.; Kalbermatten, D.F.; Pierer, G.; Terenghi, G.; Kingham, P.J. Regeneration potential and survival of transplanted undifferentiated adipose tissue-derived stem cells in peripheral nerve conduits. J. Plast. Reconstr. Aesthetic Surg. 2010, 63, e811–e817. [Google Scholar] [CrossRef] [PubMed]
- Kuhbier, J.W.; Allmeling, C.; Reimers, K.; Hillmer, A.; Kasper, C.; Menger, B.; Brandes, G.; Guggenheim, M.; Vogt, P.M. Interactions between spider silk and cells—NIH/3T3 fibroblasts seeded on miniature weaving frames. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Barcelos, A.S.; Rodrigues, A.C.; Silva, M.D.P.; Padovani, C.R. Inside-out vein graft and inside-out artery graft in rat sciatic nerve repair. Microsurgery 2003, 23, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Hall, S. Axonal regeneration through acellular muscle grafts. J. Anat. 1997, 190, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Protocols, D.; Weiss, T.; Taschner-Mandl, S.; Ambros, P.F.; Ambros, I.M. Schwann Cells; Springer: Berlin, Germany, 2018; Volume 1739, pp. 67–86. [Google Scholar]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Resch, A.; Wolf, S.; Mann, A.; Weiss, T.; Stetco, A.-L.; Radtke, C. Co-Culturing Human Adipose Derived Stem Cells and Schwann Cells on Spider Silk—A New Approach as Prerequisite for Enhanced Nerve Regeneration. Int. J. Mol. Sci. 2019, 20, 71. https://doi.org/10.3390/ijms20010071
Resch A, Wolf S, Mann A, Weiss T, Stetco A-L, Radtke C. Co-Culturing Human Adipose Derived Stem Cells and Schwann Cells on Spider Silk—A New Approach as Prerequisite for Enhanced Nerve Regeneration. International Journal of Molecular Sciences. 2019; 20(1):71. https://doi.org/10.3390/ijms20010071
Chicago/Turabian StyleResch, Annika, Sonja Wolf, Anda Mann, Tamara Weiss, Alexandra-Larissa Stetco, and Christine Radtke. 2019. "Co-Culturing Human Adipose Derived Stem Cells and Schwann Cells on Spider Silk—A New Approach as Prerequisite for Enhanced Nerve Regeneration" International Journal of Molecular Sciences 20, no. 1: 71. https://doi.org/10.3390/ijms20010071
APA StyleResch, A., Wolf, S., Mann, A., Weiss, T., Stetco, A.-L., & Radtke, C. (2019). Co-Culturing Human Adipose Derived Stem Cells and Schwann Cells on Spider Silk—A New Approach as Prerequisite for Enhanced Nerve Regeneration. International Journal of Molecular Sciences, 20(1), 71. https://doi.org/10.3390/ijms20010071