Insight of the Interaction between 2,4-thiazolidinedione and Human Serum Albumin: A Spectroscopic, Thermodynamic and Molecular Docking Study

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
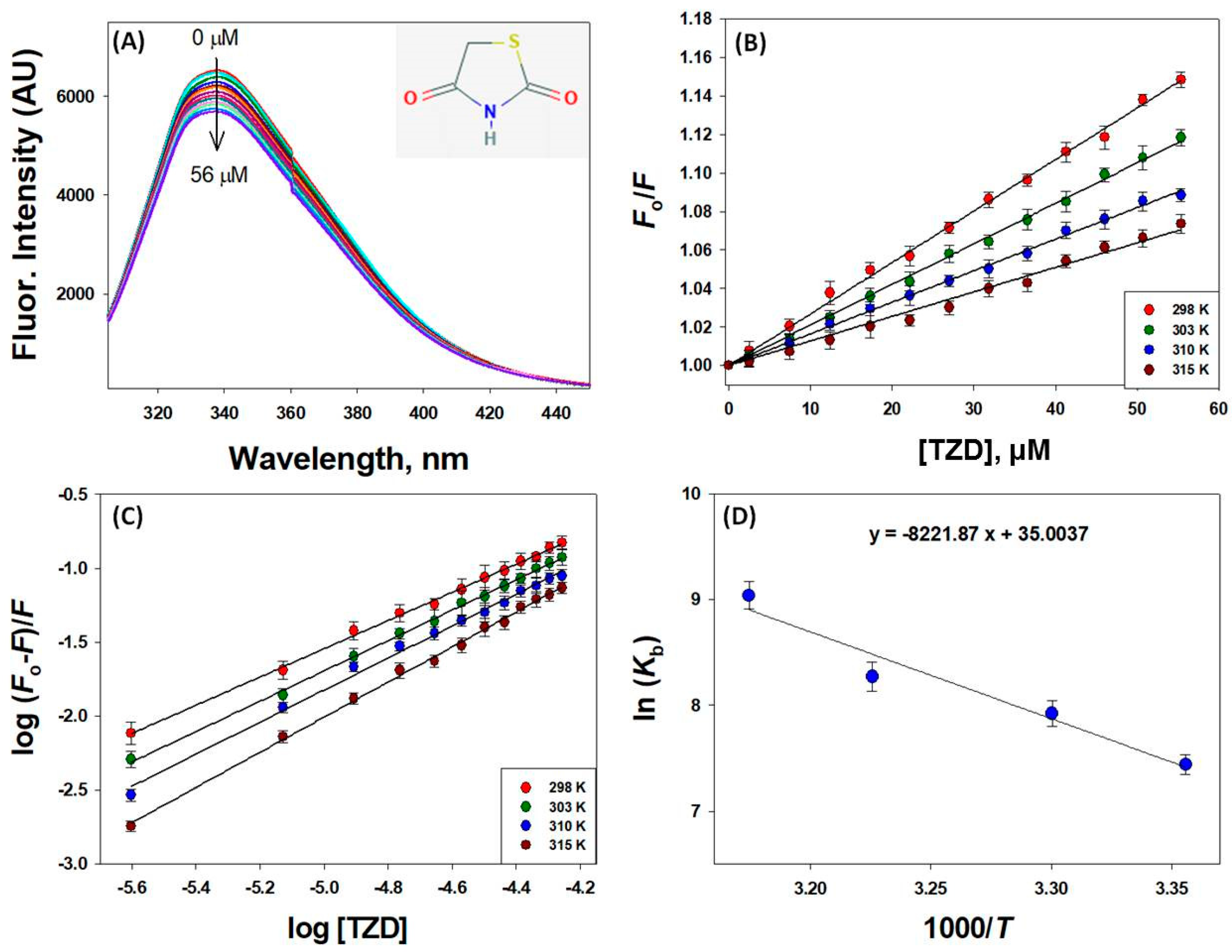
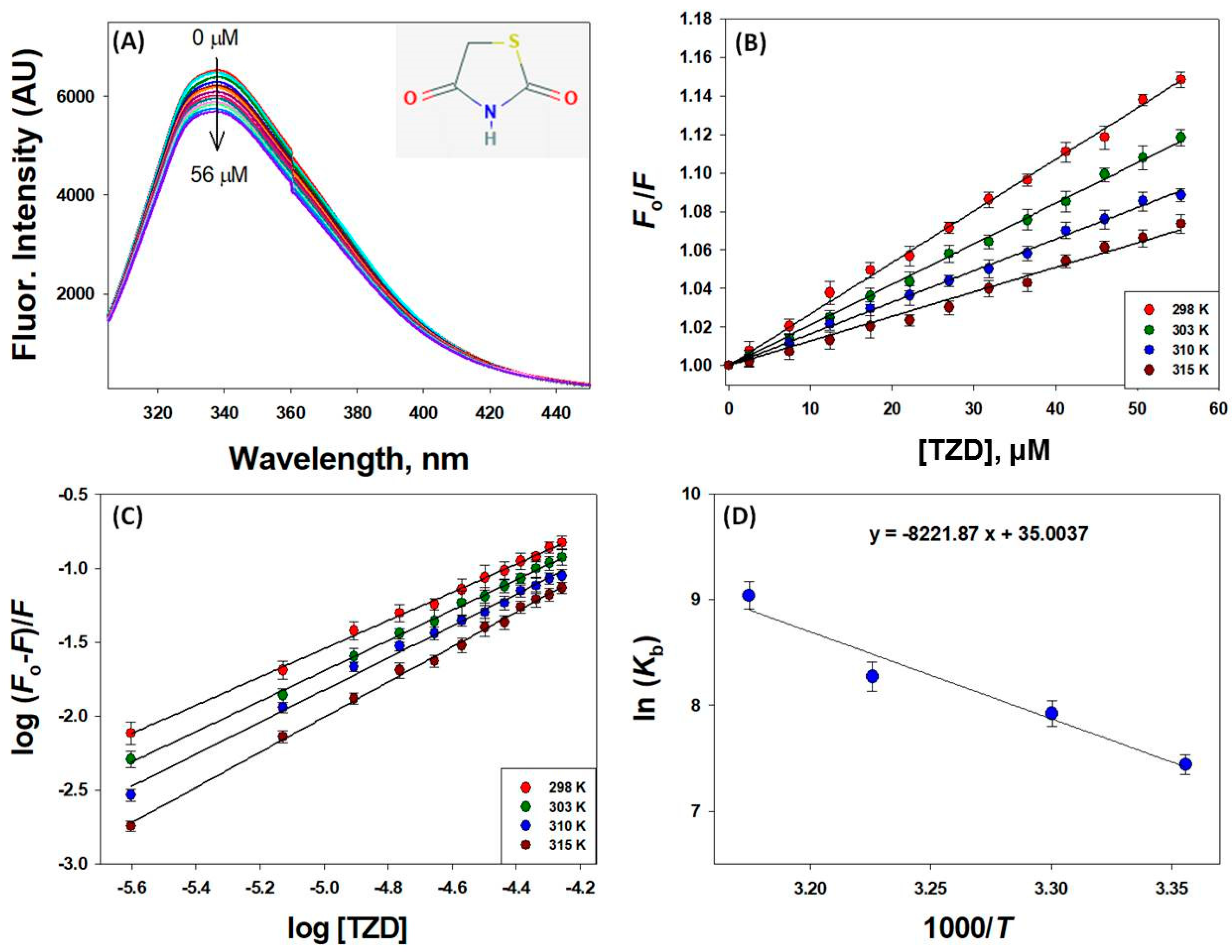
2.1. Characterization of 2,4-TZD Binding Sites on HSA
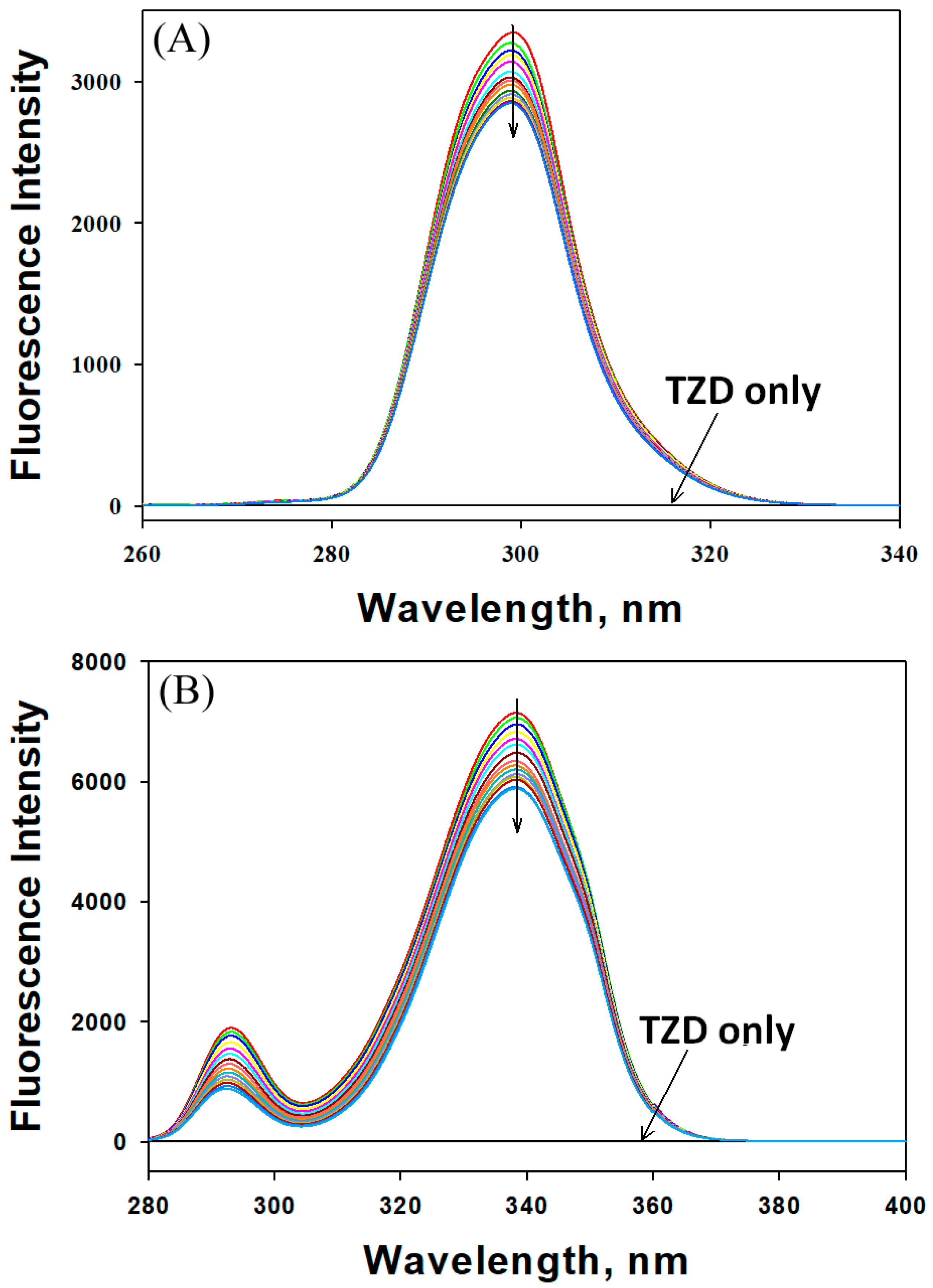
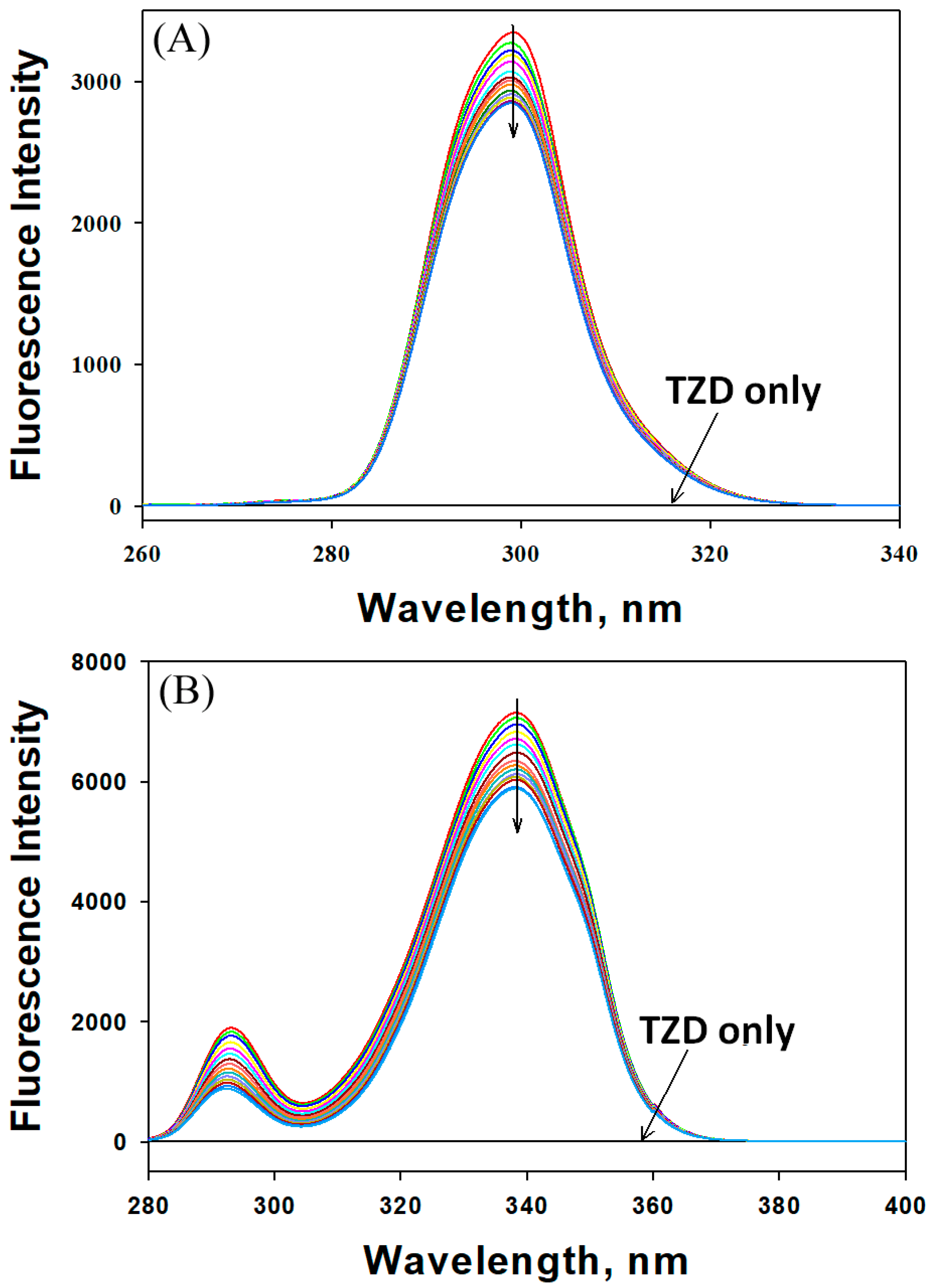
2.1.1. Mechanism of HSA Fluorescence Quenching by 2,4-TZD
2.1.2. Binding and Thermodynamics of 2,4-TZD/HSA Binding
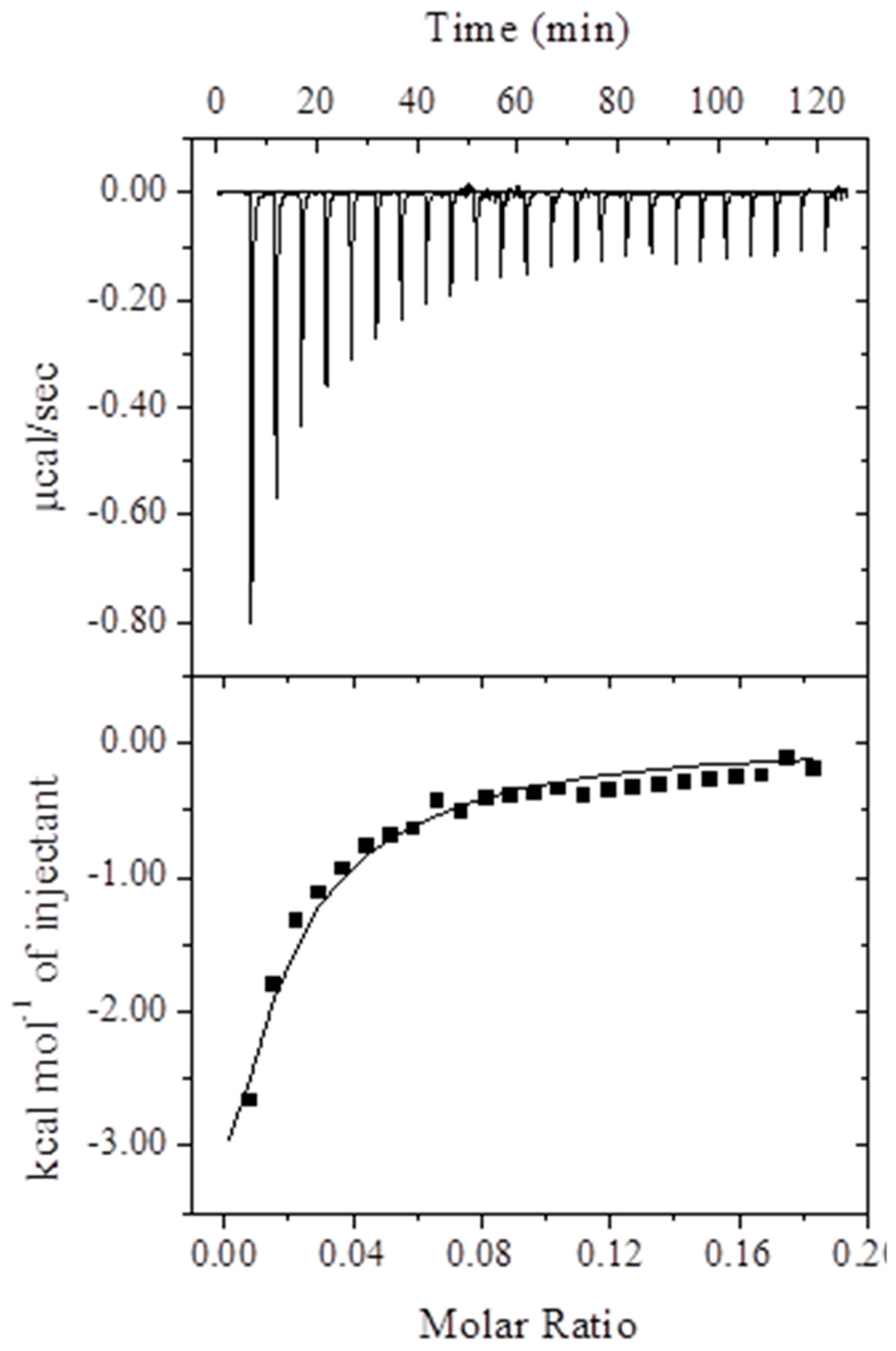
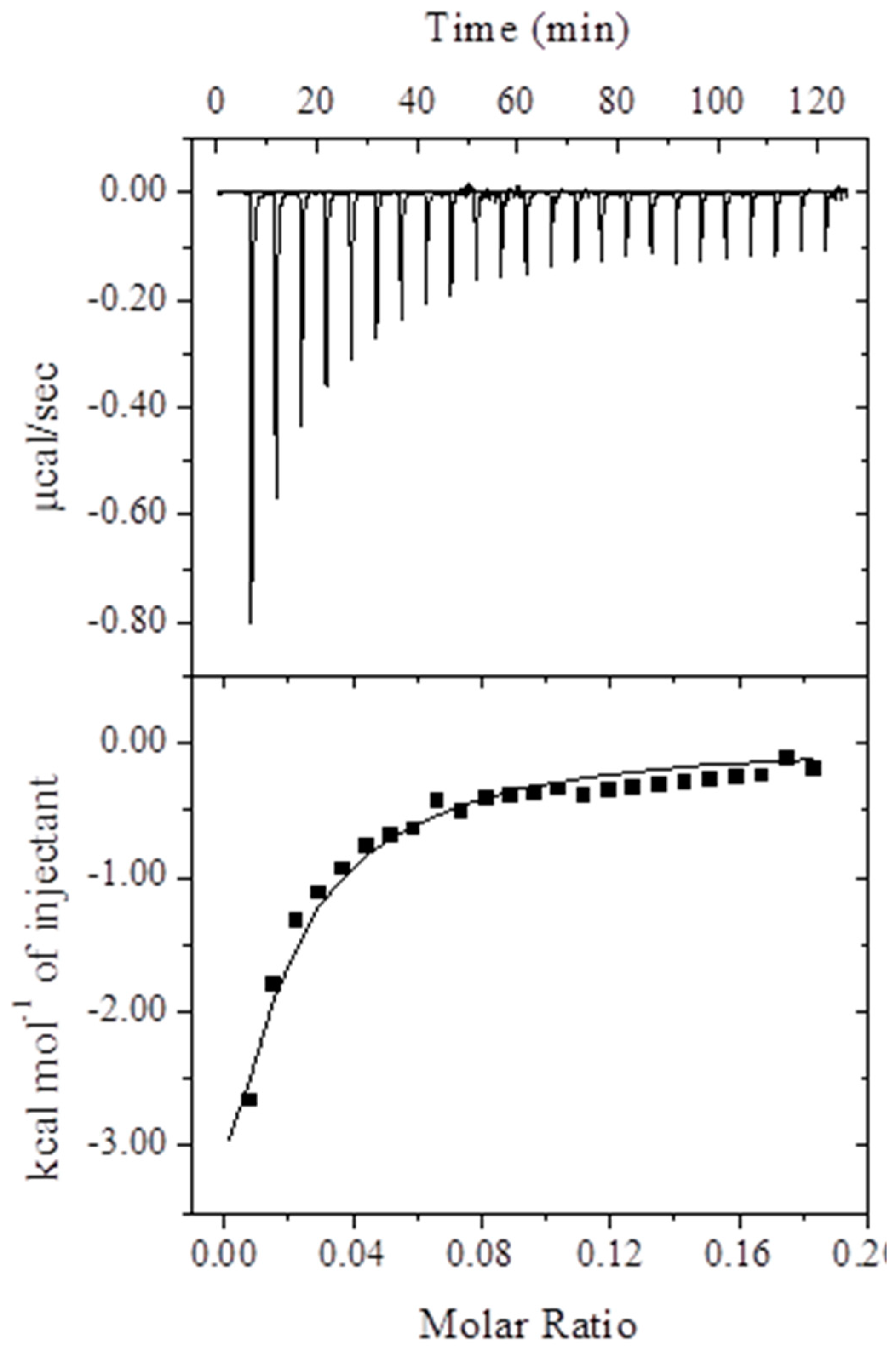
2.1.3. Isothermal Titration Calorimetry (ITC) Measurements
2.1.4. Förster Resonance Energy Transfer (FRET) between HSA and 2,4-TZD
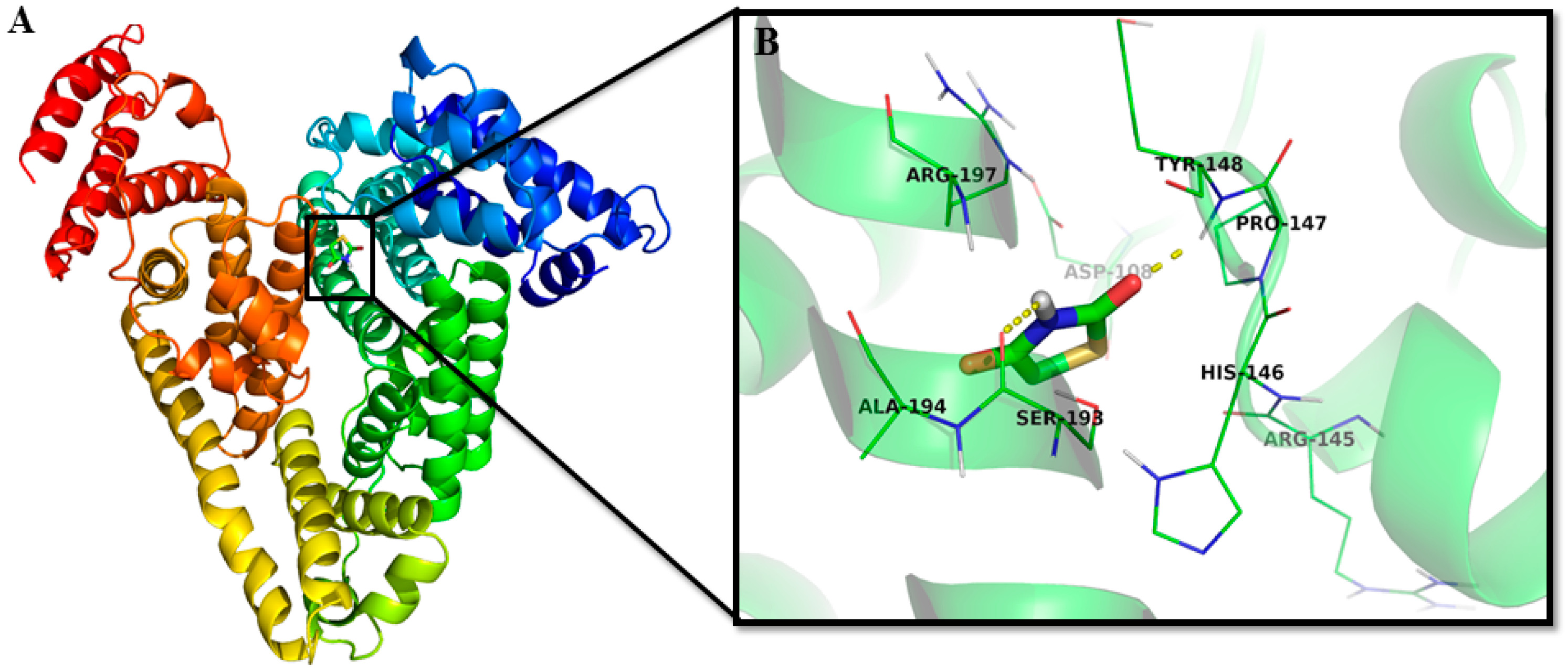
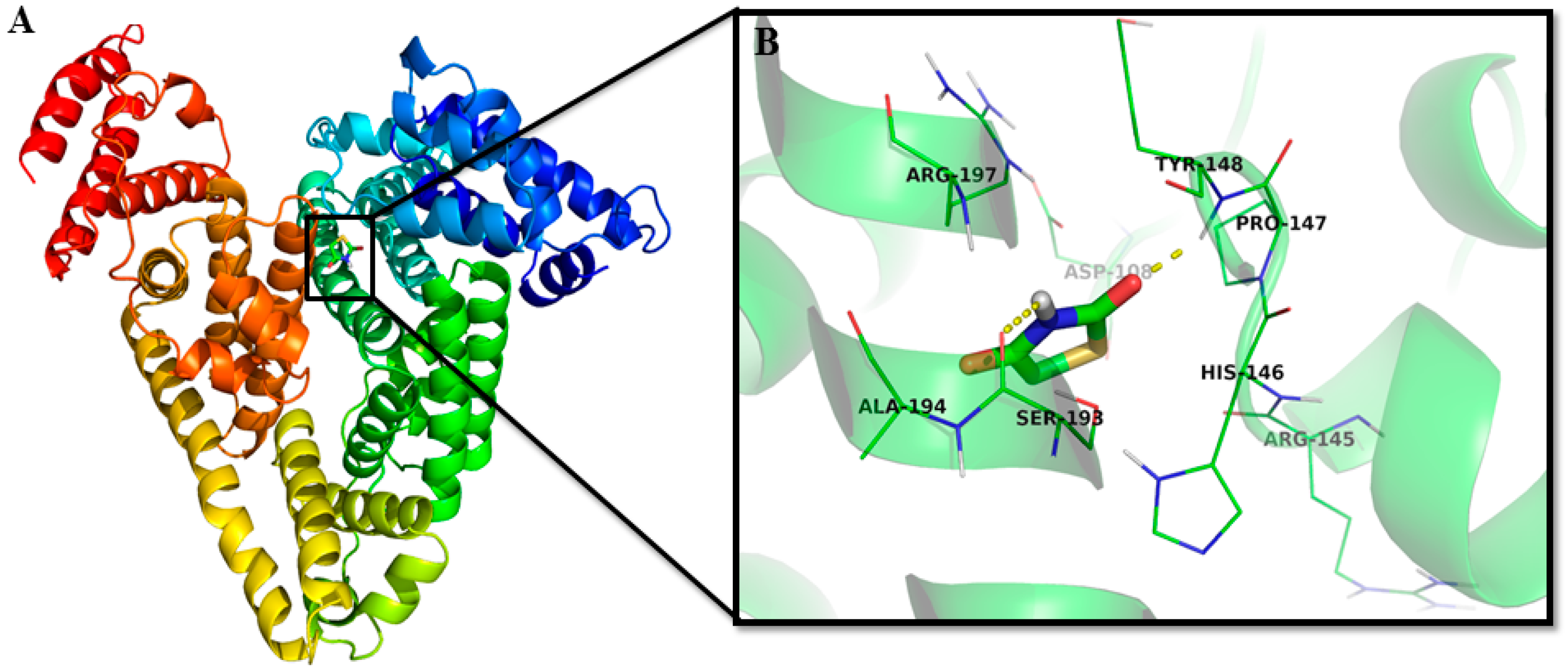
2.1.5. Location of the 2,4-TZD Binding Site on HSA by Molecular Docking
2.2. Effect of 2,4-TZD on the Conformation of HSA
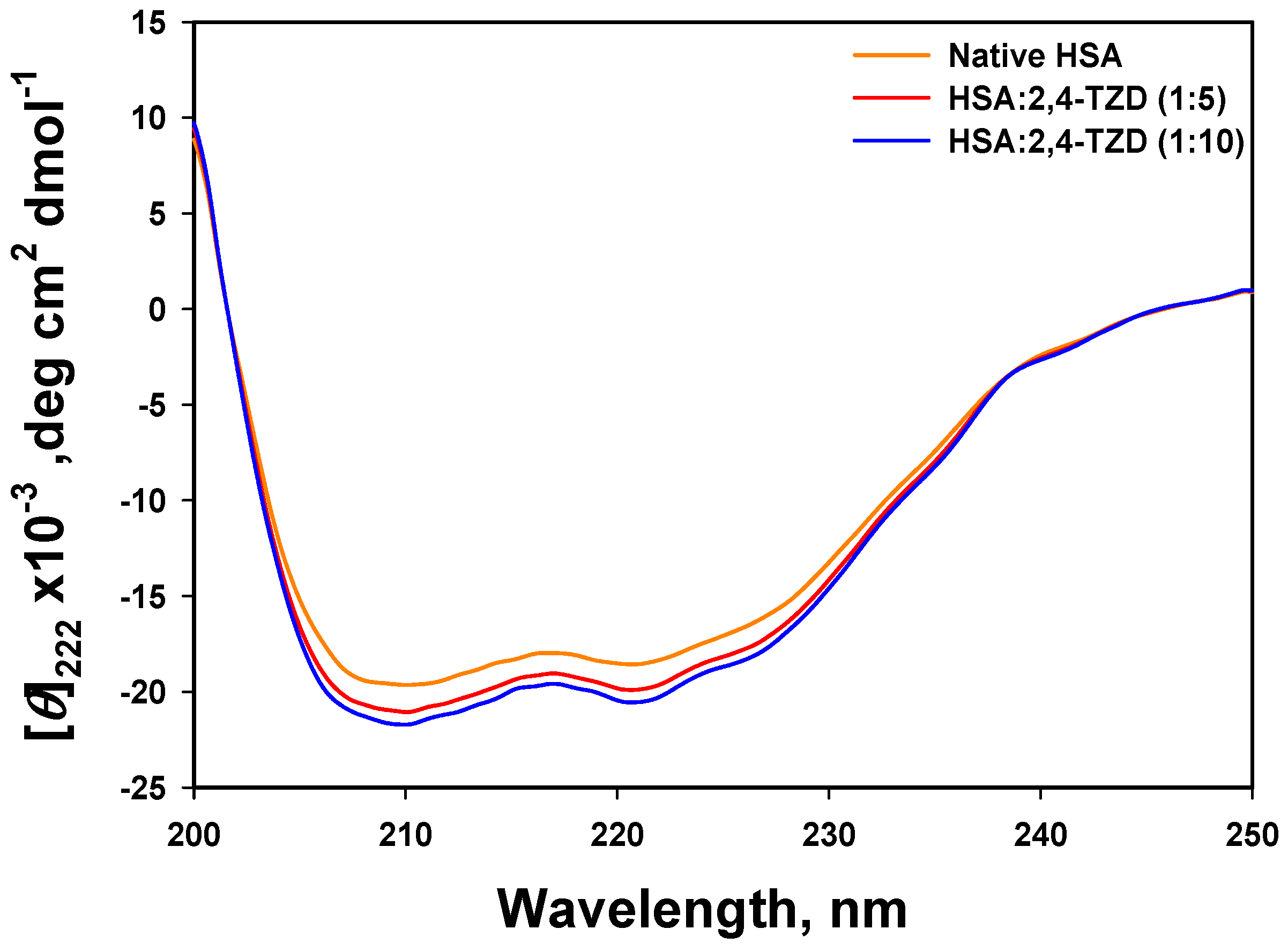
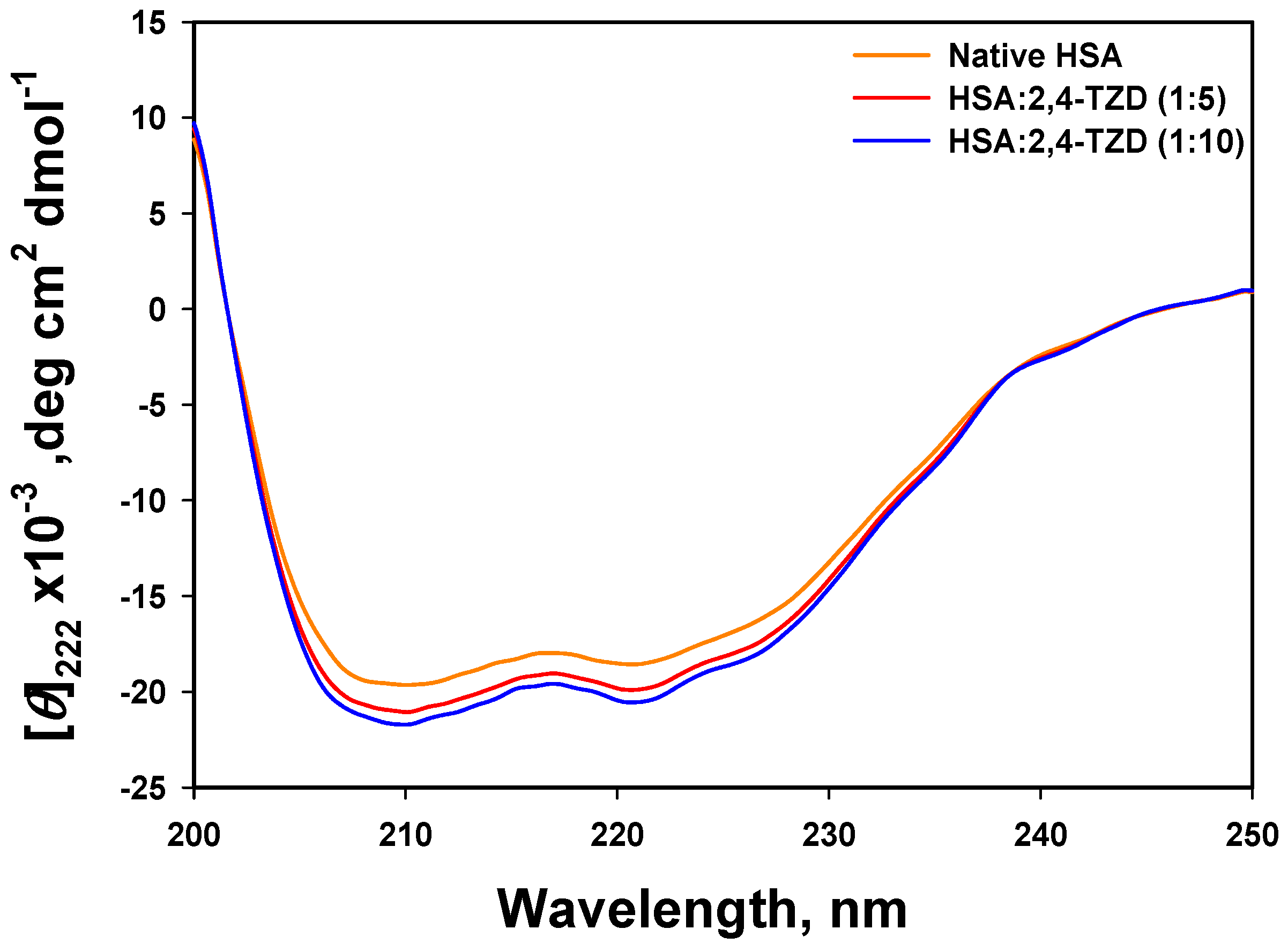
2.2.1. Far-UV Circular Dichroism (CD) Analysis
2.2.2. Synchronous Fluorescence Analysis
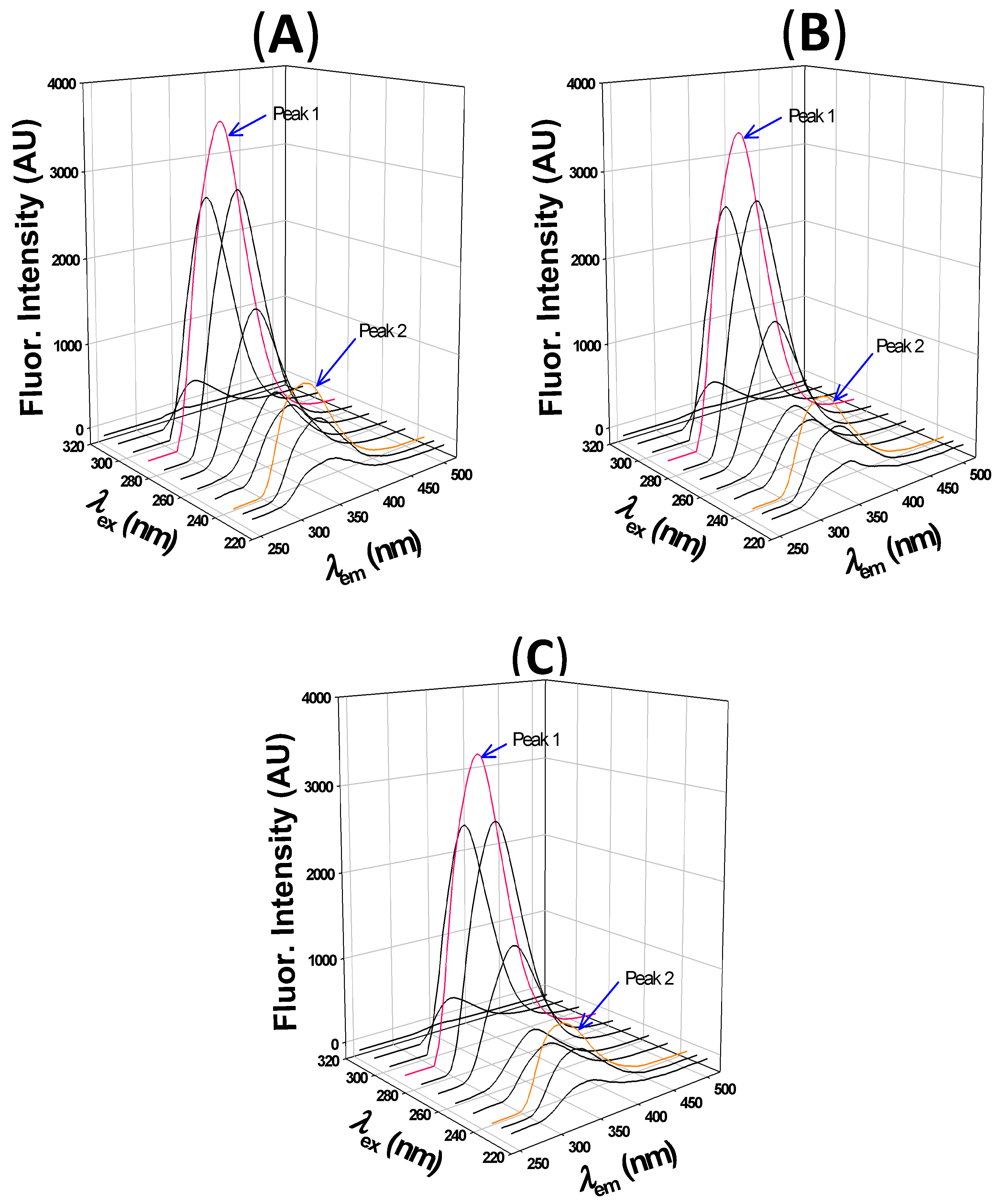
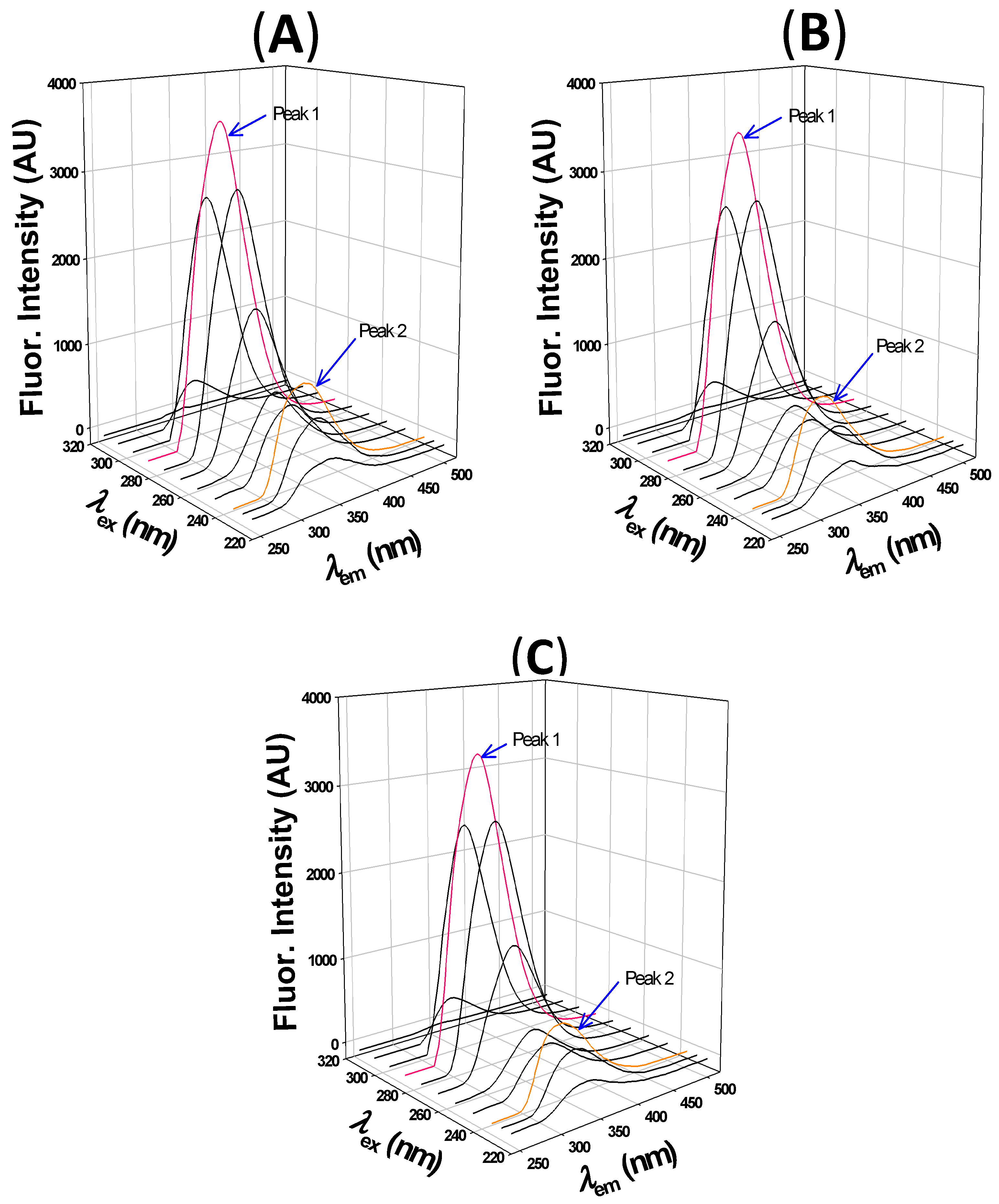
2.2.3. 3D fluorescence Analysis
2.3. Effect of 2,4-TZD on the thermodynamic Stability and Unfolding of HSA
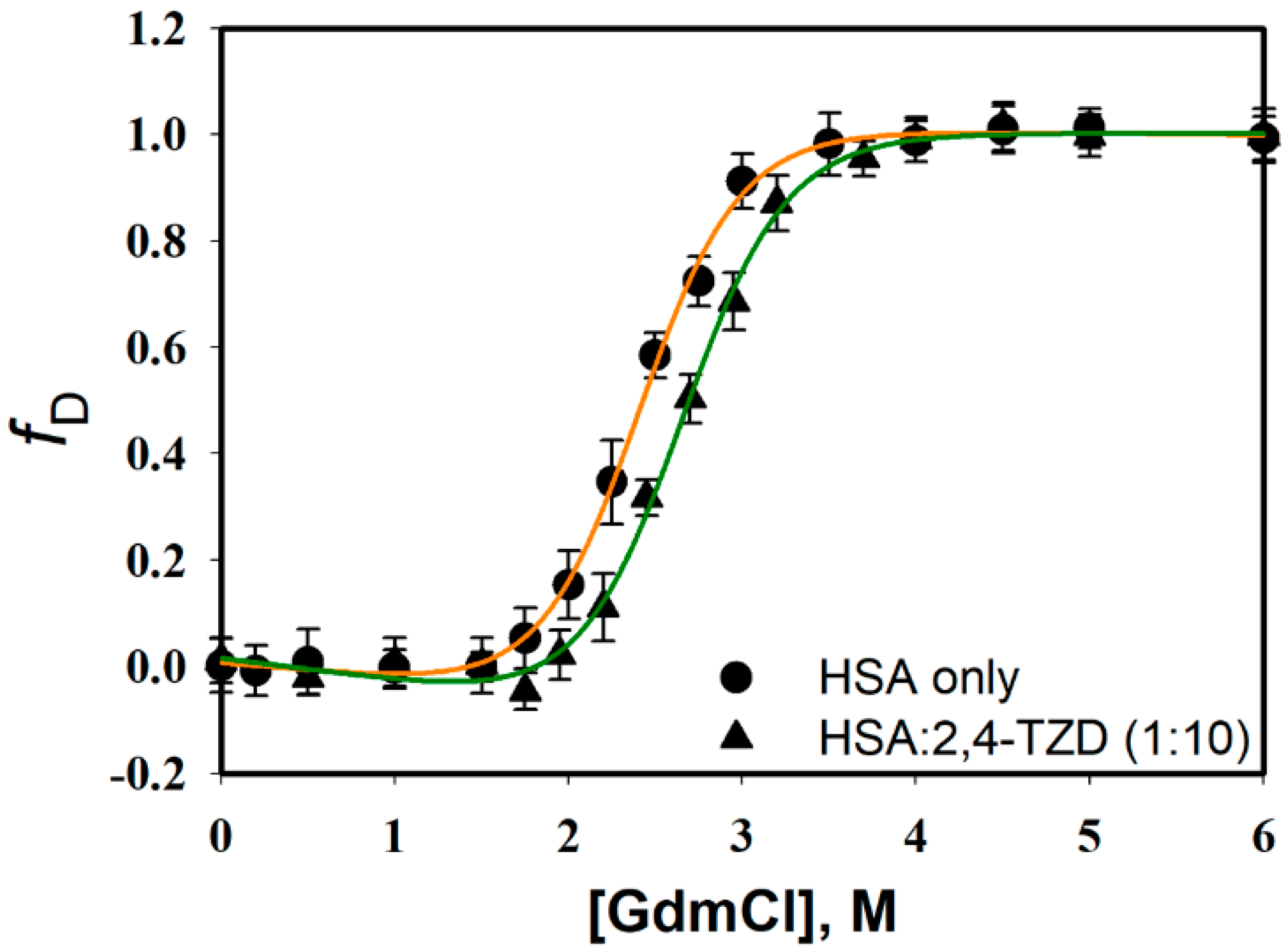
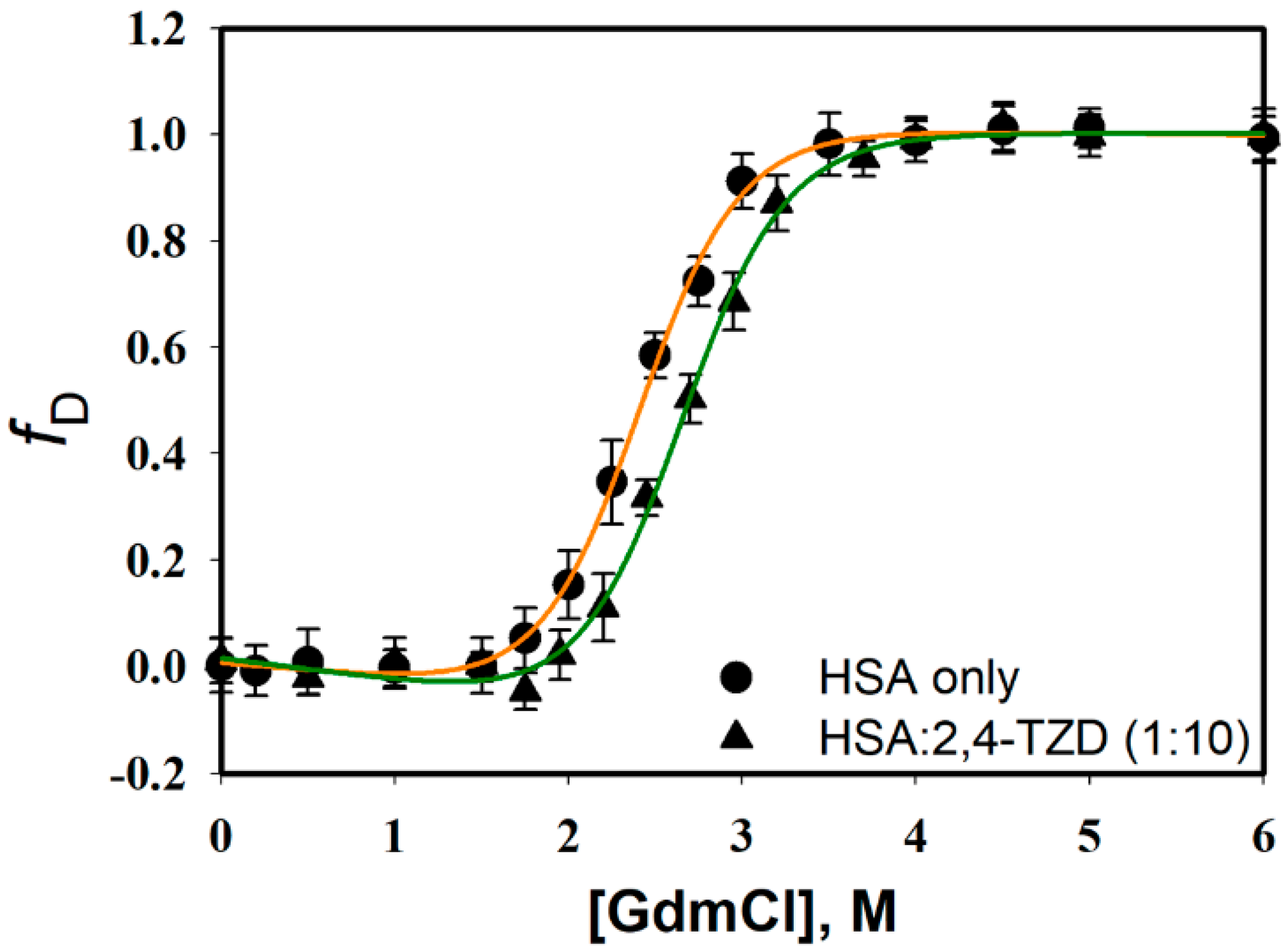
2.3.1. Guanidinium Chloride (GdmCl)-induced Denaturation
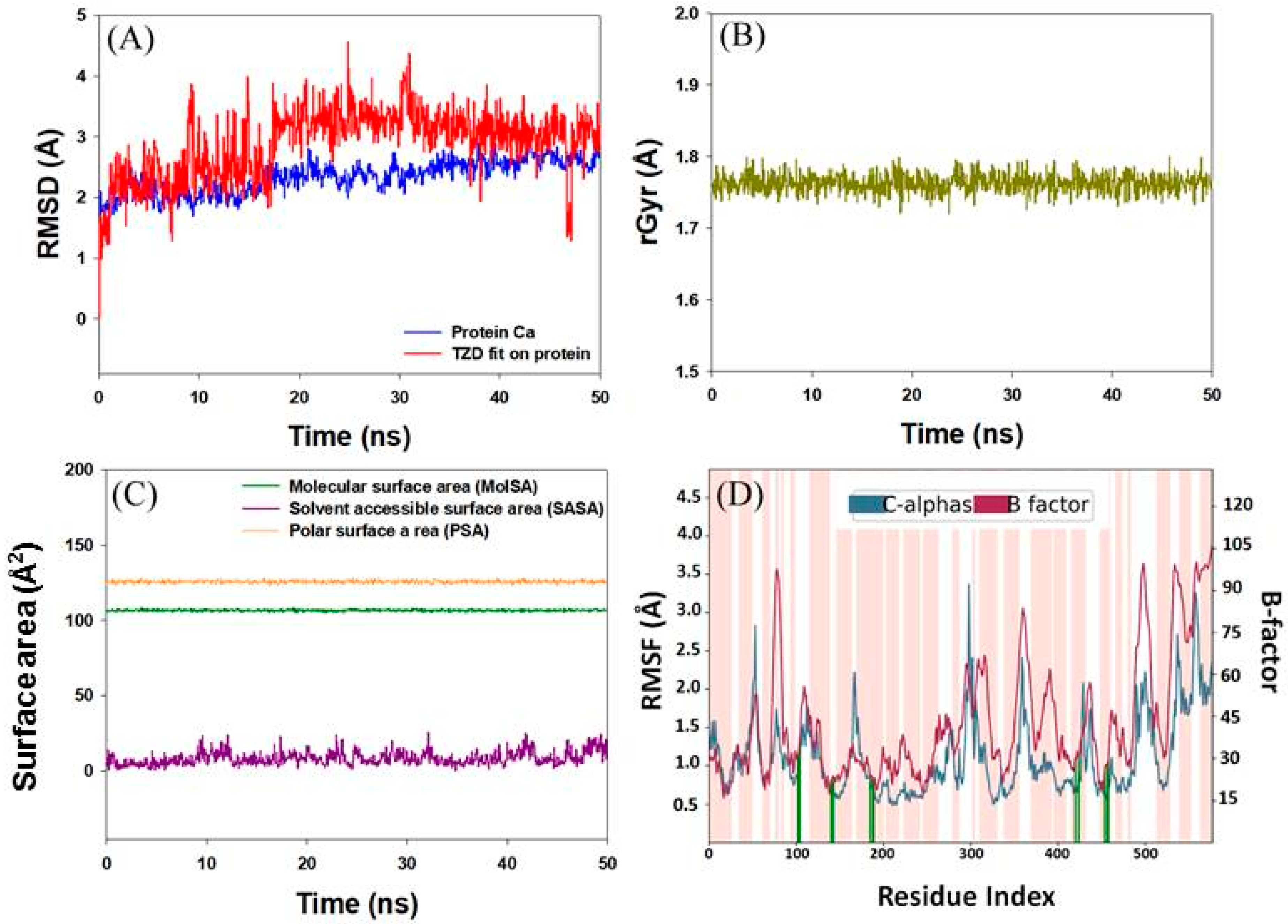
2.3.2. Analysis of Molecular Dynamics Simulation
3. Materials and Methods
3.1. Materials
3.2. Sample Preparation
3.3. Fluorescence Quenching Measurements
3.4. Synchronous and 3-D Fluorescence Measurements
3.5. Far-UV Circular Dichroism Measurements
3.6. Isothermal Titration Calorimetry (ITC) Measurements
3.7. Molecular Docking
3.8. Molecular Dynamics Simulation
3.9. GdmCl-induced Denaturation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HSA | Human Serum Albumin |
| 2,4-TZD | 2,4-thiazolidinedione |
| ITC | Isothermal Calorimetry |
| CD | Circular Dichroism |
| FRET | Förster Resonance Energy Transfer |
| LGA | Lamarck Genetic Algorithm |
References
- Cantello, B.C.C.; Cawthorne, M.A.; Cottam, G.P.; Duff, P.T.; Haigh, D.; Hindley, R.M.; Lister, C.A.; Smith, S.A.; Thurlby, L. [[.omega.-(Heterocyclylamino)alkoxy]benzyl]-2,4-thiazolidinediones as potent antihyperglycemic agents. J. Med. Chem. 1994, 37, 3977–3985. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J. Thiazolidinediones. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2007; pp. 1–2. [Google Scholar]
- Davidson, M.A.; Mattison, D.R.; Azoulay, L.; Krewski, D. Thiazolidinedione drugs in the treatment of type 2 diabetes mellitus: Past, present and future. Crit. Rev. Toxicol 2018, 48, 52–108. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, P.K.; Doble, M. Interaction of phytochemicals with hypoglycemic drugs on glucose uptake in L6 myotubes. Phytomedicine 2011, 18, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Meerarani, P.; Reiterer, G.; Toborek, M.; Hennig, B. Zinc modulates PPARgamma signaling and activation of porcine endothelial cells. J. Nutr. 2003, 133, 3058–3064. [Google Scholar] [CrossRef] [PubMed]
- Seree, E.; Villard, P.H.; Pascussi, J.M.; Pineau, T.; Maurel, P.; Nguyen, Q.B.; Fallone, F.; Martin, P.M.; Champion, S.; Lacarelle, B.; et al. Evidence for a new human CYP1A1 regulation pathway involving PPAR-alpha and 2 PPRE sites. Gastroenterology 2004, 127, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Panigrahy, D.; Singer, S.; Shen, L.Q.; Butterfield, C.E.; Freedman, D.A.; Chen, E.J.; Moses, M.A.; Kilroy, S.; Duensing, S.; Fletcher, C.; et al. PPARgamma ligands inhibit primary tumor growth and metastasis by inhibiting angiogenesis. J. Clin. Invest. 2002, 110, 923–932. [Google Scholar] [CrossRef]
- Elkamhawy, A.; Kim, N.Y.; Hassan, A.H.E.; Park, J.E.; Yang, J.E.; Oh, K.S.; Lee, B.H.; Lee, M.Y.; Shin, K.J.; Lee, K.T.; et al. Design, synthesis and biological evaluation of novel thiazolidinedione derivatives as irreversible allosteric IKK-beta modulators. Eur. J. Med. Chem. 2018, 157, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Ahn, S.N. Structure, enzymatic activities, glycation and therapeutic potential of human serum albumin: A natural cargo. Int. J. Biol. Macromol. 2019, 123, 979–990. [Google Scholar] [CrossRef]
- Sugio, S.; Kashima, A.; Mochizuki, S.; Noda, M.; Kobayashi, K. Crystal structure of human serum albumin at 2.5 Å resolution. Protein Eng. Des. Sel. 1999, 12, 439–446. [Google Scholar] [CrossRef]
- Curry, S.; Mandelkow, H.; Brick, P.; Franks, N. Crystal structure of human serum albumin complexed with fatty acid reveals an asymmetric distribution of binding sites. Nat. Struct. Biol. 1998, 5, 827–835. [Google Scholar] [CrossRef]
- He, X.M.; Carter, D.C. Atomic structure and chemistry of human serum albumin. Nature 1992, 358, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zsila, F. Subdomain IB is the third major drug binding region of human serum albumin: Toward the three-sites model. Mol. Pharm. 2013, 10, 1668–1682. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, E.; Rabbani, G.; Zaidi, N.; Singh, S.; Rehan, M.; Khan, M.M.; Rahman, S.K.; Quadri, Z.; Shadab, M.; Ashraf, M.T.; et al. Stereo-selectivity of human serum albumin to enantiomeric and isoelectronic pollutants dissected by spectroscopy, calorimetry and bioinformatics. PloS ONE 2011, 6, e26186. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Baig, M.H.; Lee, E.J.; Cho, W.K.; Ma, J.Y.; Choi, I. Biophysical Study on the Interaction between Eperisone Hydrochloride and Human Serum Albumin Using Spectroscopic, Calorimetric, and Molecular Docking Analyses. Mol. Pharm. 2017, 14, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Khan, M.J.; Ahmad, A.; Maskat, M.Y.; Khan, R.H. Effect of copper oxide nanoparticles on the conformation and activity of beta-galactosidase. Colloids Surf. B Biointerfaces 2014, 123, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.T.; Dey, P.; Hassan, M.I.; Ahmad, F.; Batra, J.K. Functional role of glutamine 28 and arginine 39 in double stranded RNA cleavage by human pancreatic ribonuclease. PloS ONE 2011, 6, e17159. [Google Scholar] [CrossRef]
- Rehman, M.T.; Faheem, M.; Khan, A.U. Insignificant beta-lactamase activity of human serum albumin: No panic to nonmicrobial-based drug resistance. Lett. Appl. Microbiol. 2013, 57, 325–329. [Google Scholar]
- Wardell, M.; Wang, Z.; Ho, J.X.; Robert, J.; Ruker, F.; Ruble, J.; Carter, D.C. The atomic structure of human methemalbumin at 1.9 A. Biochem. Biophys. Res. Commun. 2002, 291, 813–819. [Google Scholar] [CrossRef]
- Zunszain, P.A.; Ghuman, J.; McDonagh, A.F.; Curry, S. Crystallographic analysis of human serum albumin complexed with 4Z,15E-bilirubin-IXalpha. J. Mol. Biol. 2008, 381, 394–406. [Google Scholar] [CrossRef]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural basis of the drug-binding specificity of human serum albumin. J. Mol. Biol. 2005, 353, 38–52. [Google Scholar] [CrossRef]
- Petitpas, I.; Bhattacharya, A.A.; Twine, S.; East, M.; Curry, S. Crystal structure analysis of warfarin binding to human serum albumin: Anatomy of drug site I. J. Biol. Chem. 2001, 276, 22804–22809. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.H.; Rahman, S.; Rabbani, G.; Imran, M.; Ahmad, K.; Choi, I. Multi-Spectroscopic Characterization of Human Serum Albumin Binding with Cyclobenzaprine Hydrochloride: Insights from Biophysical and In Silico Approaches. Int J. Mol. Sci. 2019, 20, 662. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Baig, M.H.; Jan, A.T.; Ju Lee, E.; Khan, M.V.; Zaman, M.; Farouk, A.E.; Khan, R.H.; Choi, I. Binding of erucic acid with human serum albumin using a spectroscopic and molecular docking study. Int. J. Biol. Macromol. 2017, 105 Pt 3, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Lee, E.J.; Ahmad, K.; Baig, M.H.; Choi, I. Binding of Tolperisone Hydrochloride with Human Serum Albumin: Effects on the Conformation, Thermodynamics, and Activity of HSA. Mol. Pharm. 2018, 15, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Gonzalez, D.; Derendorf, H. Significance of protein binding in pharmacokinetics and pharmacodynamics. J. Pharm. Sci. 2010, 99, 1107–1122. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, G.A.; Bertucci, C.; Salvadori, P. Ligand binding to a human serum albumin stationary phase: Use of same-drug competition to discriminate pharmacologically relevant interactions. Biomed. Chromatogr 1998, 12, 248–254. [Google Scholar] [CrossRef]
- Bohnert, T.; Gan, L.S. Plasma protein binding: From discovery to development. J. Pharm. Sci. 2013, 102, 2953–2994. [Google Scholar] [CrossRef]
- Yeggoni, D.P.; Kuehne, C.; Rachamallu, A.; Subramanyam, R. Elucidating the binding interaction of andrographolide with the plasma proteins: Biophysical and computational approach. Rsc. Adv. 2017, 7, 5002–5012. [Google Scholar] [CrossRef]
- Yu, S.; Schuchardt, M.; Tölle, M.; van der Giet, M.; Zidek, W.; Dzubiella, J.; Ballauff, M. Interaction of human serum albumin with uremic toxins: A thermodynamic study. Rsc. Adv. 2017, 7, 27913–27922. [Google Scholar] [CrossRef]
- Ding, F.; Liu, W.; Zhang, X.; Wu, L.J.; Zhang, L.; Sun, Y. Identification of pyrazosulfuron-ethyl binding affinity and binding site subdomain IIA in human serum albumin by spectroscopic methods. Spectrochim Acta A Mol. Biomol. Spectrosc. 2010, 75, 1088–1094. [Google Scholar] [CrossRef]
- Bian, Q.; Liu, J.; Tian, J.; Hu, Z. Binding of genistein to human serum albumin demonstrated using tryptophan fluorescence quenching. Int. J. Biol. Macromol. 2004, 34, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Faridbod, F.; Ganjali, M.R.; Larijani, B.; Riahi, S.; Saboury, A.A.; Hosseini, M.; Norouzi, P.; Pillip, C. Interaction study of pioglitazone with albumin by fluorescence spectroscopy and molecular docking. Spectrochim Acta A Mol. Biomol. Spectrosc. 2011, 78, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; He, J.; Huang, Y.; Wang, Q.; Yang, H.; Xu, K.; Li, H. Interactions of cucurbit [6,7]urils with human serum albumin and their effects on zaltoprofen transportation. Rsc. Adv. 2016, 6, 85811–85819. [Google Scholar] [CrossRef]
- Rehman, M.T.; Shamsi, H.; Khan, A.U. Insight into the binding mechanism of imipenem to human serum albumin by spectroscopic and computational approaches. Mol. Pharm. 2014, 11, 1785–1797. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.; Joshi, R.; Ottoor, D. Spectroscopic and molecular docking study to understand the binding interaction of rosiglitazone with bovine serum albumin in presence of valsartan. J. Lumin. 2018, 197, 200–210. [Google Scholar] [CrossRef]
- Ware, W.R. Oxygen quenching of fluorescence in solution: An experimental study of the diffusion process. J. Phys. Chem. 1962, 66, 455–458. [Google Scholar] [CrossRef]
- Kamtekar, N.; Pandey, A.; Agrawal, N.; Pissurlenkar, R.R.; Borana, M.; Ahmad, B. Interaction of multimicrobial synthetic inhibitor 1,2-bis(2-benzimidazolyl)-1,2-ethanediol with serum albumin: Spectroscopic and computational studies. PloS ONE 2013, 8, e53499. [Google Scholar] [CrossRef]
- Zaidi, N.; Ajmal, M.R.; Rabbani, G.; Ahmad, E.; Khan, R.H. A comprehensive insight into binding of hippuric acid to human serum albumin: A study to uncover its impaired elimination through hemodialysis. PloS ONE 2013, 8, e71422. [Google Scholar] [CrossRef]
- Yeggoni, D.P.; Gokara, M.; Manidhar, D.M.; Rachamallu, A.; Nakka, S.; Reddy, C.S.; Subramanyam, R. Binding and molecular dynamics studies of 7-hydroxycoumarin derivatives with human serum albumin and its pharmacological importance. Mol. Pharm. 2014, 11, 1117–1131. [Google Scholar] [CrossRef]
- Ishtikhar, M.; Rabbani, G.; Khan, R.H. Interaction of 5-fluoro-5’-deoxyuridine with human serum albumin under physiological and non-physiological condition: A biophysical investigation. Colloids Surf. B Biointerfaces 2014, 123, 469–477. [Google Scholar] [CrossRef]
- Stryer, L.; Haugland, R.P. Energy transfer: A spectroscopic ruler. Proc. Natl. Acad. Sci. USA 1967, 58, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Förster, T. Intramolecular energy transfer and fluorescence. Ann. Phys. Berl. 1948, 437, 55–75. [Google Scholar] [CrossRef]
- Cyril, L.; Earl, J.K.; Sperry, W.M. Biochemists Handbook; E & FN Epon Led. Press: London, UK, 1961. [Google Scholar]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Yang, J.T.; Martinez, H.M. Determination of the secondary structures of proteins by circular dichroism and optical rotatory dispersion. Biochemistry 1972, 11, 4120–4131. [Google Scholar] [CrossRef] [PubMed]
- Suo, Z.; Sun, Q.; Yang, H.; Tang, P.; Gan, R.; Xiong, X.; Li, H. Combined spectroscopy methods and molecular simulations for the binding properties of trametinib to human serum albumin. Rsc. Adv. 2018, 8, 4742–4749. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.T.; Ahmed, S.; Khan, A.U. Interaction of meropenem with ‘N’ and ‘B’ isoforms of human serum albumin: A spectroscopic and molecular docking study. J. Biomol. Struct. Dyn. 2016, 34, 1849–1864. [Google Scholar] [CrossRef] [PubMed]
- Farruggia, B.; Garcia, G.; D’Angelo, C.; Pico, G. Destabilization of human serum albumin by polyethylene glycols studied by thermodynamical equilibrium and kinetic approaches. Int. J. Biol. Macromol. 1997, 20, 43–51. [Google Scholar] [CrossRef]
- Farruggia, B.; Pico, G.A. Thermodynamic features of the chemical and thermal denaturations of human serum albumin. Int. J. Biol. Macromol. 1999, 26, 317–323. [Google Scholar] [CrossRef]
- Ahmad, B.; Muteeb, G.; Alam, P.; Varshney, A.; Zaidi, N.; Ishtikhar, M.; Badr, G.; Mahmoud, M.H.; Khan, R.H. Thermal induced unfolding of human serum albumin isomers: Assigning residual alpha helices to domain II. Int. J. Biol. Macromol. 2015, 75, 447–452. [Google Scholar] [CrossRef]
- AlAjmi, M.F.; Hussain, A.; Rehman, M.T.; Khan, A.A.; Shaikh, P.A.; Khan, R.A. Design, Synthesis, and Biological Evaluation of Benzimidazole-Derived Biocompatible Copper(II) and Zinc(II) Complexes as Anticancer Chemotherapeutics. Int. J. Mol. Sci. 2018, 19, 1492. [Google Scholar] [CrossRef]
- Rehman, M.T.; AlAjmi, M.F.; Hussain, A.; Rather, G.M.; Khan, M.A. High-Throughput Virtual Screening, Molecular Dynamics Simulation, and Enzyme Kinetics Identified ZINC84525623 as a Potential Inhibitor of NDM-1. Int. J. Mol. Sci. 2019, 20, 819. [Google Scholar] [CrossRef] [PubMed]
- AlAjmi, M.F.; Rehman, M.T.; Hussain, A.; Rather, G.M. Pharmacoinformatics approach for the identification of Polo-like kinase-1 inhibitors from natural sources as anti-cancer agents. Int. J. Biol. Macromol. 2018, 116, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Branka, A.C. Nose-Hoover chain method for nonequilibrium molecular dynamics simulation. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat Interdiscip Top. 2000, 61, 4769–4773. [Google Scholar]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Santoro, M.M.; Bolen, D.W. Unfolding free energy changes determined by the linear extrapolation method. 1. Unfolding of phenylmethanesulfonyl alpha-chymotrypsin using different denaturants. Biochemistry 1988, 27, 8063–8068. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Ahmad, F. A New Method for the Determination of Stability Parameters of Proteins from Their Heat-Induced Denaturation Curves. Anal. Biochem. 2000, 283, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Yadav, S.; Ahmad, R.; Ahmad, F. A possible origin of differences between calorimetric and equilibrium estimates of stability parameters of proteins. Biochem. J. 2000, 345 Pt 3, 711–717. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 298 K | 303 K | 310 K | 315 K |
|---|---|---|---|---|
| KSV × 103 (M−1) | 2.67 ± 0.21 | 2.10 ± 0.18 | 1.63 ± 0.14 | 1.27 ± 0.10 |
| kq × 1011 (M−1 s−1) | 4.67 ± 0.39 | 3.68 ± 0.33 | 2.85 ± 0.27 | 2.22 ± 0.21 |
| Kb × 103 (M−1) | 1.69 ± 0.15 | 2.75 ± 0.21 | 3.90 ± 0.26 | 8.42 ± 0.29 |
| n (binding stoichiometry) | 0.950 ± 0.02 | 1.02 ± 0.03 | 1.08 ± 0.02 | 1.18 ± 0.04 |
| ΔHo (kcal mol−1) | 16.34 ± 0.96 | |||
| TΔSo (kcal mol−1) | 20.73 ± 1.32 | 21.07 ± 1.08 | 21.56 ± 1.27 | 21.91 ± 1.18 |
| ΔGo (kcal mol−1) | −4.39 ± 0.63 | −4.73 ± 0.44 | −5.22 ± 0.59 | −5.57 ± 0.52 |
| Ka (M−1) | ΔHo, (kcal mol−1) | TΔSo (kcal mol−1) | ΔGo (kcal mol−1) |
|---|---|---|---|
| 2.1 ± 0.11 × 103 | −14.4 ± 1.21 | −8.65 ± 0.85 | −5.75 ± 0.48 |
| J (M−1 cm3) | Ro (nm) | r (nm) |
|---|---|---|
| 3.08 × 10−14 | 2.96 | 4.08 |
| Nature of Interaction | Category of Interaction | Distance (Å) | ΔG (kcal mol−1) | Kb × 103 (M−1) |
|---|---|---|---|---|
| Tyr148:HN-Lig:O | Hydrogen Bond | 2.07 ± 0.06 | −4.61 ± 0.54 | 1.69 ± 0.15 |
| Lig:H-Ser193:O | Hydrogen Bond | 2.57 ± 0.08 | ||
| Ala194:CA-Lig:O | Carbon Hydrogen Bond | 3.38 ± 0.08 |
| Peak 1 | Peak 2 | |||||
|---|---|---|---|---|---|---|
| Peak Position (λex/λem) nm/nm | Fluorescence Intensity (AU) | Stokes Shift (nm) | Peak Position (λex/λem) nm/nm | Fluorescence Intensity (AU) | Stokes Shift (nm) | |
| HSA | 280/335 | 3607 | 55 | 230/327 | 1077 | 97 |
| HSA:2,4-TZD (1:5) | 280/332 | 3488 | 52 | 230/324 | 935 | 94 |
| HSA:2,4-TZD (1:10) | 280/332 | 3406 | 52 | 230/324 | 809 | 94 |
| ΔGDo, kcal mol−1 | mg, kcal mol−1 M−1 | Cm, M | |
|---|---|---|---|
| HSA only | 4.94 ± 0.32 | 2.05 ± 0.14 | 2.40 ± 0.12 |
| HSA:2,4-TZD (1:1) | 5.30 ± 0.48 | 2.01 ± 0.17 | 2.61 ± 0.19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, S.; Rehman, M.T.; Rabbani, G.; Khan, P.; AlAjmi, M.F.; Hassan, M.I.; Muteeb, G.; Kim, J. Insight of the Interaction between 2,4-thiazolidinedione and Human Serum Albumin: A Spectroscopic, Thermodynamic and Molecular Docking Study. Int. J. Mol. Sci. 2019, 20, 2727. https://doi.org/10.3390/ijms20112727
Rahman S, Rehman MT, Rabbani G, Khan P, AlAjmi MF, Hassan MI, Muteeb G, Kim J. Insight of the Interaction between 2,4-thiazolidinedione and Human Serum Albumin: A Spectroscopic, Thermodynamic and Molecular Docking Study. International Journal of Molecular Sciences. 2019; 20(11):2727. https://doi.org/10.3390/ijms20112727
Chicago/Turabian StyleRahman, Safikur, Md Tabish Rehman, Gulam Rabbani, Parvez Khan, Mohamed F AlAjmi, Md. Imtaiyaz Hassan, Ghazala Muteeb, and Jihoe Kim. 2019. "Insight of the Interaction between 2,4-thiazolidinedione and Human Serum Albumin: A Spectroscopic, Thermodynamic and Molecular Docking Study" International Journal of Molecular Sciences 20, no. 11: 2727. https://doi.org/10.3390/ijms20112727