Is It Possible to Predict the Odor of a Molecule on the Basis of its Structure?

 , ,
, ,  and
and
Abstract
:
1. Introduction
1.1. The Crucial Roles of the Olfactory Sense
1.2. Human Response to Odor
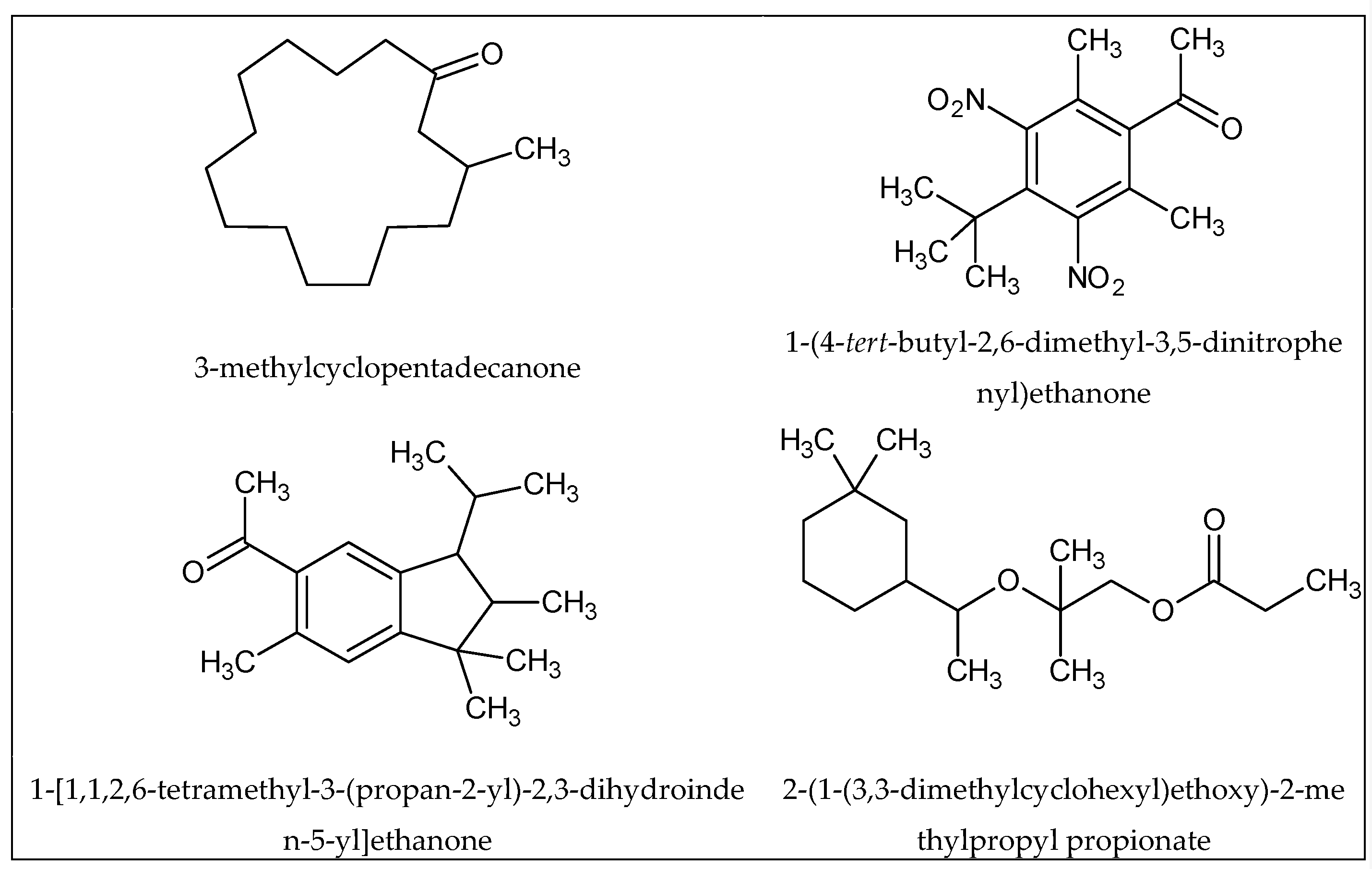
1.3. Physical and Chemical Features of Odor
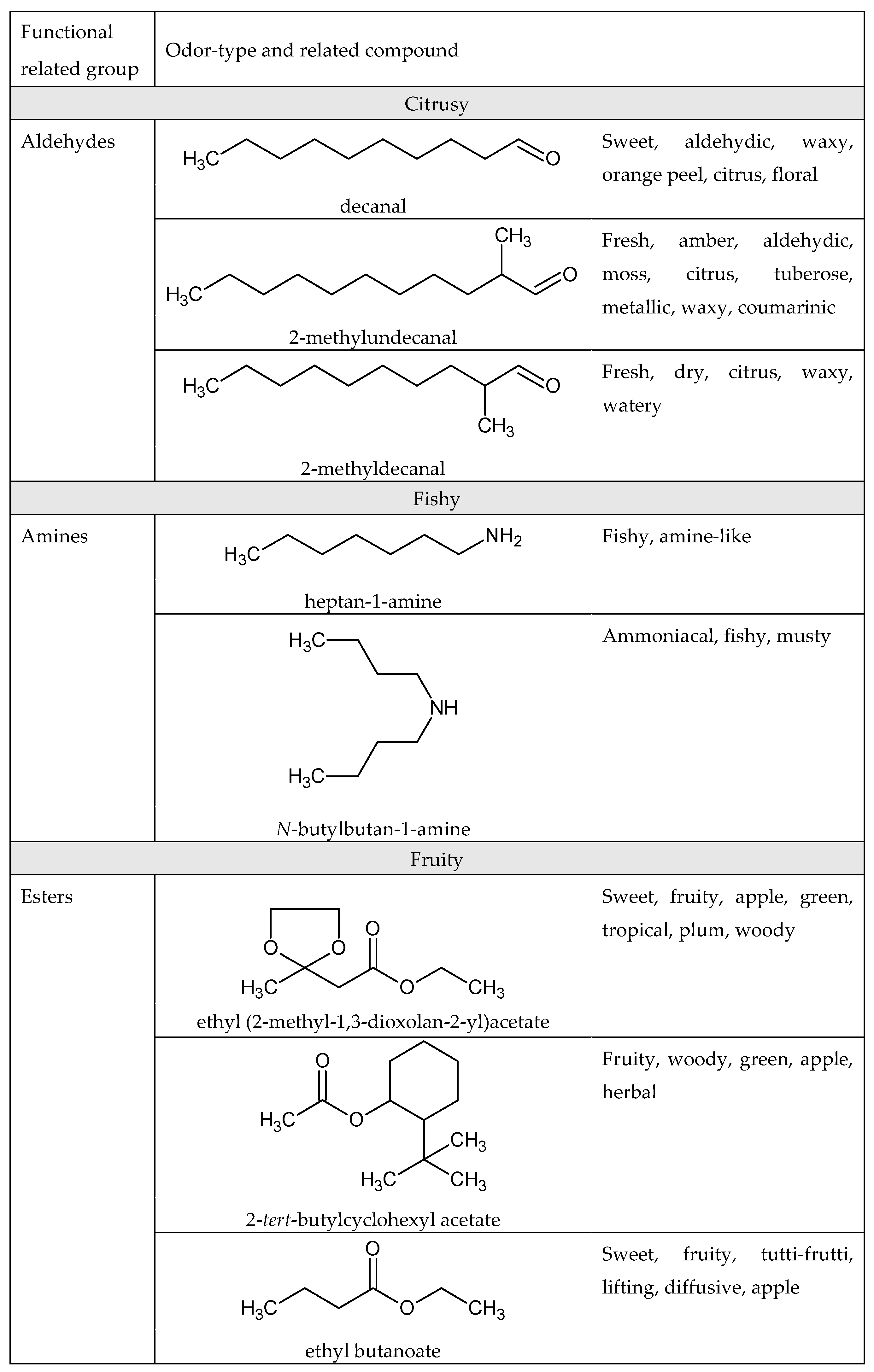
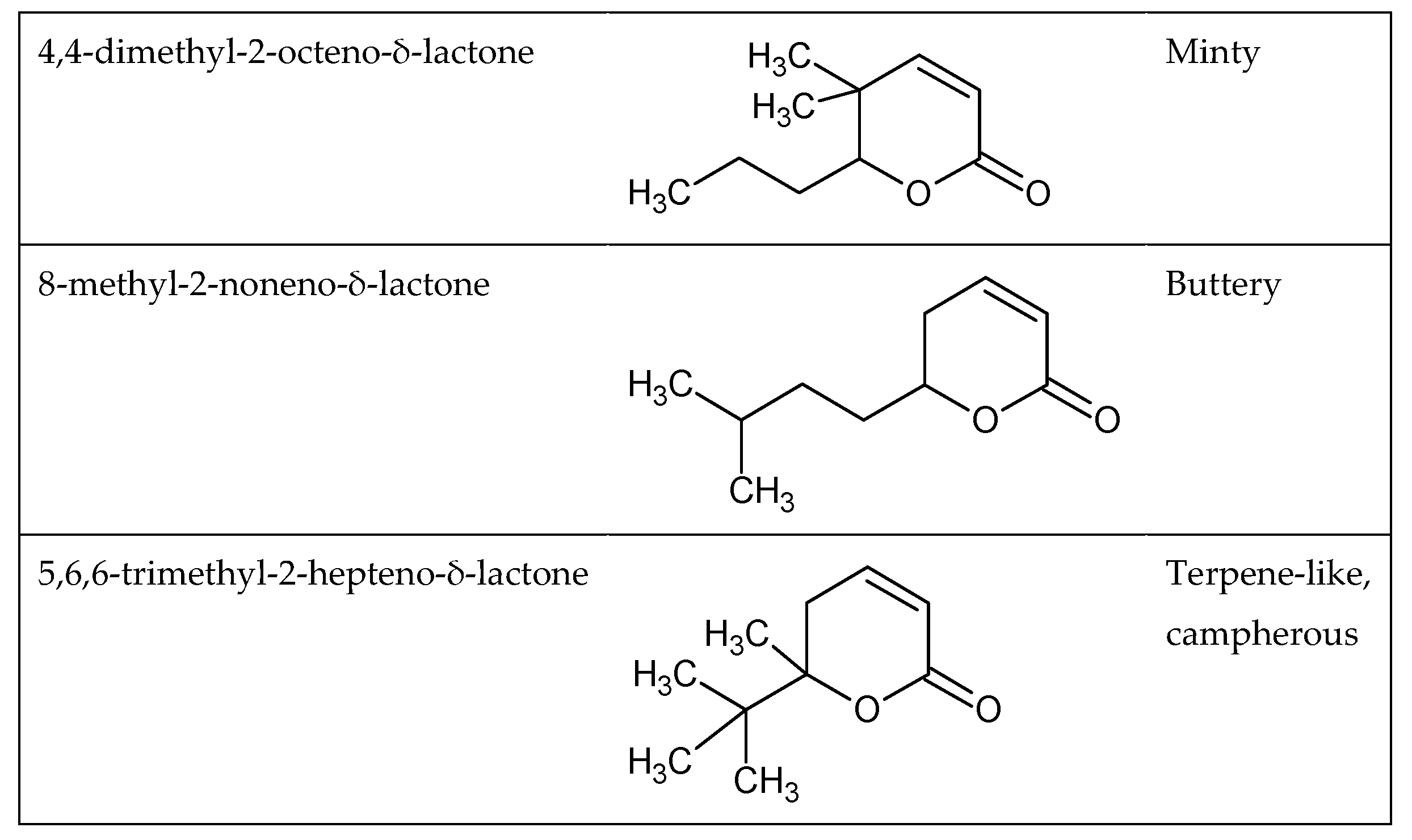
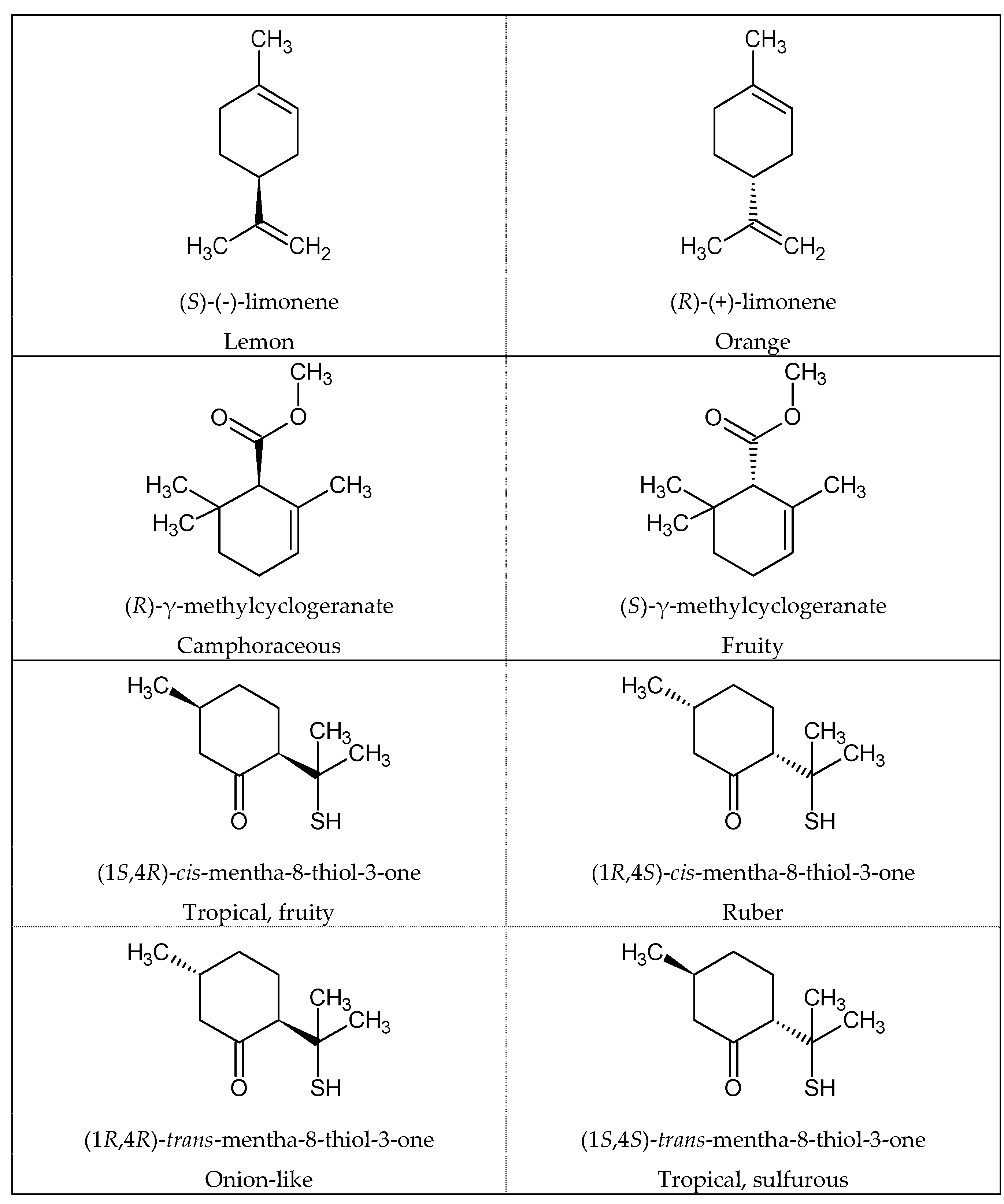
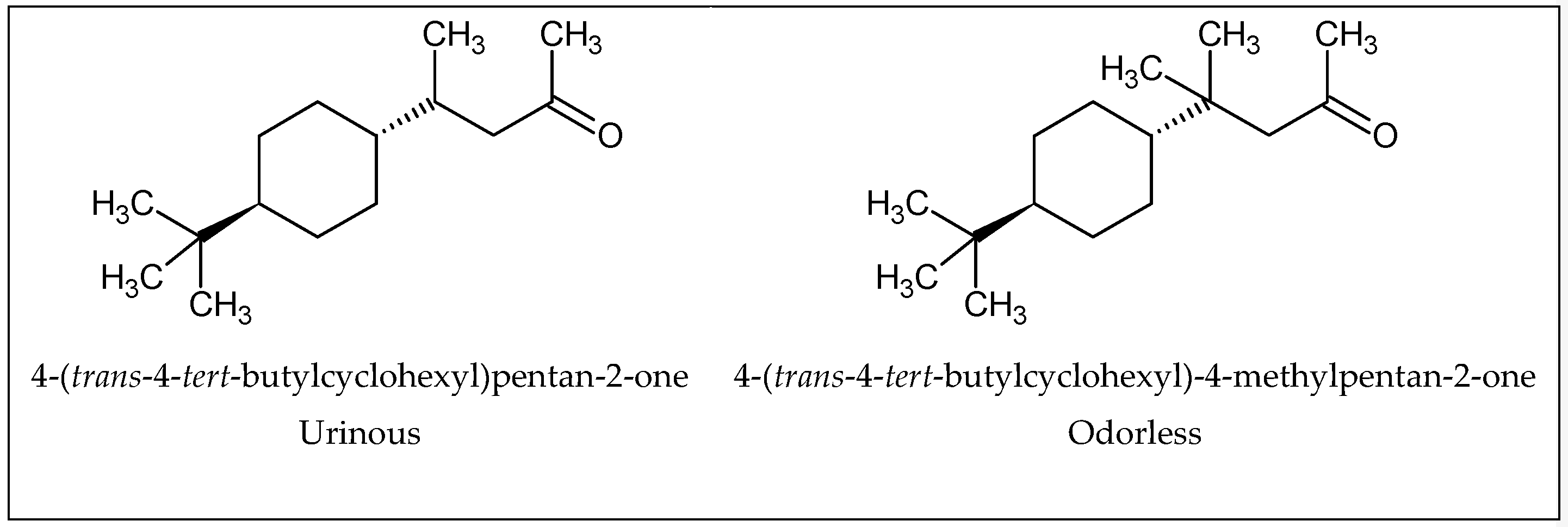
1.4. Odor and Structure Relationship
2. Olfaction Mechanisms
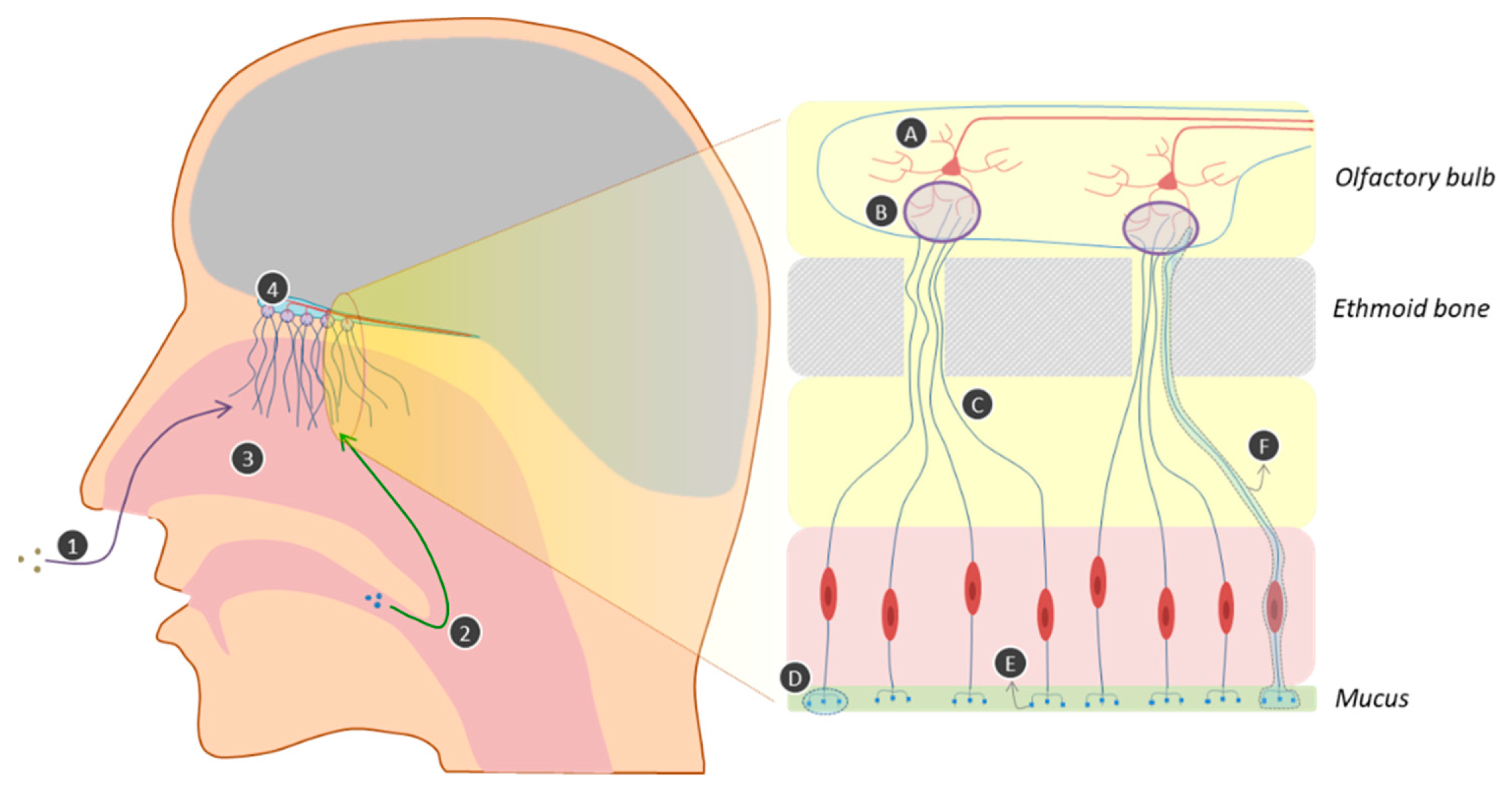
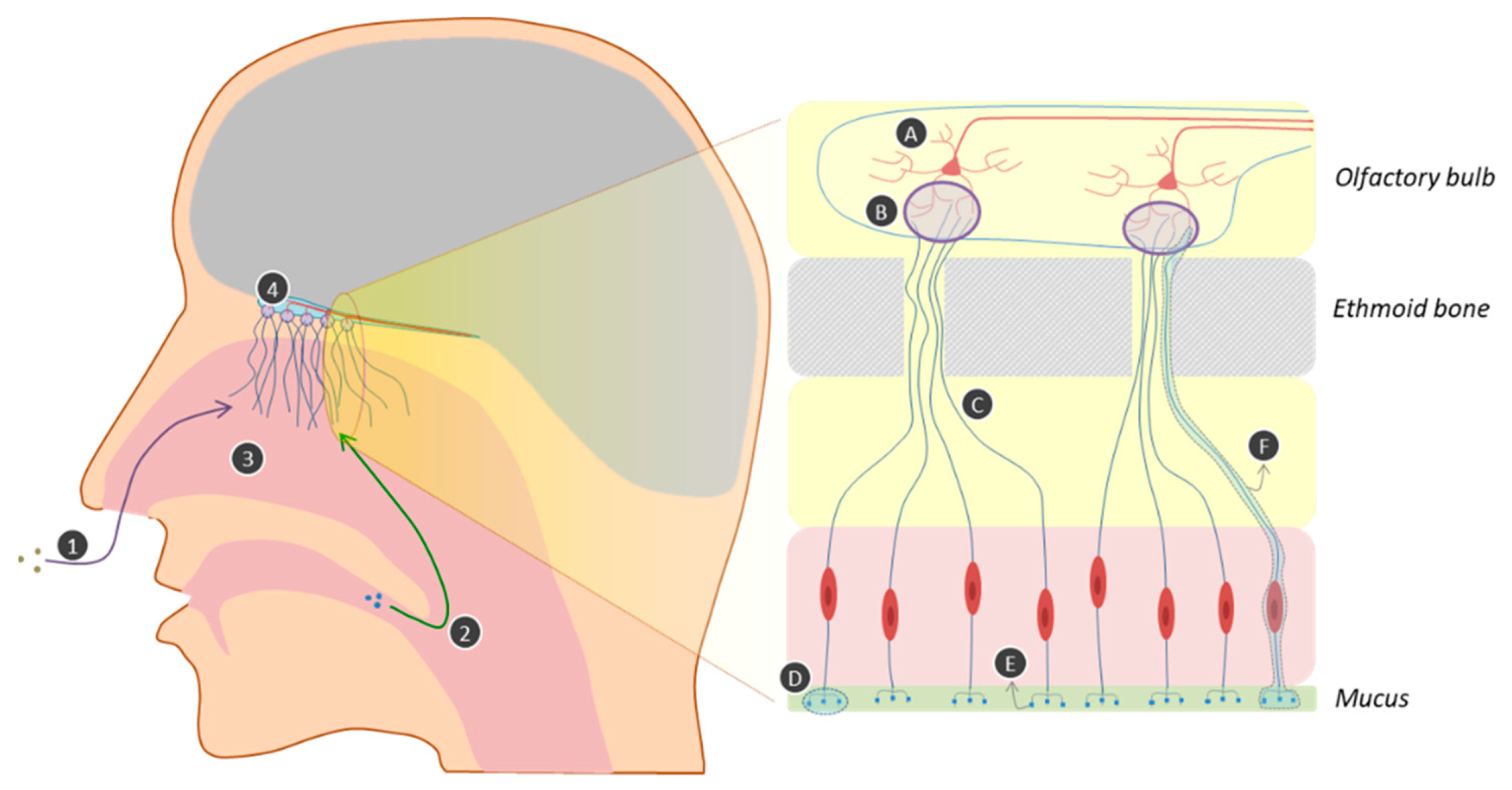
2.1. The Physiology of Olfaction
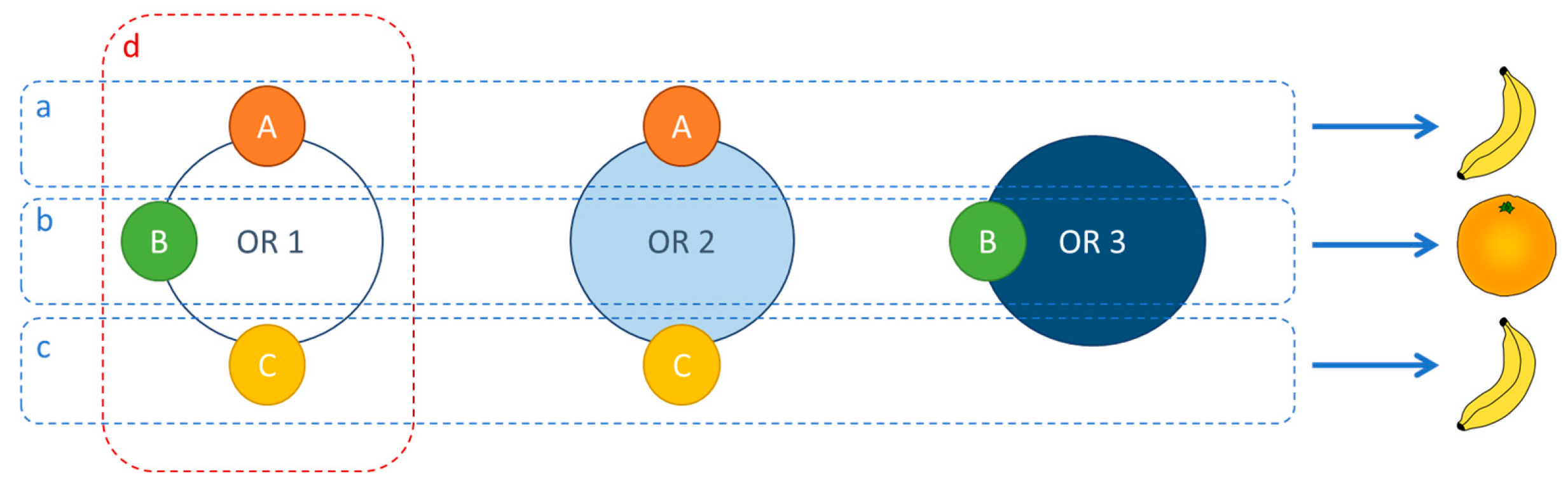
2.2. Olfaction Theory: Studying Structure–Odor Relationships
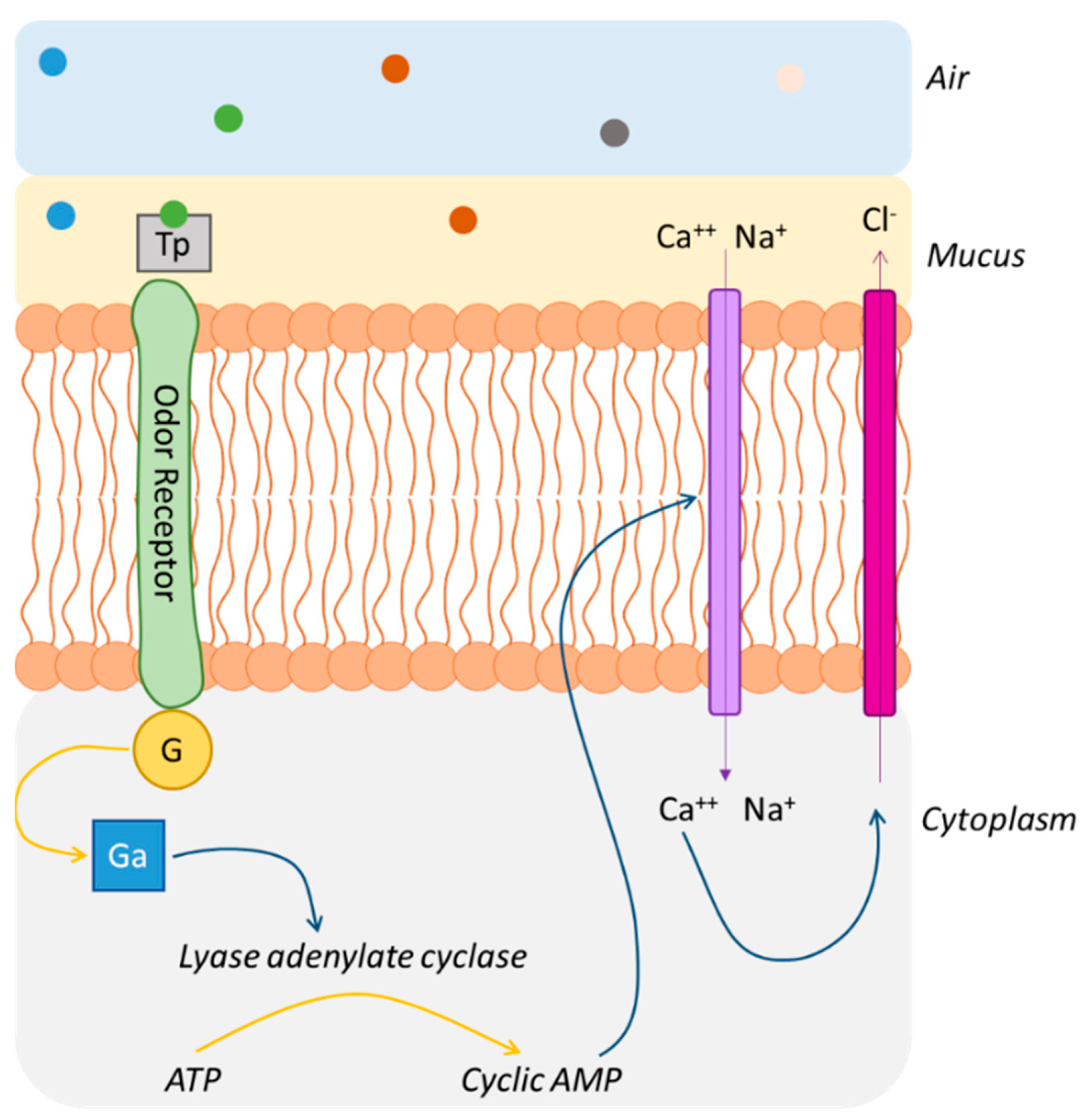
2.3. Odorant–Receptor Interactions
3. New Trends in Olfaction Studies
3.1. Importance and Diversity of Human Olfaction
3.2. Electronic Nose and Machine Learning
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Depelteau, J.S.; Brenzinger, S.; Briegel, A. Bacterial and Archaeal Cell Structure. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Grabe, V.; Sachse, S. Fundamental principles of the olfactory code. Biosystems 2018, 164, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.-E.; Han, J.H. How Flavor Works—The Science of Taste and Aroma; John Wiley & Sons, Ltd.: Chichester, UK, 2015. [Google Scholar]
- Padodara, R.J.; Jacob, N. Olfactory Sense in Different Animals. Indian J. Vet. Sci. 2014, 2, 1–14. [Google Scholar]
- Calvo-Ochoa, E.; Byrd-Jacobs, C. The Olfactory System of Zebrafish as a Model for the Study of Neurotoxicity and Injury: Implications for Neuroplasticity and Disease. Int. J. Mol. Sci. 2019, 20, 1639. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, Y.; Luo, Y.; Zhang, J.; Zhai, Y.; Yang, D.; Zhang, Z.; Li, Y.; Storm, D.; Ma, R. Gene Expression Profiles of Main Olfactory Epithelium in Adenylyl Cyclase 3 Knockout Mice. Int. J. Mol. Sci. 2015, 16, 28320–28333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raji, J.I.; DeGennaro, M. Genetic analysis of mosquito detection of humans. Curr. Opin. Insect Sci. 2017, 20, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.R.; Knaapila, A. Genetics of Taste and Smell: Poisons and Pleasures. NIH Public Access 2010, 94, 213–240. [Google Scholar]
- Vaglio, S. Chemical communication and mother-infant recognition. Commun. Integr. Biol. 2009, 2, 279–281. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, A.C. Ethnography and the senses: Challenges and possibilities. Qual. Res. Sport Exerc. 2009, 1, 21–35. [Google Scholar] [CrossRef]
- Sarafoleanu, C.; Mella, C.; Georgescu, M.; Perederco, C. The importance of the olfactory sense in the human behavior and evolution. J. Med. Life 2009, 2, 196–198. [Google Scholar]
- Ferdenzi, C.; Joussain, P.; Digard, B.; Luneau, L.; Djordjevic, J.; Bensafi, M. Individual Differences in Verbal and Non-Verbal Affective Responses to Smells: Influence of Odor Label Across Cultures. Chem. Senses 2016, 42, 37–46. [Google Scholar] [CrossRef]
- Hummel, T.; Kobal, G.; Gudziol, H.; Mackay-Sim, A. Normative data for the “Sniffin’ Sticks” including tests of odor identification, odor discrimination, and olfactory thresholds: An upgrade based on a group of more than 3,000 subjects. Eur. Arch. Oto-Rhino-Laryngology 2007, 264, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Sorokowska, A.; Hummel, T. Polska wersja testu Sniffin’ Sticks—Adaptacja i normalizacja. Otolaryngol. Pol. 2014, 68, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Sorokowska, A.; Schriever, V.A.; Gudziol, V.; Hummel, C.; Hähner, A.; Iannilli, E.; Sinding, C.; Aziz, M.; Seo, H.S.; Negoias, S.; et al. Changes of olfactory abilities in relation to age: Odor identification in more than 1400 people aged 4 to 80 years. Eur. Arch. Oto-Rhino-Laryngology 2015, 272, 1937–1944. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.; Shaman, P.; Applebaum, S.; Giberson, R.; Siksorski, L.; Rosenberg, L. Smell identification ability: Changes with age. Science 1984, 226, 1441–1443. [Google Scholar] [CrossRef] [PubMed]
- Sorokowski, P.; Karwowski, M.; Misiak, M.; Marczak, M.K.; Dziekan, M.; Hummel, T.; Sorokowska, A. Sex differences in human olfaction: A meta-analysis. Front. Psychol. 2019, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- John, P.J. Basics of Biopsychology, 7th ed.; Allyn & Bacon, Inc.: London, UK, 2007. [Google Scholar]
- Herrmann, A. The Chemistry and Biology of Volatiles; Herrmann, A., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2010; p. 428. [Google Scholar]
- De Gennaro, G.; Farella, G.; Marzocca, A.; Mazzone, A.; Tutino, M. Indoor and outdoor monitoring of volatile organic compounds in school buildings: Indicators based on health risk assessment to single out critical issues. Int. J. Environ. Res. Public Health 2013, 10, 6273–6291. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, F. Structure-odor relations: A modern perspective. In Handbook of Olfaction and Gustation; Doty, R.L., Ed.; Taylor & Francis Group: Abingdon, UK, 2003; pp. 457–492. [Google Scholar]
- Nara, K.; Saraiva, L.R.; Ye, X.; Buck, L.B. A Large-Scale Analysis of Odor Coding in the Olfactory Epithelium. J. Neurosci. 2011, 31, 9179–9191. [Google Scholar] [CrossRef] [Green Version]
- Dufossé, L.; Latrasse, A.; Spinnler, H.-E. Importance des lactones dans les arômes alimentaires. Sci des Aliment. 1994, 14, 17–50. [Google Scholar]
- Goldstein, N. Getting to know the odor compounds. Biocycle 2002, 43, 42–44. [Google Scholar]
- Buettner, A. Springer Handbook of Odor; Buettner, A., Ed.; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Laing, D.G. Relationship between Molecular Structure, Concentration and Odor Qualities of Oxygenated Aliphatic Molecules. Chem. Senses 2003, 28, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Poivet, E.; Tahirova, N.; Peterlin, Z.; Xu, L.; Zou, D.-J.; Acree, T.; Firestein, S. Functional odor classification through a medicinal chemistry approach. Sci. Adv. 2018, 4, eaao6086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarzo, M. Effect of Functional Group and Carbon Chain Length on the Odor Detection Threshold of Aliphatic Compounds. Sensors 2012, 12, 4105–4112. [Google Scholar] [CrossRef] [PubMed]
- Nobuhara, A. Syntheses of Unsaturated Lactones. Agric. Biol. Chem. 1968, 32, 1016–1020. [Google Scholar]
- Brookes, J.C.; Horsfield, A.P.; Stoneham, A.M. Odour character differences for enantiomers correlate with molecular flexibility. J. R. Soc. Interface 2009, 6, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Boelens, M.H.; van Gemert, L.J. Volatile character-impact sulfur compounds and their sensory properties. Perfum. Flavorist 1993, 18, 29–39. [Google Scholar]
- Meierhenrich, U.J.; Golebiowski, J.; Fernandez, X.; Cabrol-Bass, D. De la molécule à l’odeur: Les bases moléculaires des premières étapes de l’olfaction. L. Actual. Chim. 2005, 29–40. [Google Scholar]
- Rossiter, K.J. Structure-odor relationships. Chem. Rev. 1996, 96, 3201–3240. [Google Scholar] [CrossRef]
- Kraft, P.; Bajgrowicz, J.A.; Denis, C.; Frater, G. Odds and Trends: Recent Developments in the Chemistry of Odorants. Angew. Chemie Int. Ed. 2000, 39, 2980–3010. [Google Scholar] [CrossRef]
- Brenna, E.; Fuganti, C.; Serra, S. Enantioselective perception of chiral odorants. Tetrahedron: Asymmetry 2003, 14, 1–42. [Google Scholar] [CrossRef]
- Sell, C.S. On the unpredictability of odor. Angew. Chemie Int. Ed. 2006, 45, 6254–6261. [Google Scholar] [CrossRef] [PubMed]
- Ohloff, G.; Maurer, B.; Winter, B.; Giersch, W. Structural and Configurational Dependence of the Sensory Process in Steroids. Org. Biol. Chemie Chim. Org. Chim. Biol. 1983, 66, 192–217. [Google Scholar] [CrossRef]
- Su, C.-Y.; Menuz, K.; Carlson, J.R. Olfactory Perception: Receptors, Cells, and Circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfils, P. Odorat: De l’aéroportage au cortex. Bull. Acad. Natl. Med. 2014, 198, 1109–1120. [Google Scholar] [PubMed]
- Sela, L.; Sobel, N. Human olfaction: A constant state of change-blindness. Exp. Brain Res. 2010, 205, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Maresh, A.; Rodriguez Gil, D.; Whitman, M.C.; Greer, C.A. Principles of glomerular organization in the human olfactory bulb—Implications for odor processing. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Herden, G. Some aspects of qualitative data analysis. Math. Soc. Sci. 1993, 26, 105–138. [Google Scholar] [CrossRef]
- Boelens, M.H.; Boelens, H.; Van Gemert, L.J. Sensory Properties of Optical Isomers. Perfum. Flavonist 1993, 18, 1. [Google Scholar]
- Chastrette, M. Classification of Odors and Structure–Odor Relationships. In Olfaction, Taste, and Cognition; Rouby, C., Schaal, B., Dubois, D., Gervais, R., Holley, A., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 100–116. [Google Scholar]
- Ohloff, G.; Winter, B.; Fehr, C. Chemical Classification and Structure—Odour Relationships. In Perfumes; Springer Netherlands: Dordrecht, Netherlands, 1994; pp. 287–330. [Google Scholar]
- Jordan, R.; Kollo, M.; Schaefer, A.T. Sniffing Fast: Paradoxical Effects on Odor Concentration Discrimination at the Levels of Olfactory Bulb Output and Behavior. Eneuro 2018, 5. [Google Scholar] [CrossRef]
- Wilkes, F.J.; Laing, D.G.; Hutchinson, I.; Jinks, A.L.; Monteleone, E. Temporal processing of olfactory stimuli during retronasal perception. Behav. Brain Res. 2009, 200, 68–75. [Google Scholar] [CrossRef]
- Nagashima, A.; Touhara, K. Enzymatic Conversion of Odorants in Nasal Mucus Affects Olfactory Glomerular Activation Patterns and Odor Perception. J. Neurosci. 2010, 30, 16391–16398. [Google Scholar] [CrossRef] [PubMed]
- Thiebaud, N.; da Silva, S.V.; Jakob, I.; Sicard, G.; Chevalier, J.; Ménétrier, F.; Berdeaux, O.; Artur, Y.; Heydel, J.M.; Le Bon, A.M. Odorant Metabolism Catalyzed by Olfactory Mucosal Enzymes Influences Peripheral Olfactory Responses in Rats. PLoS ONE 2013, 8, 34–56. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P. Odorant-binding proteins. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 199–228. [Google Scholar] [CrossRef] [PubMed]
- Tegoni, M.; Pelosi, P.; Vincent, F.; Spinelli, S.; Campanacci, V.; Grolli, S.; Ramoni, R.; Cambillau, C. Mammalian odorant binding proteins. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2000, 1482, 229–240. [Google Scholar] [CrossRef]
- Homma, R.; Lv, X.; Sato, T.; Imamura, F.; Zeng, S.; Nagayama, S. Narrowly Confined and Glomerulus-Specific Onset Latencies of Odor-Evoked Calcium Transients in the Juxtaglomerular Cells of the Mouse Main Olfactory Bulb. Eneuro 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Kay, L.M.; Sherman, S.M. An argument for an olfactory thalamus. Trends Neurosci. 2007, 30, 47–53. [Google Scholar] [CrossRef]
- Geramita, M.; Urban, N.N. Differences in Glomerular-Layer-Mediated Feedforward Inhibition onto Mitral and Tufted Cells Lead to Distinct Modes of Intensity Coding. J. Neurosci. 2017, 37, 1428–1438. [Google Scholar] [CrossRef]
- Bizer, A.; Parabucki, A.; Smear, M.; Munoz, A.E.; Shusterman, R.; Bala, A.D.S.; Morris, G. Odor concentration change coding in the olfactory bulb. Eneuro 2019, 6. [Google Scholar] [CrossRef]
- Jaubert, J.N.; Tapiero, C.; Dore, J.C. The field of odours: Toward a universal language for odour relationships. Perfum. Flavorist 1995, 20, 1–16. [Google Scholar]
- David, S. Linguistic Expressions for Odors in French. In Olfaction, Taste, and Cognition; Rouby, C., Schaal, B., Dubois, D., Gervais, R., Holley, A., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 82–99. [Google Scholar]
- Block, E.; Jang, S.; Matsunami, H.; Sekharan, S.; Dethier, B.; Ertem, M.Z.; Gundala, S.; Pan, Y.; Li, S.; Li, Z.; et al. Implausibility of the vibrational theory of olfaction. Proc. Natl. Acad. Sci. 2015, 112, 2766–2774. [Google Scholar] [CrossRef]
- Malcolm Dyson, G. The scientific basis of odour. J. Soc. Chem. Ind. 1938, 57, 647–651. [Google Scholar] [CrossRef]
- De Jong, H.G.B.S. G.G.. model for the stimulation of organ of smell. Proc. Acad. Sci. 1937, 40, 302–306. [Google Scholar]
- Davies, J.T.; Taylor, F.H. A Model System for the Olfactory Membrane. Nature 1954, 174, 693–694. [Google Scholar] [CrossRef] [PubMed]
- Briggs, M.H.; Duncan, R.B. Odor receptors. Nature 1961, 91, 1310–1311. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, B.; Misra, T.N.; Switzer, R. Mechanism of Olfactory Transduction. Nature 1968, 217, 423–427. [Google Scholar] [CrossRef]
- Mozell, M.M. Evidence for a Chromatographic Model of Olfaction. J. Gen. Physiol. 2004, 56, 46–63. [Google Scholar] [CrossRef]
- Amoore, J.E. Specific anosmia and the concept of primary odors. Chem. Senses 1977, 2, 267–281. [Google Scholar] [CrossRef]
- Triller, A.; Boulden, E.A.; Churchill, A.; Hatt, H.; Englund, J.; Spehr, M.; Sell, C.S. Odorant–Receptor Interactions and Odor Percept: A Chemical Perspective. Chem. Biodivers. 2008, 5, 862–886. [Google Scholar] [CrossRef]
- Amoore, J.E. Stereochemical theory of olfaction. Nature 1963, 198, 271–272. [Google Scholar] [CrossRef]
- Silva Teixeira, C.S.; Cerqueira, N.M.F.S.A.; Silva Ferreira, A.C. Unravelling the Olfactory Sense: From the Gene to Odor Perception. Chem. Senses 2015, 41, 105–121. [Google Scholar] [CrossRef] [Green Version]
- DeMaria, S.; Ngai, J. The cell biology of smell. J. Cell Biol. 2010, 191, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Block, E. Molecular Basis of Mammalian Odor Discrimination: A Status Report. J. Agric. Food Chem. 2018, 66, 13346–13366. [Google Scholar] [CrossRef] [PubMed]
- McGann, J.P. Poor human olfaction is a 19th-century myth. Science 2017, 356, eaam7263. [Google Scholar] [CrossRef] [PubMed]
- Malnic, B. Searching for the ligands of odorant receptors. Mol. Neurobiol. 2007, 35, 175–181. [Google Scholar] [CrossRef] [PubMed]
- De March, C.A.; Yu, Y.; Ni, M.J.; Adipietro, K.A.; Matsunami, H.; Ma, M.; Golebiowski, J. Conserved Residues Control Activation of Mammalian G Protein- Coupled Odorant Receptors. J. Am. Chem. Soc. 2015, 137, 8611–8616. [Google Scholar] [CrossRef] [PubMed]
- De March, C.A.; Kim, S.; Antonczak, S.; Goddard, W.A., III; Golebiowski, J. G protein-coupled odorant receptors: From sequence to structure. Protein Sci. 2015, 24, 1543–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, O.; Etter, S.; Spreafico, M.; Bordoli, L.; Schwede, T.; Vogel, H.; Pick, H. The Mouse Eugenol Odorant Receptor: Structural and Functional Plasticity of a Broadly Tuned Odorant Binding Pocket. Biochemistry 2011, 50, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Man, O.; Gilad, Y.; Lancet, D. Prediction of the odorant binding site of olfactory receptor proteins by human – mouse comparisons. Protein Sci. 2004, 13, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Floriano, W.B.; Vaidehi, N.; Goddard, W.A. Making sense of olfaction through predictions of the 3-D structure and function of olfactory receptors. Chem. Senses 2004, 29, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Mohrhardt, J.; Nagel, M.; Fleck, D.; Ben-Shaul, Y.; Spehr, M. Signal detection and coding in the accessory olfactory system. Chem. Senses 2018, 43, 667–695. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.A. Pheromones and Mammalian Behavior. In The Neurobiology of Olfaction; Menini, A., Ed.; CRC Press/Taylor & Francis: Abingdon, UK, 2010. [Google Scholar]
- Gomez-Diaz, C.; Benton, R. The joy of sex pheromones. EMBO Rep. 2013, 14, 874–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milardi, D.; Colussi, C.; Grande, G.; Vincenzoni, F.; Pierconti, F.; Mancini, F.; Baroni, S.; Castagnola, M.; Marana, R.; Pontecorvi, A. Olfactory receptors in semen and in the male tract: From proteome to proteins. Front. Endocrinol. 2018, 8, 379. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, J.K.; Wilson, D.A. Human Olfaction: It Takes Two Villages. Curr. Biol. 2018, 28, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Soudry, Y.; Lemogne, C.; Malinvaud, D.; Consoli, S.M.; Bonfils, P. Olfactory system and emotion: Common substrates. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2011, 128, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menashe, I.; Abaffy, T.; Hasin, Y.; Goshen, S.; Yahalom, V.; Luetje, C.W.; Lancet, D. Genetic Elucidation of Human Hyperosmia to Isovaleric Acid. PLoS Biol. 2007, 5. [Google Scholar] [CrossRef] [PubMed]
- Croy, I.; Olgun, S.; Mueller, L.; Schmidt, A.; Muench, M.; Hummel, C.; Gisselmann, G.; Hatt, H.; Hummel, T. Peripheral adaptive filtering in human olfaction? Three studies on prevalence and effects of olfactory training in specific anosmia in more than 1600 participants. Cortex 2015, 73, 180–187. [Google Scholar] [CrossRef]
- Trimmer, C.; Keller, A.; Murphy, N.R.; Snyder, L.L.; Willer, J.R.; Nagai, M.H.; Katsanis, N.; Vosshall, L.B.; Matsunami, H.; Mainland, J.D. Genetic variation across the human olfactory receptor repertoire alters odor perception. Proc. Natl. Acad. Sci. 2019, 16, 9475–9480. [Google Scholar] [CrossRef]
- Feng, G.; Zhou, W. Nostril-specific and structure-based olfactory learning of chiral discrimination in human adults. Elife 2019, 8, e41296. [Google Scholar] [CrossRef]
- Nagappan, P.G.; Subramaniam, S.; Wang, D.Y. Olfaction as a soldier-- a review of the physiology and its present and future use in the military. Mil. Med. Res. 2017, 4, 9. [Google Scholar] [CrossRef]
- Doty, R.L.; Kamath, V. The influences of age on olfaction: A review. Front. Psychol. 2014, 5, 1–20. [Google Scholar] [CrossRef]
- Persaud, K.; Dodd, G. Analysis of discrimination mechanisms in the mammalian olfactory system using a model nose. Nature 1982, 299, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Zhu, J.; Knoll, W. From Gas Sensors to Biomimetic Artificial Noses. Chemosensors 2018, 6, 32. [Google Scholar] [CrossRef]
- Gardner, J.W.; Bartlett, P.N. A brief history of electronic noses. Sens. Actuators B Chem. 1994, 18, 210–211. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhu, J.; Knoll, W. Odorant-binding proteins as sensing elements for odour monitoring. Sensors 2018, 18, 3248. [Google Scholar] [CrossRef] [PubMed]
- Comini, E.; Faglia, G.; Sberveglieri, G.; Pan, Z.; Wang, Z.L. Stable and highly sensitive gas sensors based on semiconducting oxide nanobelts. Appl. Phys. Lett. 2002, 81, 1869–1871. [Google Scholar] [CrossRef]
- Severin, E.J.; Sanner, R.D.; Doleman, B.J.; Lewis, N.S. Differential Detection of Enantiomeric Gaseous Analytes Using Carbon Black−Chiral Polymer Composite, Chemically Sensitive Resistors. Anal. Chem. 1998, 70, 1440–1443. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.D. Review of Electronic-nose Technologies and Algorithms to Detect Hazardous Chemicals in the Environment. Procedia Technol. 2012, 1, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Wojnowski, W.; Dymerski, T.; Gębicki, J.; Namieśnik, J. Electronic Noses in Medical Diagnostics. Curr. Med. Chem. 2019, 26, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Loutfi, A.; Coradeschi, S.; Mani, G.K.; Shankar, P.; Rayappan, J.B.B. Electronic noses for food quality: A review. J. Food Eng. 2015, 144, 103–111. [Google Scholar] [CrossRef]
- Zou, G.; Xiao, Y.; Wang, M.; Zhang, H. Detection of bitterness and astringency of green tea with different taste by electronic nose and tongue. PLoS ONE 2018, 13, e0206517. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Li, Z.; Han, B.; Gholam Hosseini, H.; Zhou, H.; Wang, S.; Luo, D. Discrimination of Two Cultivars of Alpinia Officinarum Hance Using an Electronic Nose and Gas Chromatography-Mass Spectrometry Coupled with Chemometrics. Sensors 2019, 19, 572. [Google Scholar] [CrossRef] [PubMed]
- Chastrette, M.; Cretin, D.; El Aïdi, C. Structure−Odor Relationships: Using Neural Networks in the Estimation of Camphoraceous or Fruity Odors and Olfactory Thresholds of Aliphatic Alcohols. J. Chem. Inf. Comput. Sci. 1996, 36, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Cherqaoui, D.; Esseffar, M.; Villemin, D.; Cense, J.-M.; Chastrette, M.; Zakarya, D. Structure-musk odour relationship studies of tetralin and indan compounds using neural networks. New J. Chem. 1998, 22, 839–843. [Google Scholar] [CrossRef]
- Wailzer, B.; Klocker, J.; Buchbauer, G.; Ecker, G.; Wolschann, P. Prediction of the Aroma Quality and the Threshold Values of Some Pyrazines Using Artificial Neural Networks. J. Med. Chem. 2001, 44, 2805–2813. [Google Scholar] [CrossRef] [PubMed]
- Gabler, S.; Soelter, J.; Hussain, T.; Sachse, S.; Schmuker, M. Physicochemical vs. Vibrational Descriptors for Prediction of Odor Receptor Responses. Mol. Inform. 2013, 32, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Lötsch, J.; Kringel, D.; Hummel, T. Machine Learning in Human Olfactory Research. Chem. Senses 2019, 44, 11–22. [Google Scholar] [CrossRef]
- Giacalone, D.; Degn, T.K.; Yang, N.; Liu, C.; Fisk, I.; Münchow, M. Common roasting defects in coffee: Aroma composition, sensory characterization and consumer perception. Food Qual. Prefer. 2019, 71, 463–474. [Google Scholar] [CrossRef] [Green Version]
- Sáenz-Navajas, M.-P.; Arias, I.; Ferrero-del-Teso, S.; Fernández-Zurbano, P.; Escudero, A.; Ferreira, V. Chemo-sensory approach for the identification of chemical compounds driving green character in red wines. Food Res. Int. 2018, 109, 138–148. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genva, M.; Kenne Kemene, T.; Deleu, M.; Lins, L.; Fauconnier, M.-L. Is It Possible to Predict the Odor of a Molecule on the Basis of its Structure? Int. J. Mol. Sci. 2019, 20, 3018. https://doi.org/10.3390/ijms20123018
Genva M, Kenne Kemene T, Deleu M, Lins L, Fauconnier M-L. Is It Possible to Predict the Odor of a Molecule on the Basis of its Structure? International Journal of Molecular Sciences. 2019; 20(12):3018. https://doi.org/10.3390/ijms20123018
Chicago/Turabian StyleGenva, Manon, Tierry Kenne Kemene, Magali Deleu, Laurence Lins, and Marie-Laure Fauconnier. 2019. "Is It Possible to Predict the Odor of a Molecule on the Basis of its Structure?" International Journal of Molecular Sciences 20, no. 12: 3018. https://doi.org/10.3390/ijms20123018
APA StyleGenva, M., Kenne Kemene, T., Deleu, M., Lins, L., & Fauconnier, M.-L. (2019). Is It Possible to Predict the Odor of a Molecule on the Basis of its Structure? International Journal of Molecular Sciences, 20(12), 3018. https://doi.org/10.3390/ijms20123018