Heavy Metal Pollutions: State of the Art and Innovation in Phytoremediation

 ,
,  and
and 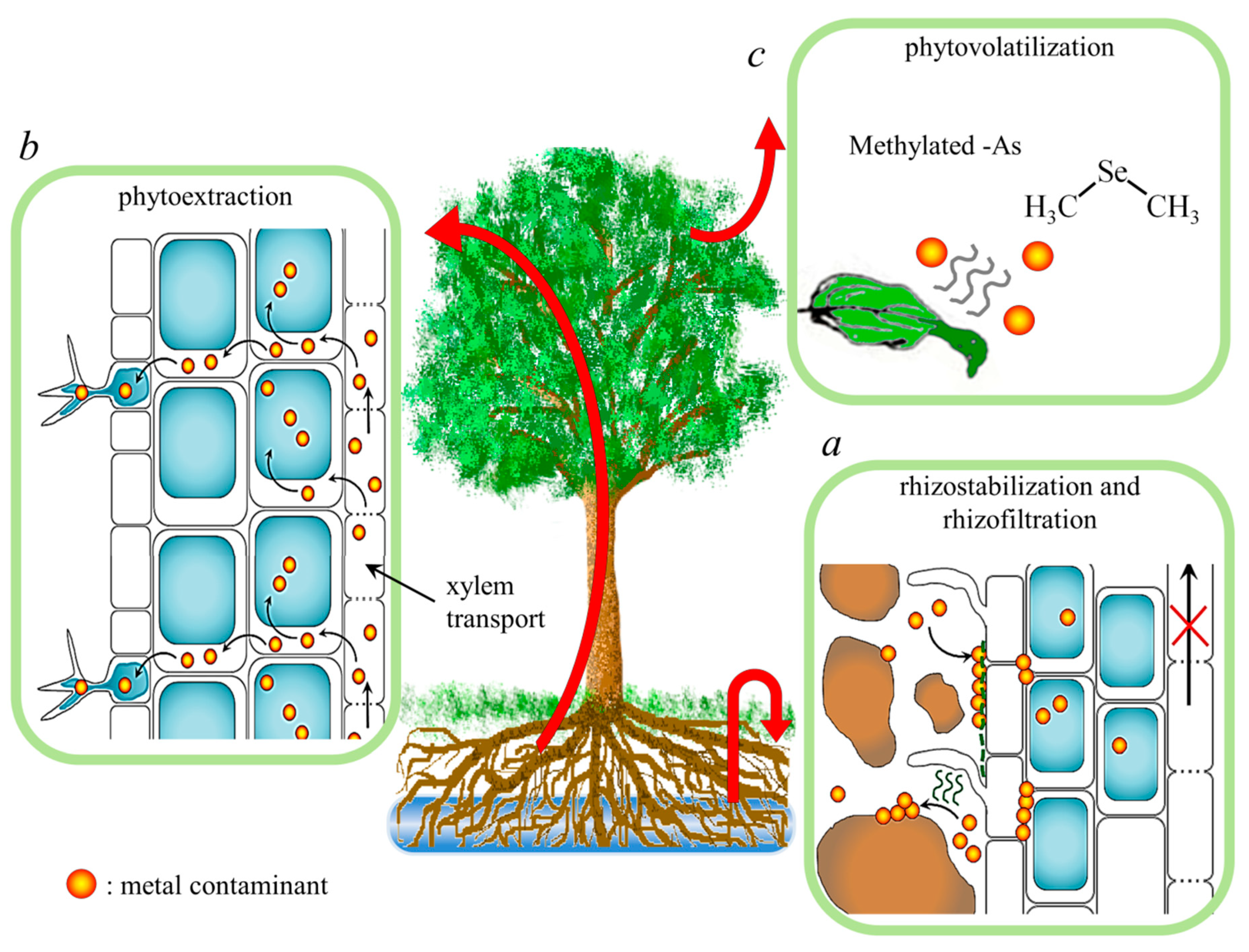{kind=link}
{kind=link}
Abstract
1. Introduction
2. Plant Mechanisms for Heavy Metal Tolerance and Detoxification
3. Phytoremediation
3.1. Strategies for Phytoremediation
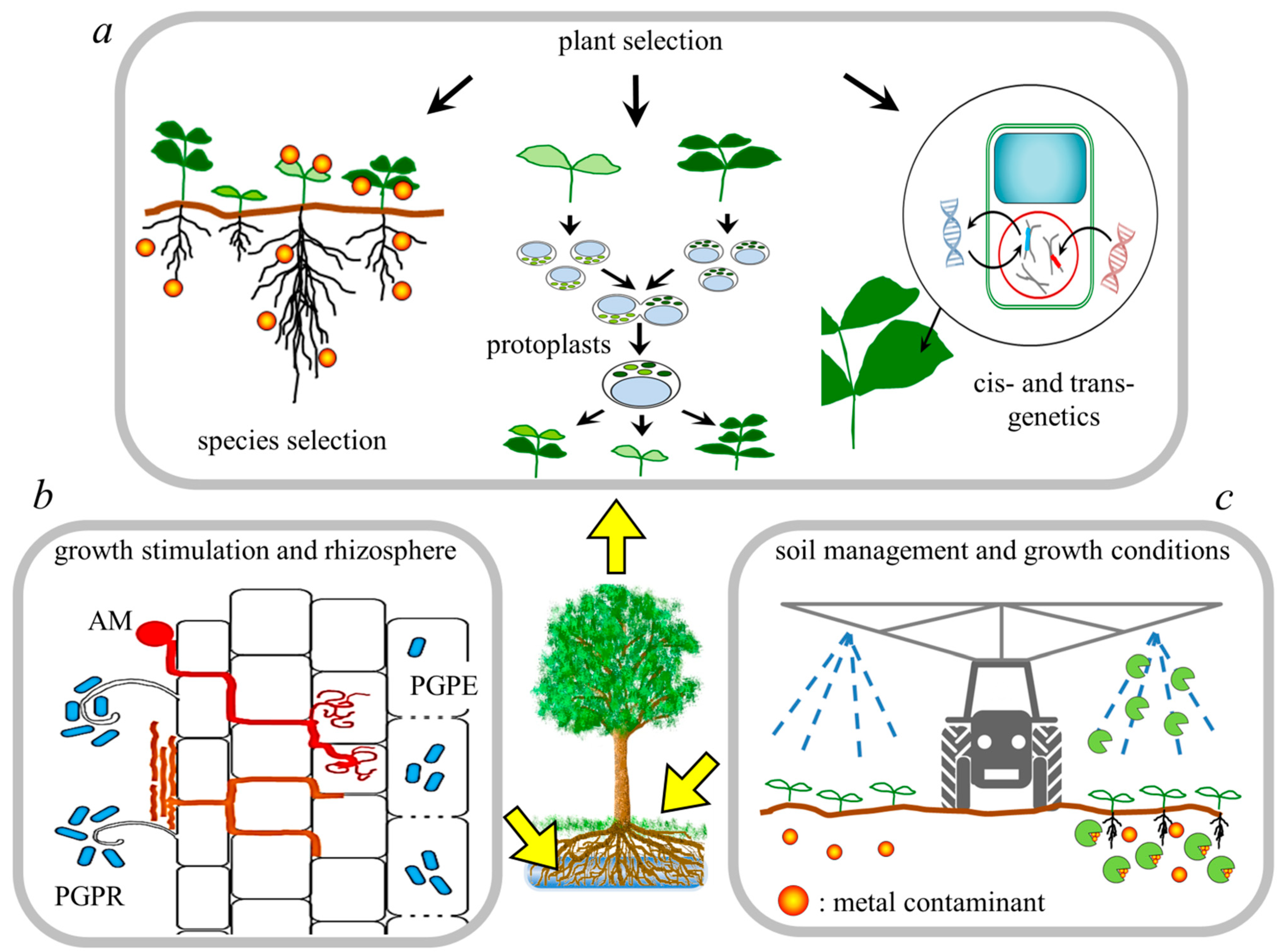
3.2. Advancement in the Field of Phytoremediation
3.2.1. Choosing the Best Plant Genotype
3.2.2. Changing the Growth Conditions
3.2.3. Enhancing the Plant-Microorganism Interactions
4. Conclusions
Conflicts of Interest
References
- Baxter, I. Ionomics: Studying the social network of mineral nutrients. Curr. Opin. Plant Biol. 2009, 12, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Krämer, U.; Talke, I.N.; Hanikenne, M. Transition metal transport. FEBS Lett. 2007, 581, 2263–2272. [Google Scholar] [CrossRef] [PubMed]
- DalCorso, G.; Manara, A.; Furini, A. An overview of heavy metal challenge in plants: From roots to shoots. Metallomics 2013, 5, 1117–1132. [Google Scholar] [CrossRef] [PubMed]
- Boughriet, A.; Proix, N.; Billon, G.; Recourt, P.; Ouddane, B. Environmental impacts of heavy metal discharges from a smelter in Deûle-canal sediments (Northern France): Concentration levels and chemical fractionation. Water Air Soil Pollut. 2007, 180, 83–95. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press Inc.: San Diego, CA, USA, 2012; pp. 191–248. [Google Scholar]
- Krämer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Gustin, J.L.; Zanis, M.J.; Salt, D.E. Structure and evolution of the plant cation diffusion facilitator family of ion transporters. BMC Evol. Biol. 2011, 11, 76–89. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Physiological limits to zinc biofortification of edible crops. Front. Plant Sci. 2011, 2, 80. [Google Scholar] [CrossRef] [PubMed]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta 2000, 1465, 190–198. [Google Scholar] [CrossRef]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef]
- Cailliatte, R.; Schikora, A.; Briat, J.F.; Mari, S.; Curie, C. High-affinity manganese uptake by the metal transporter NRAMP1 is essential for Arabidopsis growth in low manganese conditions. Plant Cell 2010, 22, 904–917. [Google Scholar] [CrossRef]
- Bastow, E.L.; de la Torre, V.S.G.; Maclean, A.E.; Green, R.T.; Merlot, S.; Thomine, S.; Balk, J. Vacuolar iron stores gated by NRAMP3 and NRAMP4 are the primary source of iron in germinating seeds. Plant Physiol. 2018, 177, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.Y.; Choi, H.; Segami, S.; Cho, H.T.; Martinoia, E.; Maeshima, M.; Lee, Y. AtHMA1contributes to the detoxification of excess Zn(II) in Arabidopsis. Plant J. 2009, 58, 737–753. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.E.; Mills, R.F. P(1B)-ATPases—An ancient family of transition metal pumps with diverse functions in plants. Trends Plant. Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Hanikenne, M.; Baurain, D. Origin and evolution of metal P-type ATPases in Plantae (Archaeplastida). Front. Plant Sci. 2014, 4, 544. [Google Scholar] [CrossRef] [PubMed]
- Desbrosses-Fonrouge, A.G.; Voigt, K.; Schröder, A.; Arrivault, S.; Thomine, S.; Krämer, U. Arabidopsis thaliana MTP1 is a Zn transporter in the vacuolar membrane which mediates Zn detoxification and drives leaf Zn accumulation. FEBS Lett. 2005, 579, 4165–4174. [Google Scholar] [CrossRef] [PubMed]
- Dalvi, A.A.; Bhalerao, S.A. Response of plants towards heavy metal toxicity: An overview of avoidance, tolerance and uptake mechanism. Ann. Plant Sci. 2013, 2, 362–368. [Google Scholar]
- Manara, A. Plant Responses to Heavy Metal Toxicity. In Plants and Heavy Metals; Furini, A., Ed.; Springer: Cham, Switzerland, 2012; pp. 27–53. [Google Scholar]
- Rai, V.K. Role of amino acids in plant responses to stress. Biol. Plant. 2002, 45, 481–487. [Google Scholar] [CrossRef]
- Howden, R.; Goldsbrough, P.B.; Andersen, C.R.; Cobbett, C.S. Cadmium-sensitive, cad1 mutants of Arabidopsis thaliana are phytochelatin deficient. Plant Physiol. 1995, 107, 1059–1066. [Google Scholar] [CrossRef]
- Kohler, A.; Blaudez, D.; Chalot, M.; Martin, F. Cloning and expression of multiple metallothioneins from hybrid poplar. New Phytol. 2004, 164, 83–93. [Google Scholar] [CrossRef]
- Guo, J.; Xu, L.; Su, Y.; Wang, H.; Gao, S.; Xu, J.; Que, Y. ScMT2-1-3, A metallothionein gene of sugarcane, plays an important role in the regulation of heavy metal tolerance/accumulation. BioMed Res. Int. 2013, 2013, 904769. [Google Scholar] [CrossRef]
- Huang, H.; Gupta, D.K.; Tian, S.; Yang, X.; Li, T. Lead tolerance and physiological adaptation mechanism in roots of accumulating and non-accumulating ecotypes of Sedum alfredii. Environ. Sci. Pollut. Res. 2012, 19, 1640–1651. [Google Scholar] [CrossRef] [PubMed]
- Wijnhoven, S.; Leuven, R.S.E.W.; van der Velde, G.; Jungheim, G.; Koelemij, E.I.; de Vries, F.T.; Eijsackers, H.J.P.; Smits, A.J.M. Heavy-metal concentrations in small mammals from a diffusely polluted floodplain: Importance of species- and location-specific characteristics. Arch. Environ. Contam. Toxicol. 2007, 52, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Ensley, B.D. Rationale for use of phytoremediation. In Phytoremediation of Toxic Metals: Using Plants to Clean up the Environment; Raskin, I., Ensley, B.D., Eds.; Wiley: New York, NY, USA, 2000; pp. 3–11. [Google Scholar]
- Fasani, E.; Manara, A.; Martini, F.; Furini, A.; DalCorso, G. The potential of genetic engineering of plants for the remediation of soils contaminated with heavy metals. Plant Cell Environ. 2018, 41, 1201–1232. [Google Scholar] [CrossRef] [PubMed]
- Fasani, E. Plants that hyperaccumulate heavy metals. In Plants and Heavy Metals; Furini, A., Ed.; Springer: Cham, Switzerland, 2012; pp. 55–74. [Google Scholar]
- Mertens, J.; Van Nevel, L.; De Schrijver, A.; Piesschaert, F.; Oosterbaan, A.; Tack, F.M.; Verheyen, K. Tree species effect on the redistribution of soil metals. Environ. Pollut. 2007, 149, 173–181. [Google Scholar] [CrossRef]
- Dickinson, N.M.; Baker, A.J.; Doronila, A.; Laidlaw, S.; Reeves, R.D. Phytoremediation of inorganics: Realism and synergies. Int. J. Phytoremed. 2009, 11, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.H.; Anderson, C.W.N.; Dickinson, N.M. Phytoextraction: Where’s the action? J. Geochem. Explor. 2015, 151, 34–40. [Google Scholar] [CrossRef]
- De Souza, M.P.; Lytle, C.M.; Mulholland, M.M.; Otte, M.L.; Terry, N. Selenium assimilation and volatilization from dimethylselenoniopropionate by Indian mustard. Plant Physiol. 2000, 122, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Williams, P.N.; Meharg, A.A. Exposure to inorganic arsenic from rice: A global health issue? Environ. Pollut. 2008, 154, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Li, B.; Shao, J.J.; Wang, T.; He, B.; Shi, J.B.; Ye, Z.H.; Jiang, G.B. Accumulation of total mercury and methylmercury in rice plants collected from different mining areas in China. Environ. Pollut. 2014, 184, 179–186. [Google Scholar] [CrossRef]
- Moreno, F.N.; Anderson, C.W.N.; Stewart, R.B.; Robinson, B.H. Phytofiltration of mercury-contaminated water: Volatilisation and plant-accumulation aspects. Environ. Exp. Bot. 2008, 62, 78–85. [Google Scholar] [CrossRef]
- Brunner, I.; Luster, J.; Günthardt-Goerg, M.S.; Frey, B. Heavy metal accumulation and phytostabilisation potential of tree fine roots in a contaminated soil. Environ. Pollut. 2008, 152, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Berti, W.R.; Cunningham, S.D. Phytostabilization of metals. In Phytoremediation of Toxic Metals: Using Plants to Clean up the Environment; Raskin, I., Ensley, B.D., Eds.; Wiley: New York, NY, USA, 2000; pp. 71–88. [Google Scholar]
- Mench, M.; Vangronsveld, J.; Bleeker, P.; Ruttens, A.; Geebelen, W.; Lepp, N. Phytostabilisation of metal-contaminated sites. In Phytoremediation of Metal-Contaminated Soils; Morel, J.L., Echevarria, G., Goncharova, N., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 109–190. [Google Scholar]
- Yadav, B.K.; Siebel, M.A.; Van Bruggen, J.J.A. Rhizofiltration of a Heavy Metal (Lead) Containing Wastewater Using the Wetland Plant Carex pendula. CLEAN Soil Air Water 2011, 39, 467–474. [Google Scholar] [CrossRef]
- Dushenkov, V.; Kumar, P.B.; Motto, H.; Raskin, I. Rhizofiltration: The use of plants to remove heavy metals from aqueous streams. Environ. Sci. Technol. 1995, 29, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Dushenkov, S.; Kapulnik, Y. Phytofiltration of metals. In Phytoremediation of Toxic Metals—Using Plants to Clean up the Environment; Raskin, I., Ensley, B.D., Eds.; Wiley: New York, NY, USA, 2000; pp. 89–106. [Google Scholar]
- Raskin, I.; Kumar, P.B.A.N.; Dushenkov, S.; Salt, D.E. Bioconcentration of heavy metals by plants. Curr. Opin. Biotechnol. 1994, 5, 285–290. [Google Scholar] [CrossRef]
- Salt, D.E.; Blaylock, M.; Kumar, N.P.; Dushenkov, V.; Ensley, B.D.; Chet, I.; Raskin, I. Phytoremediation: A novel strategy for the removal of toxic metals from the environment using plants. Biotechnology 1995, 13, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Raoufa, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and wastewater treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef] [PubMed]
- Molazadeh, M.; Ahmadzadeh, H.; Pourianfar, H.R.; Lyon, S.; Rampelotto, P.H. The use of microalgae for coupling wastewater treatment with CO2 biofixation. Front. Bioeng. Biotechnol. 2019, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Judd, S.; van den Broeke, L.J.; Shurair, M.; Kuti, Y.; Znad, H. Algal remediation of CO2 and nutrient discharges: A review. Water Res. 2015, 87, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Roccotiello, E.; Marescotti, P.; Di Piazza, S.; Cecchi, G.; Mariotti, M.G.; Zotti, M. Biodiversity in Metal-Contaminated Sites-Problem and Perspective—A Case Study. In Biodiversity in Ecosystems—Linking Structure and Function; Blanco, J.A., Ed.; IntechOpen: London, UK, 2015; pp. 563–582. [Google Scholar]
- Laxman Singh, K.; Sudhakar, G.; Swaminathan, S.K.; Muralidhar Rao, C. Identification of elite native plants species for phytoaccumulation and remediation of major contaminants in uranium tailing ponds and its affected area. Environ. Dev. Sustain. 2015, 17, 57–81. [Google Scholar] [CrossRef]
- Barrutia, O.; Artetxe, U.; Hernández, A.; Olano, J.M.; García-Plazaola, J.I.; Garbisu, C.; Becerril, J.M. Native plant communities in an abandoned Pb-Zn mining area of northern Spain: Implications for phytoremediation and germplasm preservation. Int. J. Phytoremed. 2011, 13, 256–270. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Hanus-Fajerska, E.; Smoleń, S.; Muszyńska, E. In vitro selection for lead tolerance in shoot culture of Daphne species. Acta Sci. Pol. Hortorum Cultus 2015, 14, 129–142. [Google Scholar]
- Dushenkov, S.; Skarzhinskaya, M.; Glimelius, K.; Gleba, D.; Raskin, I. Bioengineering of a phytoremediation plant by means of somatic hybridization. Int. J. Phytoremed. 2002, 4, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Brewer, E.P.; Saunders, J.A.; Scott, A.J.; Chaney, R.L.; McIntosh, M.S. Somatic hybridization between the zinc accumulator Thlaspi caerulescens and Brassica napus. Theor. Appl. Genet. 1999, 99, 761–771. [Google Scholar] [CrossRef]
- Lyyra, S.; Meagher, R.B.; Kim, T.; Heaton, A.; Montello, P.; Balish, R.S.; Merkle, S.A. Coupling two mercury resistance genes in Eastern cottonwood enhances the processing of organomercury. Plant Biotechnol. J. 2007, 5, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Shim, D.; Kim, S.; Choi, Y.I.; Song, W.Y.; Park, J.; Youk, E.S.; Jeong, S.C.; Martinoia, E.; Noh, E.W.; Lee, Y. Transgenic poplar trees expressing yeast cadmium factor 1 exhibit the characteristics necessary for the phytoremediation of mine tailing soil. Chemosphere 2013, 90, 1478–1486. [Google Scholar] [CrossRef] [PubMed]
- Didovyk, A.; Borek, B.; Tsimring, L.; Hasty, J. Transcriptional regulation with CRISPR-Cas9: Principles, advances, and applications. Curr. Opin. Biotechnol. 2016, 40, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Basharat, Z.; Novo, L.A.B.; Yasmin, A. Genome editing weds CRISPR: What is in it for phytoremediation? Plants 2018, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.P.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [PubMed]
- Sleegers, F.; Hisle, M. Redesigning Abandoned Gas Stations Through Phytotechnologies. In Phytoremediation; Ansari, A.A., Gill, S.S., Gill, R., Lanza, G.R., Newman, L., Eds.; Springer: Cham, Switzerland, 2018; Volume 6, pp. 3–20. [Google Scholar]
- Rezania, S.; Taib, S.M.; Din, M.F.M.; Dahalan, F.A.; Kamyab, H. Comprehensive review on phytotechnology: Heavy metals removal by diverse aquatic plants species from wastewater. J. Hazard. Mater. 2016, 318, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Newete, S.W.; Byrne, M.J. The capacity of aquatic macrophytes for phytoremediation and their disposal with specific reference to water hyacinth. Environ. Sci. Pollut. Res. 2016, 23, 10630–10643. [Google Scholar] [CrossRef] [PubMed]
- Marchand, L.; Mench, M.; Jacob, D.L.; Otte, M.L. Metal and metalloid removal in constructed wetlands, with emphasis on the importance of plants and standardized measurements: A review. Environ. Pollut. 2010, 158, 3447–3461. [Google Scholar] [CrossRef] [PubMed]
- Odinga, C.A.; Swalaha, F.M.; Otieno, F.A.O.; Ranjith, K.R.; Bux, F. Investigating the efficiency of constructed wetlands in the removal of heavy metals and enteric pathogens from wastewater. Environ. Technol. Rev. 2013, 2, 1–16. [Google Scholar] [CrossRef]
- Ladislas, S.; Gerente, C.; Chazarenc, F.; Brisson, J.; Andres, Y. Floating treatment wetlands for heavy metal removal in highway stormwater ponds. Ecol. Eng. 2015, 80, 85–91. [Google Scholar] [CrossRef]
- Ning, D.; Huang, Y.; Pan, R.; Wang, F.; Wang, H. Effect of eco-remediation using planted floating bed system on nutrients and heavy metals in urban river water and sediment: A field study in China. Sci. Total Environ. 2014, 485, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Bavandpour, F.; Zou, Y.; He, Y.; Saeed, T.; Sun, Y.; Sun, G. Removal of dissolved metals in wetland columns filled with shell grits and plant biomass. Chem. Eng. J. 2018, 331, 234–241. [Google Scholar] [CrossRef]
- Dan, A.; Oka, M.; Fujii, Y.; Soda, S.; Ishigaki, T.; Machimura, T.; Ike, M. Removal of heavy metals from synthetic landfill leachate in lab-scale vertical flow constructed wetlands. Sci. Total Environ. 2017, 584, 742–750. [Google Scholar]
- Lizama Allende, K.; Fletcher, T.D.; Sun, G. Enhancing the removal of arsenic, boron and heavy metals in subsurface flow constructed wetlands using different supporting media. Water Sci. Technol. 2011, 63, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Fasani, E.; DalCorso, G.; Zerminiani, A.; Ferrarese, A.; Campostrini, P.; Furini, A. Phytoremediatory efficiency of Chrysopogon zizanioides in the treatment of landfill leachate: A case study. Environ. Sci. Pollut. Res. 2019, 26, 10057–10069. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Bauer, E.O. Evaluation of Populus and Salix continuously irrigated with landfill leachate I. Genotype-specific elemental phytoremediation. Int. J. Phytoremed. 2007, 9, 281–306. [Google Scholar] [CrossRef]
- Aronsson, P.; Dahlin, T.; Dimitriou, I. Treatment of landfill leachate by irrigation of willow coppice—Plant response and treatment efficiency. Environ. Pollut. 2010, 158, 795–804. [Google Scholar] [CrossRef]
- Coyle, D.R.; Zalesny, J.A.; Zalesny, R.S., Jr.; Wiese, A.H. Irrigating poplar energy crops with landfill leachate negatively affects soil micro-and meso-fauna. Int. J. Phytoremed. 2011, 13, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Burges, A.; Alkorta, I.; Epelde, L.; Garbisu, C. From phytoremediation of soil contaminants to phytomanagement of ecosystem services in metal contaminated sites. Int. J. Phytoremed. 2018, 20, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Burges, A.; Epelde, L.; Benito, G.; Artetxe, U.; Becerril, J.M.; Garbisu, C. Enhancement of ecosystem services during endophyte-assisted aided phytostabilization of metal contaminated mine soil. Sci. Total Environ. 2016, 562, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Karczewska, A.; Gałka, B.; Dradrach, A.; Lewińska, K.; Mołczan, M.; Cuske, M.; Gersztyn, L.; Litak, K. Solubility of arsenic and its uptake by ryegrass from polluted soils amended with organic matter. J. Geochem. Explor. 2017, 182, 193–200. [Google Scholar] [CrossRef]
- Alvarenga, P.; Gonçalves, A.P.; Fernandes, R.M.; De Varennes, A.; Vallini, G.; Duarte, E.; Cunha-Queda, A.C. Organic residues as immobilizing agents in aided phytostabilization: (I) Effects on soil chemical characteristics. Chemosphere 2009, 74, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Duo, L.A.; Lian, F.; Zhao, S.L. Enhanced uptake of heavy metals in municipal solid waste compost by turfgrass following the application of EDTA. Environ. Monit. Assess. 2010, 165, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Sung, K. Effects of chelates on soil microbial properties, plant growth and heavy metal accumulation in plants. Ecol. Eng. 2014, 73, 386–394. [Google Scholar] [CrossRef]
- Zhao, S.; Jia, L.; Duo, L. The use of a biodegradable chelator for enhanced phytoextraction of heavy metals by Festuca arundinacea from municipal solid waste compost and associated heavy metal leaching. Bioresour. Technol. 2013, 129, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Attinti, R.; Barrett, K.R.; Datta, R.; Sarkar, D. Ethylenediaminedisuccinic acid (EDDS) enhances phytoextraction of lead by vetiver grass from contaminated residential soils in a panel study in the field. Environ. Pollut. 2017, 225, 524–533. [Google Scholar] [CrossRef]
- Ehsan, S.; Ali, S.; Noureen, S.; Mahmood, K.; Farid, M.; Ishaque, W.; Shakoor, M.B.; Rizwan, M. Citric acid assisted phytoremediation of cadmium by Brassica napus L. Ecotoxicol. Environ. Saf. 2014, 106, 164–172. [Google Scholar] [CrossRef]
- Aderholt, M.; Vogelien, D.L.; Koether, M.; Greipsson, S. Phytoextraction of contaminated urban soils by Panicum virgatum L. enhanced with application of a plant growth regulator (BAP) and citric acid. Chemosphere 2017, 175, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Fässler, E.; Evangelou, M.W.; Robinson, B.H.; Schulin, R. Effects of indole-3-acetic acid (IAA) on sunflower growth and heavy metal uptake in combination with ethylene diamine disuccinic acid (EDDS). Chemosphere 2010, 80, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Vamerali, T.; Bandiera, M.; Hartley, W.; Carletti, P.; Mosca, G. Assisted phytoremediation of mixed metal (loid)-polluted pyrite waste: Effects of foliar and substrate IBA application on fodder radish. Chemosphere 2011, 84, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Hadi, F.; Bano, A.; Fuller, M.P. The improved phytoextraction of lead (Pb) and the growth of maize (Zea mays L.): The role of plant growth regulators (GA3 and IAA) and EDTA alone and in combinations. Chemosphere 2010, 80, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Cameselle, C.; Gouveia, S. Phytoremediation of mixed contaminated soil enhanced with electric current. J. Hazard. Mater. 2019, 361, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Han, F.X.; Shao, X.; Guo, K.; McComb, J.; Arslan, Z.; Zhang, Z. Electro-kinetic remediation coupled with phytoremediation to remove lead, arsenic and cesium from contaminated paddy soil. Ecotoxicol. Environ. Saf. 2016, 125, 16–24. [Google Scholar] [CrossRef]
- Luo, J.; Yang, D.; Qi, S.; Wu, J.; Gu, X.S. Using solar cell to phytoremediate field-scale metal polluted soil assisted by electric field. Ecotox. Environ. Saf. 2018, 165, 404–410. [Google Scholar] [CrossRef]
- Mueller, N.C.; Nowack, B. Nanoparticles for remediation: Solving big problems with little particles. Elements 2010, 6, 395–400. [Google Scholar] [CrossRef]
- Liang, S.X.; Jin, Y.; Liu, W.; Li, X.; Shen, S.G.; Ding, L. Feasibility of Pb phytoextraction using nano-materials assisted ryegrass: Results of a one-year field-scale experiment. J. Environ. Manag. 2017, 190, 170–175. [Google Scholar] [CrossRef]
- Moameri, M.; Khalaki, M.A. Capability of Secale montanum trusted for phytoremediation of lead and cadmium in soils amended with nano-silica and municipal solid waste compost. Environ. Sci. Pollut. R. 2017, 1–8. [Google Scholar] [CrossRef]
- Singh, J.; Lee, B.K. Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max): A possible mechanism for the removal of Cd from the contaminated soil. J. Environ. Manag. 2016, 170, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.W.; Fallman, K.; Puschenreiter, M. The role of plant-associated bacteria in the mobilization and phytoextraction of trace elements in contaminated soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.E.D.; Boon, E.; St-Arnaud, M.; Hijri, M. Molecular biodiversity of arbuscular mycorrhizal fungi in trace metal-polluted soils. Mol. Ecol. 2011, 20, 3469–3483. [Google Scholar] [CrossRef] [PubMed]
- Ker, K.; Charest, C. Nickel remediation by AM-colonized sunflower. Mycorrhiza 2010, 20, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Enkhtuya, B.; Rydlová, J.; Vosátka, M. Effectiveness of indigenous and non-indigenous isolates of arbuscular mycorrhizal fungi in soils from degraded ecosystems and man-made habitats. Appl. Soil Ecol. 2002, 14, 201–211. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Prieto-Fernández, Á.; Álvarez-López, V.; Cabello-Conejo, M.I.; Acea, M.J.; Kidd, P.S. Nickel solubilizing capacity and characterization of rhizobacteria isolated from hyperaccumulating and non-hyperaccumulating subspecies of Alyssum serpyllifolium. Int. J. Phytoremed. 2011, 13, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Glick, B.R. Plant growth-promoting bacteria and metals phytoremediation. In Phytotechnologies: Remediation of Environmental Contaminants; Anjum, N.A., Pereira, M.E., Ahmad, I., Duarte, A.C., Umar, S., Khan, N.A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 361–376. [Google Scholar]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.V.; Singh, N.; Behl, H.M.; Srivastava, S. Influence of plant growth promoting bacteria and its mutant on heavy metal toxicity in Brassica juncea grown in fly ash amended soil. Chemosphere 2008, 72, 678–683. [Google Scholar] [CrossRef]
- Glick, B.R. Using soil bacteria to facilitate phytoremediation. Biotechnol. Adv. 2010, 28, 367–374. [Google Scholar] [CrossRef]
- Arshad, M.; Saleem, M.; Hussain, S. Perspectives of bacterial ACC deaminase in phytoremediation. Trends Biotechnol. 2007, 25, 356–362. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Luo, Y.; Freitas, H. Inoculation of endophytic bacteria on host and non-host plants—Effects on plant growth and Ni uptake. J. Hazard. Mater. 2011, 195, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Rajkumar, M.; Freitas, H. Isolation and characterization of Ni mobilizing PGPB from serpentine soils and their potential in promoting plant growth and Ni accumulation by Brassica spp. Chemosphere 2009, 75, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.F.; Xia, J.J.; Jiang, C.Y.; He, L.Y.; Qian, M. Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.Y.; Sheng, X.F.; Qian, M.; Wang, Q.Y. Isolation and characterization of a heavy metal resistant Burkholderia sp. from heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal polluted soil. Chemosphere 2008, 72, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; Vamerali, T.; Mattarozzi, M.; Dramis, L.; Sanangelantoni, A.M. Combined endophytic inoculants enhance nickel phytoextraction from serpentine soil in the hyperaccumulator Noccaea caerulescens. Front. Plant Sci. 2015, 6, 638. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Freitas, H. Influence of metal resistant-plant growth promoting bacteria on the growth of Ricinus communis in soil contaminated with heavy metals. Chemosphere 2008, 71, 834–842. [Google Scholar] [CrossRef]
- He, H.; Ye, Z.; Yang, D.; Yan, J.; Xiao, L.; Zhong, T.; Yuan, M.; Cai, X.; Fang, Z.; Jing, Y. Characterization of endophytic Rahnella sp. JN6 from Polygonum pubescens and its potential in promoting growth and Cd, Pb, Zn uptake by Brassica napus. Chemosphere 2013, 90, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Mesa, V.; Navazas, A.; González-Gil, R.; González, A.; Weyens, N.; Lauga, B.; Gallego, J.L.R.; Sánchez, J.; Peláez, A. Use of endophytic and rhizosphere bacteria to improve phytoremediation of arsenic-contaminated industrial soils by autochthonous Betula celtiberica. Appl. Environ. Microbiol. 2017, 83, e03411-16. [Google Scholar] [CrossRef] [PubMed]
- Kaldorf, M.; Kuhn, A.J.; Schroder, W.H.; Hildebrandt, U.; Bothe, H. Selective element deposits in maize colonized by a heavy metal tolerance conferring arbuscular mycorrhizal fungus. J. Plant Physiol. 1999, 154, 718–728. [Google Scholar] [CrossRef]
- Tonin, C.; Vandenkoornhuyse, P.; Joner, E.J.; Straczek, J.; Leyval, C. Assessment of arbuscular mycorrhizal fungi diversity in the rhizosphere of Viola calaminaria and effect of these fungi on heavy metal uptake by clover. Mycorrhiza 2001, 10, 161–168. [Google Scholar] [CrossRef]
- Pham, V.J.T.; Kim, J. Cultivation of unculturable soil bacteria. Trends Biotechnol. 2012, 30, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Mattarozzi, M.; Manfredi, M.; Montanini, B.; Gosetti, F.; Sanangelantoni, A.M.; Marengo, E.; Careri, M.; Visioli, G. A metaproteomic approach dissecting major bacterial functions in the rhizosphere of plants living in serpentine soil. Anal. Bioanal. Chem. 2017, 409, 2327–2339. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Sanschagrin, S.; Maynard, C.; St-Arnaud, M.; Greer, C.W. Microbial expression profiles in the rhizopshere of willows depend on soil contamination. ISME J. 2014, 8, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.H.; Joly, S.; Pitre, F.E.; Yergeau, E. Increasing phytoremediation efficiency and reliability using novel omics approaches. Trends Biotechnol. 2014, 32, 271–280. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DalCorso, G.; Fasani, E.; Manara, A.; Visioli, G.; Furini, A. Heavy Metal Pollutions: State of the Art and Innovation in Phytoremediation. Int. J. Mol. Sci. 2019, 20, 3412. https://doi.org/10.3390/ijms20143412
DalCorso G, Fasani E, Manara A, Visioli G, Furini A. Heavy Metal Pollutions: State of the Art and Innovation in Phytoremediation. International Journal of Molecular Sciences. 2019; 20(14):3412. https://doi.org/10.3390/ijms20143412
Chicago/Turabian StyleDalCorso, Giovanni, Elisa Fasani, Anna Manara, Giovanna Visioli, and Antonella Furini. 2019. "Heavy Metal Pollutions: State of the Art and Innovation in Phytoremediation" International Journal of Molecular Sciences 20, no. 14: 3412. https://doi.org/10.3390/ijms20143412
APA StyleDalCorso, G., Fasani, E., Manara, A., Visioli, G., & Furini, A. (2019). Heavy Metal Pollutions: State of the Art and Innovation in Phytoremediation. International Journal of Molecular Sciences, 20(14), 3412. https://doi.org/10.3390/ijms20143412