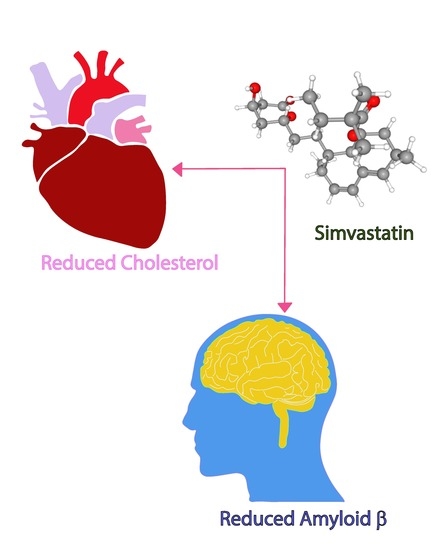
Simvastatin Efficiently Reduces Levels of Alzheimer’s Amyloid Beta in Yeast



Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Older Cells Do Not Degrade Aβ42 Fused to GFP Effectively
2.2. Statins Reduce Levels of Aβ42 Fused to GFP
2.3. Is Reduction of Aβ Fused to GFP Related to Inhibition of Ergosterol Synthesis?
2.4. Expression of Native Aβ42 Using pYEX-BX
2.5. Simvastatin Reduces Levels of Native Aβ42 Expressed in Saccharomyces cerevisiae
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Growth
4.2. Hydrolysis of Statins
4.3. Effect of Statins and Fluconazole on Yeast Cells
4.4. Flow Cytometry
4.5. Native Aβ42 Vector Construction
4.6. Transformation of Plasmids into S. cerevisiae
4.7. Quantitative MALDI-TOF Mass Spectrometry to Detect the Levels of Aβ42
4.7.1. Growth of Transformants Expressing Native Aβ42
4.7.2. Sample Preparation for MALDI-TOF
4.7.3. Measurement of Relative Quantity of Aβ42 Using MALDI-TOF
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| Aβ42 | β-amyloid of 42 amino acids |
| BACE | Beta-secretase enzymes |
| DMSO | Dimethyl sulphoxide |
| GFP | Green fluorescent protein |
| HCCA | 2-Cyano-3-(4-hydroxyphenyl) acrylic acid |
| IPOD | Insoluble protein deposit |
| MALDI | Matrix Assisted Laser Desorption Ionization |
| PBS | Phosphate buffered saline |
| TBS | Tris buffered saline |
| TBS-T | Tris buffered saline with 0.05% Tween 20 |
| TOF | Time of flight |
| μM | Micromolar |
References
- Wolozin, B.; Wang, S.W.; Li, N.C.; Lee, A.; Lee, T.A.; Kazis, L.E. Simvastatin is associated with a reduced incidence of dementia and Parkinson’s disease. BMC Med. 2007, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Wang, G.; Wang, Y.; Han, D.; Bi, J.; Yuan, Y.; Yao, T.; Wan, Z.; Li, H.; Mei, X. Neuroprotective effect of simvastatin via inducing the autophagy on spinal cord injury in the rat Model. BioMed Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Parsons, R.B.; Price, G.C.; Farrant, J.K.; Subramaniam, D.; Adeagbo-Sheikh, J.; Austen, B.M. Statins inhibit the dimerization of β-secretase via both isoprenoid and cholesterol-mediated mechanisms. Biochem. J. 2006, 399, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.L.; Aneta, G.; Ingrid, O.; Manhart, B.L.; Kelly, H.O.; Robert, V. Statins cause intracellular accumulation of amyloid precursor protein, β-secretase-cleaved fragments, and amyloid β-peptide via an isoprenoid-dependent mechanism. J. Biol. Chem. 2005, 280, 18755–18770. [Google Scholar] [CrossRef] [PubMed]
- Sjogren, M.; Michelle, M.D.; Gustafsonc, P.Z.; Ingmar, S. Cholesterol and Alzheimer’s disease-Is there a relation? Mech. Ageing Dev. 2006, 127, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Wolozin, B.; Manger, J.; Bryant, R.; Cordy, J.; Green, R.C.; McKee, A. Cholesterol, Alzheimer’s disease and statins: Re-assessing the relationship between cholesterol, statins and Alzheimer’s disease. Acta Neuropathol. Scand. Suppl. 2006, 185, 63–70. [Google Scholar] [CrossRef]
- Fassbender, K.; Simons, M.; Bergmann, C.; Stroick, M.; Lütjohann, D.; Keller, P.; Runz, H. Simvastatin strongly reduces levels of Alzheimer’s disease β-amyloid peptides Aβ42 and Aβ40 in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2001, 98, 5856–5861. [Google Scholar] [CrossRef]
- Wahrle, S.; Pritam, D.; Andrew, C.N.; Chris, M.; Mikio, S.; Takeshi, K.; Linda, H. Cholesterol-dependent γ-secretase activity in buoyant cholesterol-rich membrane. Neurobiol. Dis. 2002, 9, 11–23. [Google Scholar] [CrossRef]
- Cordy, J.M.; Hussain, I.; Dingwall, C.; Hooper, N.M.; Turner, A.J. Exclusively targeting β-secretase to lipid rafts by GPI-anchor addition up-regulates β-site processing of the amyloid precursor protein. Proc. Natl. Acad. Sci. USA 2003, 100, 11735–11740. [Google Scholar] [CrossRef]
- Refolo, L.M.; Malester, B.; LaFrancois, J.; Bryant-Thomas, T.; Wang, R.; Tint, G.S.; Sambamurti, K.; Duff, K.; Pappolla, M.A. Hypercholesterolemia accelerates the Alzheimer’s amyloid pathology in a transgenic mouse model. Neurobiol. Dis. 2000, 7, 321–331. [Google Scholar] [CrossRef]
- Shie, F.S.; Jin, L.W.; Cook, D.G.; Leverenz, J.B.; LeBoeuf, R.C. Diet-induced hypercholesterolemia enhances brain Aβ accumulation in transgenic mice. Neuroreport 2002, 13, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, K.; Barrett-Connor, E.; Lin, F.; Grady, D. Serum lipoprotein levels, statin use and cognitive function in older women. Arch. Neurol. 2002, 59, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Friedhoff, L.T.; Edward, C.; Neil, S.M.G.; Joseph, D.B. Treatment with controlled-release lovastatin decreases serum concentrations of human β-amyloid (Aβ) peptide. Int. J. Neuropsychopharmacol. 2001, 4, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Sparks, D.L.; Sabbagh, M.N.; Connor, D.J.; Lopez, J.; Launer, L.J.; Browne, P.; Wasser, D.; Johnson-Traver, S.; Lochhead, J.; Ziolwolski, C. Atorvastatin for the treatment of mild to moderate Alzheimer disease: Preliminary results. Arch. Neurol. 2005, 62, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Porzoor, A.; Macreadie, I. Yeast as a model for studies on Aβ aggregation toxicity in Alzheimer’s disease, autophagic responses, and drug screening. In Systems Biology of Alzheimer’s Disease; Springer: Berlin/Heidelberg, Germany, 2016; pp. 217–226. [Google Scholar]
- Caine, J.; Sankovich, S.; Antony, H.; Waddington, L.; Macreadie, P.; Varghese, J.; Macreadie, I. Alzheimer’s Aβ fused to green fluorescent protein induces growth stress and a heat shock response. FEMS Yeast Res. 2007, 7, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Westermeyer, C.; Macreadie, I.G. Simvastatin reduces ergosterol levels, inhibits growth and causes loss of mtDNA in Candida glabrata. FEMS Yeast Res. 2007, 7, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Macreadie, I.G.; Failla, P.; Horaitis, O.; Azad, A.A. Production of HIV-1 Vpu with pYEULCBX, a convenient vector for the production of non-fused proteins in yeast. Biotechnol. Lett. 1992, 14, 639–642. [Google Scholar] [CrossRef]
- Macreadie, I.G. Yeasts, Molecular and Therapeutic Applications. In Kirk-Othmer Encyclopedia of Chemical Technology; Wiley: Hoboken, NJ, USA, 2017; pp. 1–24. [Google Scholar]
- Macreadie, I.G.; Johnson, G.; Schlosser, T.; Macreadie, P.I. Growth inhibition of Candida species and Aspergillus fumigatus by statins. FEMS Microbiol. Lett. 2006, 262, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Vilimanovich, U.; Bosnjak, M.; Bogdanovic, A.; Markovic, I.; Isakovic, A.; Kravic-Stevovic, T.; Mircic, A.; Trajkovic, V.; Bumbasirevic, V. Statin-mediated inhibition of cholesterol synthesis induces cytoprotective autophagy in human leukemic cells. Eur. J. Pharmacol. 2015, 765, 415–428. [Google Scholar] [CrossRef]
- Bharadwaj, P.R.; Verdile, G.; Barr, R.K.; Gupta, V.; Steele, J.W.; Lachenmayer, M.L.; Martins, R.N. Latrepirdine (Dimebon (™)) enhances autophagy and reduces intracellular GFP-Aβ42 levels in yeast. J. Alzheimer’s Dis. 2012, 32, 949–967. [Google Scholar] [CrossRef]
- Nair, S.; Traini, M.; Dawes, I.W.; Perroned, G.G. Genome-wide analysis of Saccharomyces cerevisiae identifies cellular processes affecting intracellular aggregation of Alzheimer’s amyloid-β42: Importance of lipid homeostasis. Mol. Biol. Cell 2014, 25, 2235–2249. [Google Scholar] [CrossRef] [PubMed]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.C.; Mett, J.; Weingärtner, O.; Laufs, U.; et al. Plant sterols the better cholesterol in Alzheimer’s disease? A mechanistical study. J. Neurosci. 2013, 33, 1607287. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.F.; Domenico, D.; Butterfield, D.A. Statins more than cholesterol lowering agents in Alzheimer disease: Their pleiotropic functions as potential therapeutic targets. Biochem. Pharmacol. 2014, 88, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Shepardson, N.E.; Shankar, G.M.; Selkoe, D.J. Cholesterol and statins in Alzheimer’s Disease: I. Review of epidemiological and preclinical studies. Arch. Neurol. 2011, 68, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, B.; Mousavi, B.; Macreadie, I.G. Yeast model of amyloid-β and tau aggregation in Alzheimer’s disease. J. Alzheimer’s Dis. 2015, 47, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Antony, H. Study of Cellular Responses and Protein Interactions of Aβ in Yeast models. Ph.D. Thesis, RMIT University, Melbourne, Australia, March 2008. Available online: https://researchbank.rmit.edu.au/eserv/rmit:8692/Antony.pdf (accessed on 18 July 2019).
- Lorenz, R.; Parks, L. Effects of lovastatin (mevinolin) on sterol levels and on activity of azoles in Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 1990, 34, 1660–1665. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.W.; Roder, H.; Hunsucker, S.W. Quantitative matrix-assisted laser desorption/ionization mass spectrometry. Brief. Funct. Genom. Proteomics 2008, 7, 355–370. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhakal, S.; Subhan, M.; Fraser, J.M.; Gardiner, K.; Macreadie, I. Simvastatin Efficiently Reduces Levels of Alzheimer’s Amyloid Beta in Yeast. Int. J. Mol. Sci. 2019, 20, 3531. https://doi.org/10.3390/ijms20143531
Dhakal S, Subhan M, Fraser JM, Gardiner K, Macreadie I. Simvastatin Efficiently Reduces Levels of Alzheimer’s Amyloid Beta in Yeast. International Journal of Molecular Sciences. 2019; 20(14):3531. https://doi.org/10.3390/ijms20143531
Chicago/Turabian StyleDhakal, Sudip, Mishal Subhan, Joshua M. Fraser, Kenneth Gardiner, and Ian Macreadie. 2019. "Simvastatin Efficiently Reduces Levels of Alzheimer’s Amyloid Beta in Yeast" International Journal of Molecular Sciences 20, no. 14: 3531. https://doi.org/10.3390/ijms20143531
APA StyleDhakal, S., Subhan, M., Fraser, J. M., Gardiner, K., & Macreadie, I. (2019). Simvastatin Efficiently Reduces Levels of Alzheimer’s Amyloid Beta in Yeast. International Journal of Molecular Sciences, 20(14), 3531. https://doi.org/10.3390/ijms20143531